

Effects of Macleaya cordata Extract on Growth Performance, Serum Biochemical Parameters, and Intestinal Health of Juvenile American Eel (Anguilla rostrata)

Abstract
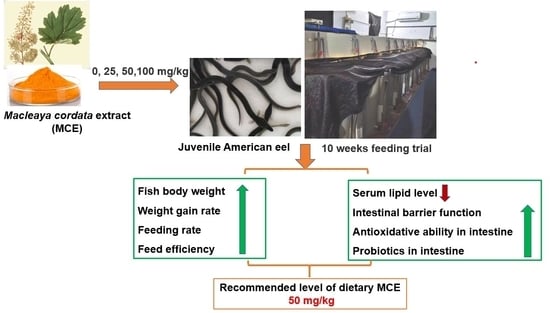
:
1. Introduction
2. Materials and Methods
2.1. Feeding Trial
2.2. Sample Collection
2.3. Growth Performance Calculation
2.4. Measurement of Serum Biochemical Parameters
2.5. Measurement of Intestinal Antioxidant Parameters
2.6. Observation and Measurement of Intestinal Morphology
2.7. Intestinal Microbiota Profiling
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Serum Biochemical Parameters
3.3. Intestinal Antioxidant Parameters
3.4. Intestinal Morphology
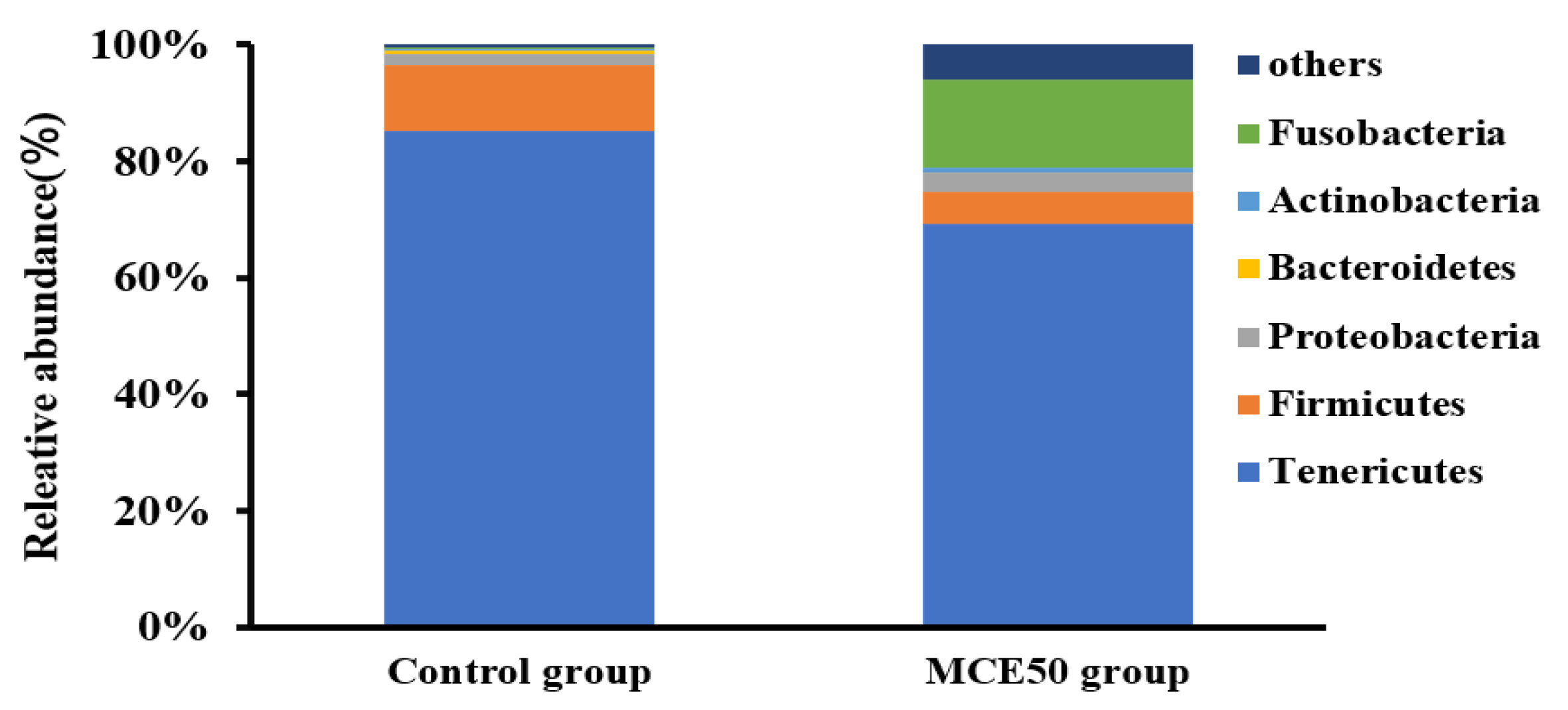
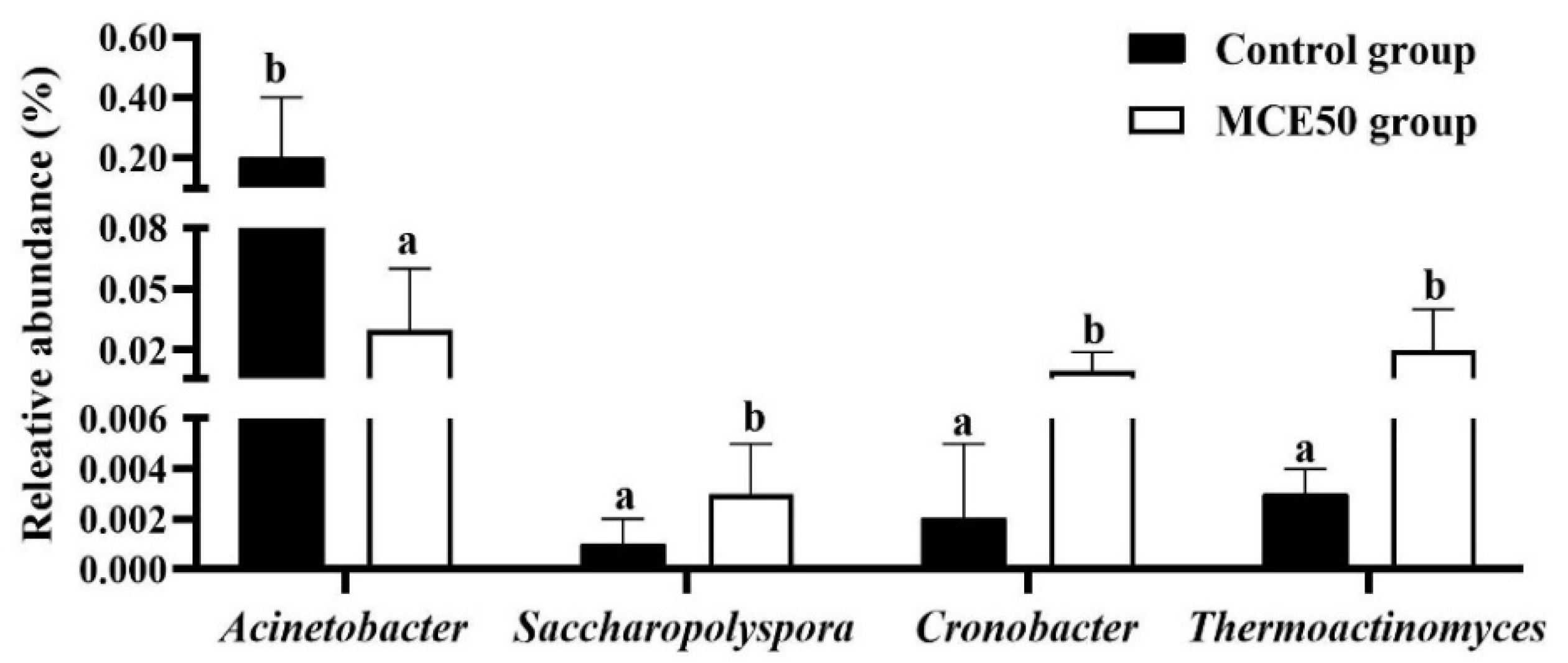
3.5. Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Tadese, D.A.; Song, C.; Sun, C.; Liu, B.; Liu, B.; Zhou, Q.; Xu, P.; Ge, X.; Liu, M.; Xu, X.; et al. The role of currently used medicinal plants in aquaculture and their action mechanisms: A review. Rev. Aquac. 2022, 14, 816–847. [Google Scholar] [CrossRef]
- Hou, B.; Zeng, J. Biological activities of sanguinarine and application of Macleaya cordata extract in animal production. Chin. J. Anim. Nutr. 2018, 30, 413–420. [Google Scholar] [CrossRef]
- Lin, L.; Liu, Y.C.; Huang, J.L.; Liu, X.B.; Qing, Z.X.; Zeng, J.G.; Liu, Z.Y. Medicinal plants of the genus Macleaya (Macleaya cordata, Macleaya microcarpa): A review of their phytochemistry, pharmacology, and toxicology. Phytother. Res. 2017, 32, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Zhong, L.; Chen, T.; Shi, Y.; Xu, S.D. Dietary sanguinarine supplementation on the growth performance, immunity and intestinal health of grass carp (Ctenopharyngodon idellus) fed cottonseed and rapeseed meal diets. Aquaculture 2020, 528, 735521. [Google Scholar] [CrossRef]
- Shi, Y.; Zhong, L.; Chen, K.J.; Fan, Y.D.; Xie, K.; Zhang, J.Z.; Dai, J.; Hu, Y. Sanguinarine attenuates hydrogen peroxide-induced toxicity in liver of Monopterus albus: Role of oxidative stress, inflammation and apoptosis. Fish Shellfish Immun. 2022, 125, 190–199. [Google Scholar] [CrossRef]
- Qin, F.; Patel, R.; Yan, C.; Liu, W. NADPH oxidase is involved in angiotensin II induced apoptosis in H9C2 cardiac muscle cells: Effects of apocynin. Free Radical Bio. Med. 2006, 40, 236–246. [Google Scholar] [CrossRef]
- Guan, G.; Ding, S.; Yin, Y.; Duraipandiyan, V.; AlDhabi, N.A.; Liu, G. Macleaya cordata extract alleviated oxidative stress and altered innate immune response in mice challenged with enterotoxigenic Escherichia coli. Sci. China Life Sci. 2019, 62, 1019–1027. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and anti-virulence activities. Int. J. Antimicrob. Ag. 2014, 44, 377–386. [Google Scholar] [CrossRef]
- Zhou, S.; Li, W.X.; Wang, Y.Q.; Zou, H.; Wu, S.G.; Wang, G.T. Anthelmintic efficacies of three common disinfectants and extracts of four traditional Chinese medicinal plants against Gyrodactylus kobayashii (Monogenea) in goldfish (Carassius auratus). Aquaculture 2017, 466, 72–77. [Google Scholar] [CrossRef]
- Liu, G.; Guan, G.P.; Fang, J.; Martinez, Y.; Chen, S.; Bin, P.; Duraipandiyan, V.; Gong, T.; Tossou, M.C.; Al-Dhabi, N.A.; et al. Macleaya cordata extract decreased diarrhea score and enhanced intestinal barrier function in growing piglets. Biomed Res. Int. 2016, 2016, 1069585. [Google Scholar] [CrossRef] [PubMed]
- Imanpoor, M.R.; Roohi, Z. Effects of Sangrovit®-supplemented diet on growth performance, blood biochemical parameters, survival and stress resistance to salinity in the Caspian roach (Rutilus rutilus) fry. Aquac. Res. 2015, 47, 2874–2880. [Google Scholar] [CrossRef]
- Rawling, M.D.; Merrifield, D.L.; Davies, S.J. Preliminary assessment of dietary supplementation of Sangrovit® on red tilapia (Oreochromis niloticus) growth performance and health. Aquaculture 2009, 294, 118–122. [Google Scholar] [CrossRef]
- Abdelnaby, E.A.; Mohamed, M.F.; Gammazh, A.K. Pharmacological studies of feed additives (Sanguinarine and Saccharomyces cerevisiae) on growth performance, hematological and intestinal bacterial count with challenge test by Aeromonas hydrophila in Cyprinus carpio. Global. Anim. Sci. J. 2013, 11, 1154–1172. [Google Scholar] [CrossRef]
- Zhang, M.L.; Wang, Y.; Zhai, S.W. Effects of dietary compound acidifiers supplementation on growth performance and intestinal health of juvenile American eels (Anguilla rostrata) cultured in cement tanks. Isr. J. Aquacult-Bamid. 2021, 73, 1520998. [Google Scholar] [CrossRef]
- Yuan, Y.; Yuan, Y.M.; Dai, Y.Y.; Gong, Y.C.; Yuan, Y.Q. Development status and trends in the eel farming industry in Asia. N. Am. J. Aquacult. 2021, 84, 3–17. [Google Scholar] [CrossRef]
- Shi, Y.; Ma, D.Y.; Zhai, S.W. Revealing the difference of intestinal microbiota composition of cultured European eels (Anguilla anguilla) with different growth rates. Isr. J. Aquacult-Bamid. 2020, 72, 959575. [Google Scholar] [CrossRef]
- Guo, D.; Xie, M.; Xiao, H.; Xu, L.; Zhang, S.; Chen, X.; Wu, Z. Bacillus subtilis supplementation in a high-fat diet modulates the gut microbiota and ameliorates hepatic lipid accumulation in grass carp (Ctenopharyngodon idella). Fishes 2022, 7, 94. [Google Scholar] [CrossRef]
- Tian, Y.; Cai, J.; Gui, W.; Nichols, R.G.; Koo, I.; Zhang, J.; Patterson, A.D. Berberine directly impacts the gut microbiota to promote intestinal farnesoid X receptor activation. Drug Metab. Dispos. 2019, 47, 86–93. [Google Scholar] [CrossRef]
- Farris, N.W.; Kim, D.; Hamidoghli, A.; Won, S.; Lee, S.; Bae, J. Dietary α-Tocopheryl acetate and arachidonic acid synergistically improves superoxide dismutase activity in female Japanese eel broodstock, Anguilla japonica. Aquaculture 2020, 522, 735100. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, Y.; Liu, Y.Y.; Che, C.B. Effects of sanguinarine on immune and intestinal inflammation related to gene expression in rice field eels (Monopterus albus) induced by LPS. J. Fish. Sci. China 2020, 27, 125–137. [Google Scholar] [CrossRef]
- Fukudome, I.; Kobayashi, M.; Dabanaka, K.; Maeda, H.; Okamoto, K.; Okabayashi, T.; Hanazaki, K. Diamine oxidase as a marker of intestinal mucosal injury and the effect of soluble dietary fiber on gastrointestinal tract toxicity after intravenous 5-fluorouracil treatment in rats. Med. Mol. Morphol. 2013, 47, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Mourente, G.; Tocher, D.R.; Diaz, E.; Grau, A.; Pastor, E. Relationships between antioxidants, antioxidant enzyme activities and lipid peroxidation products during early development in dentex dentex eggs and larvae. Aquaculture 1999, 179, 309–324. [Google Scholar] [CrossRef]
- Shi, Q.; Xiong, X.; Wen, Z.; Qin, C.; Li, R.; Zhang, Z.; Gong, Q.; Wu, X. Cu/Zn superoxide dismutase and catalase of Yangtze sturgeon, Acipenser dabryanus: Molecular cloning, tissue distribution and response to fasting and refeeding. Fishes 2022, 7, 35. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.W.; Zhu, J.Y.; Liu, L.L.; Liu, Y.C.; Zhu, H. Dietary sanguinarine affected immune response, digestive enzyme activity and intestinal microbiota of koi carp (Cryprinus carpiod). Aquaculture 2019, 502, 72–79. [Google Scholar] [CrossRef]
- Bussabong, P.; Rairat, T.; Chuchird, N.; Keetanon, A.; Phansawat, P.; Cherdkeattipol, K.; Pichitkul, P.; Kraitavin, W. Effects of isoquinoline alkaloids from Macleaya cordata on growth performance, survival, immune response, and resistance to Vibrio parahaemolyticus infection of Pacific white shrimp (Litopenaeus vannamei). PLoS ONE 2021, 16, e0251343. [Google Scholar] [CrossRef]
- Su, X.; Ji, D.; Yao, J.; Zou, Y.; Yan, M. Comparative analysis of intestinal characteristics of Largemouth Bass (Micropterus salmoides) and intestinal flora with different growth rates. Fishes 2022, 7, 65. [Google Scholar] [CrossRef]
- Saravanan, K.; Sivaramakrishnan, T.; Praveenraj, J.; Kiruba, S.R.; Haridas Harsha, K.S.; Varghese, B. Effects of single and multi-strain probiotics on the growth, hemato-immunological, enzymatic activity, gut morphology and disease resistance in Rohu, Labeo rohita. Aquaculture 2021, 540, 736749. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y.; Xue, M.; Xiao, Z.; Fan, Y.; Zeng, L.; Zhou, Y. Effects of dietary Enterococcus faecalis YFI-G720 on the growth, immunity, serum biochemical, intestinal morphology, intestinal microbiota, and disease resistance of Crucian Carp (Carassius auratus). Fishes 2022, 7, 18. [Google Scholar] [CrossRef]
- Ingerslev, H.C.; Jørgensen, L.V.G.; Strube, M.L.; Larsen, N.; Dalsgaard, I.; Boye, M. The development of the gut microbiota in rainbow trout (Oncorhynchus mykiss) is affected by first feeding and diet type. Aquaculture 2014, 424, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Feher, M.; Fauszt, P.; Tolnai, E.; Fidler, G.; Pesti, A.G.; Stagel, A.; Szucs, I.; Biro, S.; Remenyik, J.; Paholcsek, M.; et al. Effects of phytonutrient-supplemented diets on the intestinal microbiota of Cyprinus carpio. PLoS ONE 2021, 16, e0248537. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.J.; Xiao, S.Y.; Chen, S.Y.; Xu, Q.; Yang, Z.; Liu, J.; Wang, H.Q.; Lan, S.L. Effects of fermented mulberry leaves on growth, serum antioxidant capacity, digestive enzyme activities and microbial compositions of the intestine in Crucian (Carassius carassius). Aquac. Res. 2021, 52, 6356–6366. [Google Scholar] [CrossRef]
- Burtseva, O.; Kublanovskaya, A.; Fedorenko, T.; Lobakova, E.; Chekanov, K. Gut microbiome of the white sea fish revealed by 16s rRNA metabarcoding. Aquaculture 2020, 533, 736175. [Google Scholar] [CrossRef]
- Lin, M.; Zeng, C.X.; Jia, X.Q.; Zhai, S.W.; Li, Z.Q.; Ma, Y. The composition and structure of the intestinal microflora of Anguilla marmorata at different growth rates: A deep sequencing study. J. Appl. Microbiol. 2019, 126, 1340–1352. [Google Scholar] [CrossRef]
- Chen, Z.N.; Wang, X.Q.; Luo, L.T.; Wang, P.; Tu, K.F.; Yang, T.; Hu, Y.Z.; Xiong, G. High-throughput sequencing analysis of the effects of sanguinarine on Trionyx sinensis intestinal microbiota. Prog. Fish. Sci. 2021, 42, 177–185. [Google Scholar] [CrossRef]
- Zhai, S.; Wang, Y.; He, Y.; Chen, X.H. Oligomeric proanthocyanidins counteracts the negative effects of high level of dietary histamine on American Eel (Anguilla rostrata). Front. Mar. Sci. 2020, 7, 549145. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Ear, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Lobionda, S.; Sittipo, P.; Kwon, H.Y.; Lee, Y.K. The role of gut microbiota in intestinal inflammation with respect to diet and extrinsic stressors. Microorganisms 2019, 7, 271. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, Z.; Liu, X.; Xu, Y.; Bussabong, B.; Wang, B. Structural characteristics and succession of intestinal microbiota for Paralichthys olivaceus during the early life stage. Aquac. Res. 2019, 50, 529–540. [Google Scholar] [CrossRef]
- Azman, A.S.; Mawang, C.I.; Khairat, J.E.; Abubakar, S. Actinobacteria-a promising natural source of anti-biofilm agents. Int. Microbiol. 2019, 22, 403–409. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Dharmaraj, D.; Rajendran, K.; Karuppiah, K.; Ethiraj, K. Pharmacological activities of coral reef associated actinomycetes, Saccharopolyspora sp. IMA1. Biocatal. Agr. Biotechnol. 2020, 28, 101748. [Google Scholar] [CrossRef]
- Gupta, T.B.; Risson, A.N.; Brightwell, G.; Maclean, P.; Jauregui, R. Draft genome sequence of Thermoactinomyces vulgaris strain AGRTWHS02, isolated from pasture soil of a sheep dairy farm in New Zealand. Microbiol. Resour. Announc. 2022, 11, e0007622. [Google Scholar] [CrossRef] [PubMed]
- Fehr, A.; Eshwar, A.K.; Neuhauss, S.C.; Ruetten, M.; Vaughan, L. Evaluation of zebrafish as a model to study the pathogenesis of the opportunistic pathogen Cronobacter turicensis. Emerg. Microbes. Infec. 2015, 4, e29. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Kumar, S.H.; Nayak, B.B.; Lekshmi, M. Isolation and identification of Cronobacter spp. from fish and shellfish sold in retail markets. Curr. Microbiol. 2021, 78, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.H.; Liu Gao, P.W.; Xiao, L.C.; Shuang, S.Y.; Cong, H.; Du, Q.Y.; Zhang, X.W. Effects of Macleaya cordata extract on TLR20 and the proinflammatory cytokines in acute spleen injury of loach (Misgurnus anguillicaudatus) against Aeromonas hydrophila infection. Aquaculture 2021, 544, 737105. [Google Scholar] [CrossRef]
- Miao, F.; Yang, X.J.; Zhou, L.; Hu, H.J.; Zheng, F.; Ding, X.D.; Sun, W. Structural modification of sanguinarine and chelerythrine and their antibacterial activity. Natural Product Res. 2011, 25, 863–875. [Google Scholar] [CrossRef]
- Beuria, T.K.; Santra, M.K.; Panda, D. Sanguinarine blocks cytokinesis in bacteria by inhibiting FtsZ assembly and bundling. Biochemistry 2005, 44, 16584–16593. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Control Group | MCE25 Group | Mce50 Group | Mce100 Group |
|---|---|---|---|---|
| FBW (g/fish) | 19.86 ± 0.49 a | 19.77 ± 0.42 a | 22..93 ± 0.57 b | 23.30 ± 0.49 b |
| WGR (%) | 81.82 ± 4.59 a | 79.76 ± 2.54 a | 109.88 ± 4.76 b | 113.93 ± 3.97 b |
| SGR (%/d) | 0.81 ± 0.03 a | 0.79 ± 0.02 a | 1.00 ± 0.03 b | 1.03 ± 0.02 b |
| FR (%) | 1.02 ± 0.03 a | 1.01 ± 0.01 a | 1.17 ± 0.02 b | 1.05 ± 0.04 a |
| FE (%) | 76.69 ± 4.10 a | 76.00 ± 2.39 a | 82.80 ± 3.41 ab | 93.28 ± 2.89 b |
| SR (%) | 100 | 97.5 ± 1.60 | 99.17 ± 0.83 | 100 |
| Items | Control Group | MCE25 Group | Mce50 Group | Mce100 Group |
|---|---|---|---|---|
| TC (mmol/L) | 32.85 ± 0.77 b | 31.74 ± 0.44 b | 31.51 ± 1.19 b | 28.14 ± 0.59 a |
| TG (mmol/L) | 6.00 ± 0.11 b | 5.89 ± 0.07 b | 5.61 ± 0.08 b | 4.18 ± 0.14 a |
| HDL-C mmol/L) | 4.61 ± 0.18 a | 4.71 ± 0.12 a | 5.59 ± 0.18 b | 5.61 ± 0.17 b |
| LDL-C (mmol/L) | 21.77 ± 0.89 a | 21.79 ± 1.23 a | 21.47 ± 0.35 a | 21.81 ± 0.69 a |
| GOT (IU/L) | 13.58 ± 0.57 b | 12.98 ± 0.35 ab | 11.99 ± 0.47 a | 12.67 ± 0.34 ab |
| GPT (IU/L) | 8.77 ± 0.37 b | 8.42 ± 0.26 ab | 7.41 ± 0.38 a | 8.24 ± 0.21 ab |
| DAO (U/L) | 29.22 ± 0.71 b | 23.75 ± 1.01 a | 21.88 ± 1.03 a | 22.00 ± 1.21 a |
| D-lac (mmol/L) | 1.41 ± 0.02 c | 1.02 ± 0.01 b | 0.98 ± 0.03 b | 0.83 ± 0.05 a |
| Items | Control Group | MCE25 Group | MCE50 Group | MCE100 Group |
|---|---|---|---|---|
| T-AOC (U/mg prot) | 0.13 ± 0.01 a | 0.18 ± 0.01 b | 0.25 ± 0.01 c | 0.23 ± 0.02 c |
| SOD (U/mg prot) | 57.99 ± 1.76 a | 58.73 ± 0.12 a | 59.72 ± 0.49 a | 67.71 ± 0.94 b |
| CAT (U/mg prot) | 4.31 ± 0.15 a | 4.34 ± 0.01 a | 5.62 ± 0.37 b | 6.18 ± 0.07 b |
| GSH-PX (U/mg prot) | 15.75 ± 0.88 a | 19.06 ± 0.09 b | 21.94 ± 0.33 c | 20.68 ± 0.58 bc |
| MDA (nmol/mg prot) | 30.66 ± 0.12 c | 22.16 ± 0.64 b | 19.19 ± 0.98 a | 21.12 ± 0.66 ab |
| Items | Control Group | MCE25 Group | MCE50 Group | MCE100 Group |
|---|---|---|---|---|
| MT (μm) | 131.11 ± 1.20 a | 137.59 ± 2.74 a | 146.10 ± 1.63 b | 165.48 ± 2.64 c |
| FH (μm) | 530.69 ± 8.89 a | 559.52 ± 5.05 a | 619.47 ± 6.50 b | 599.97 ± 8.36 b |
| Items | Control Group | MCE50 Group |
|---|---|---|
| OTUs | 201 | 600 |
| Chao 1 | 148.96 ± 21.20 | 167.22 ± 14.58 |
| Shannon | 0.73 ± 0.34 | 0.85 ± 0.16 |
| Coverage rate (%) | 99 | 98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Huang, L.; Zhai, S. Effects of Macleaya cordata Extract on Growth Performance, Serum Biochemical Parameters, and Intestinal Health of Juvenile American Eel (Anguilla rostrata). Fishes 2022, 7, 229. https://doi.org/10.3390/fishes7050229
Chen R, Huang L, Zhai S. Effects of Macleaya cordata Extract on Growth Performance, Serum Biochemical Parameters, and Intestinal Health of Juvenile American Eel (Anguilla rostrata). Fishes. 2022; 7(5):229. https://doi.org/10.3390/fishes7050229
Chicago/Turabian StyleChen, Runan, Linxin Huang, and Shaowei Zhai. 2022. "Effects of Macleaya cordata Extract on Growth Performance, Serum Biochemical Parameters, and Intestinal Health of Juvenile American Eel (Anguilla rostrata)" Fishes 7, no. 5: 229. https://doi.org/10.3390/fishes7050229
APA StyleChen, R., Huang, L., & Zhai, S. (2022). Effects of Macleaya cordata Extract on Growth Performance, Serum Biochemical Parameters, and Intestinal Health of Juvenile American Eel (Anguilla rostrata). Fishes, 7(5), 229. https://doi.org/10.3390/fishes7050229