
Toxic Effects of Arsenic in Commercially Important Fish Rohu Carp, Labeo rohita of Bangladesh

 and
and
Abstract
1. Introduction
2. Materials and Methods
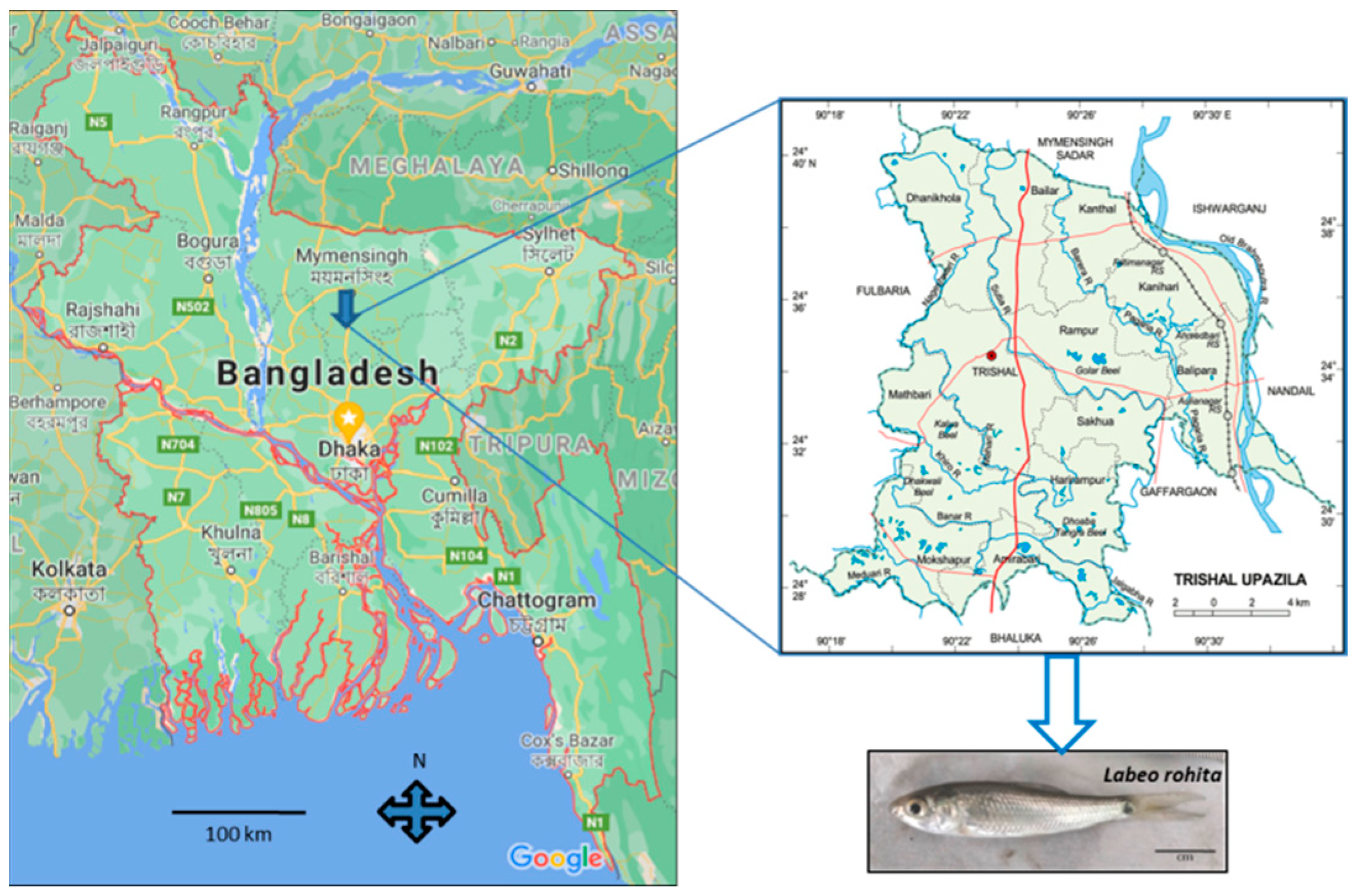
2.1. Transportation
2.2. Acclimatization
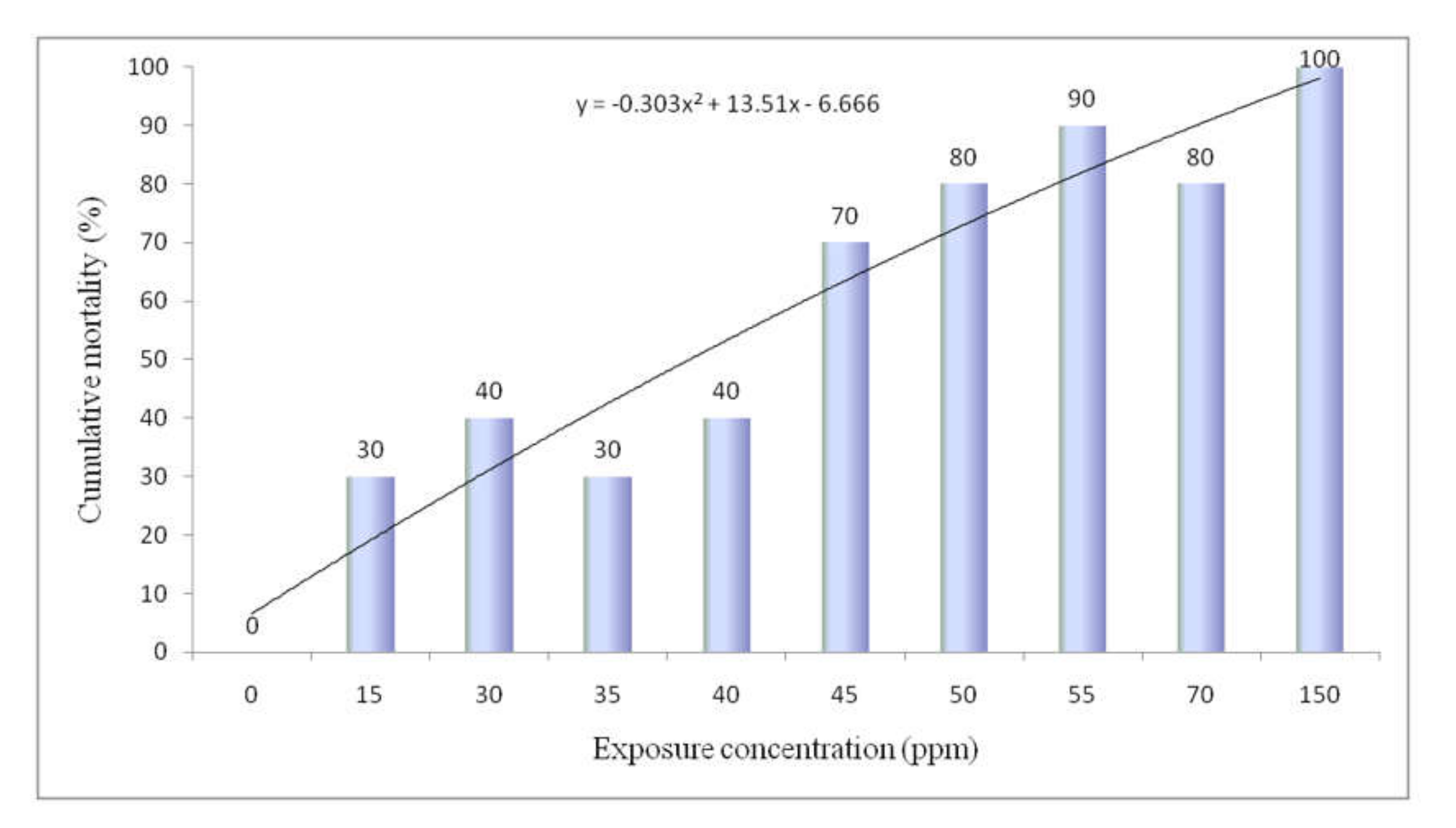
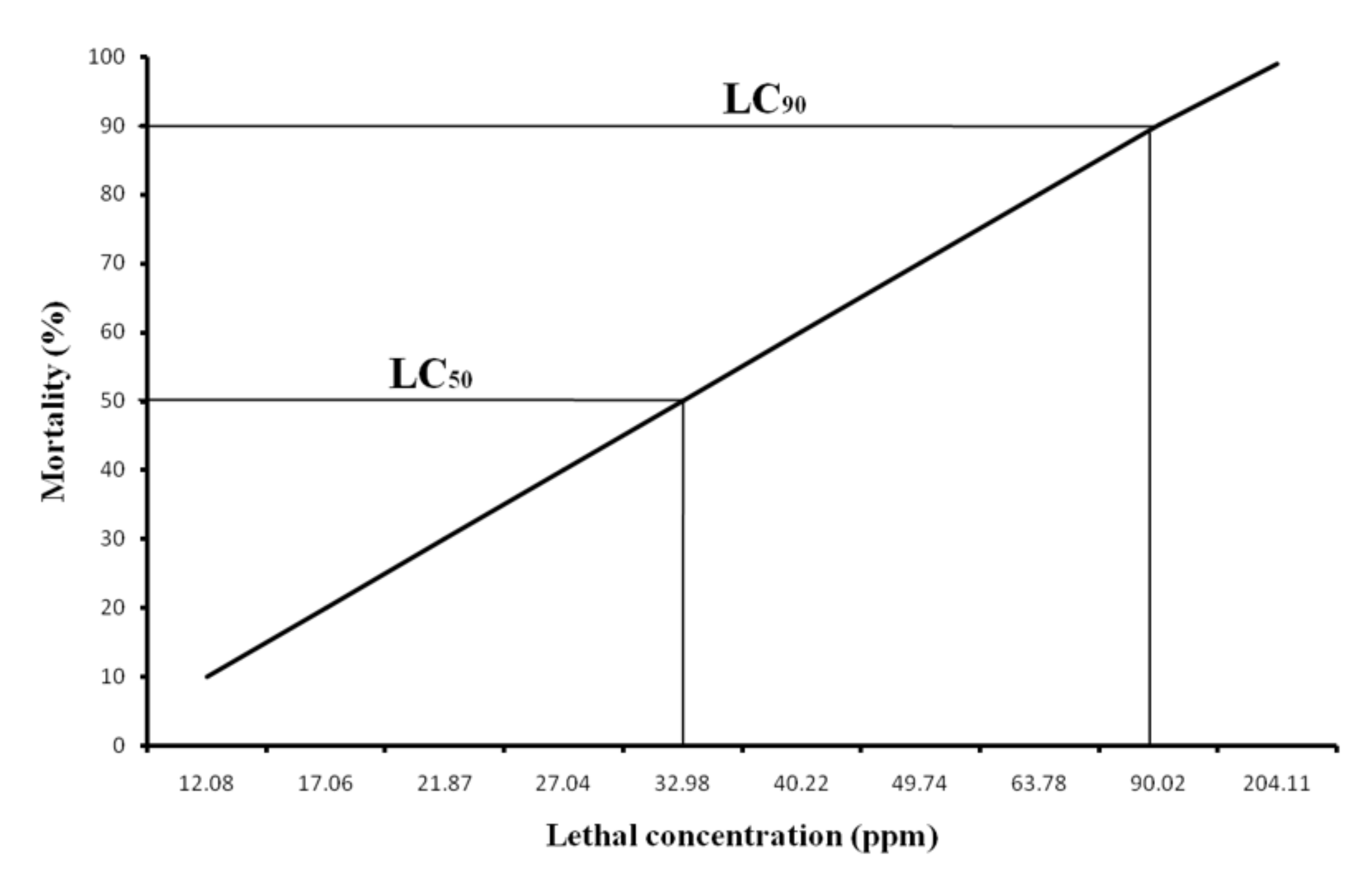
2.3. Stock Solution Preparation and LC50 Determination for Sodium Arsenite
2.4. Experimental Design for Exposure to Sodium Arsenite
2.5. Growth and Mortality Study
- (a)
- Weight gain = Mean final fish weight − Mean initial fish weight
- (b)
- Length gain (cm) = Mean final length (cm) − Mean initial length (cm)
- (c)
- (d)
- Mortality rate (%) = × 100
2.6. Morphological Observation
2.7. Clinical Signs/Behavioral Changes
2.8. Histopathology
2.9. Determination of RNA:DNA Ratio
2.10. Gene Expression Study
2.11. Data Analysis
3. Results
3.1. Physicochemical Parameters of Culture Water
3.2. Cumulative Mortality and Determination of LC50
3.3. Morphological Anomalies and Deformities
3.4. Clinical Signs/Behavioral Changes
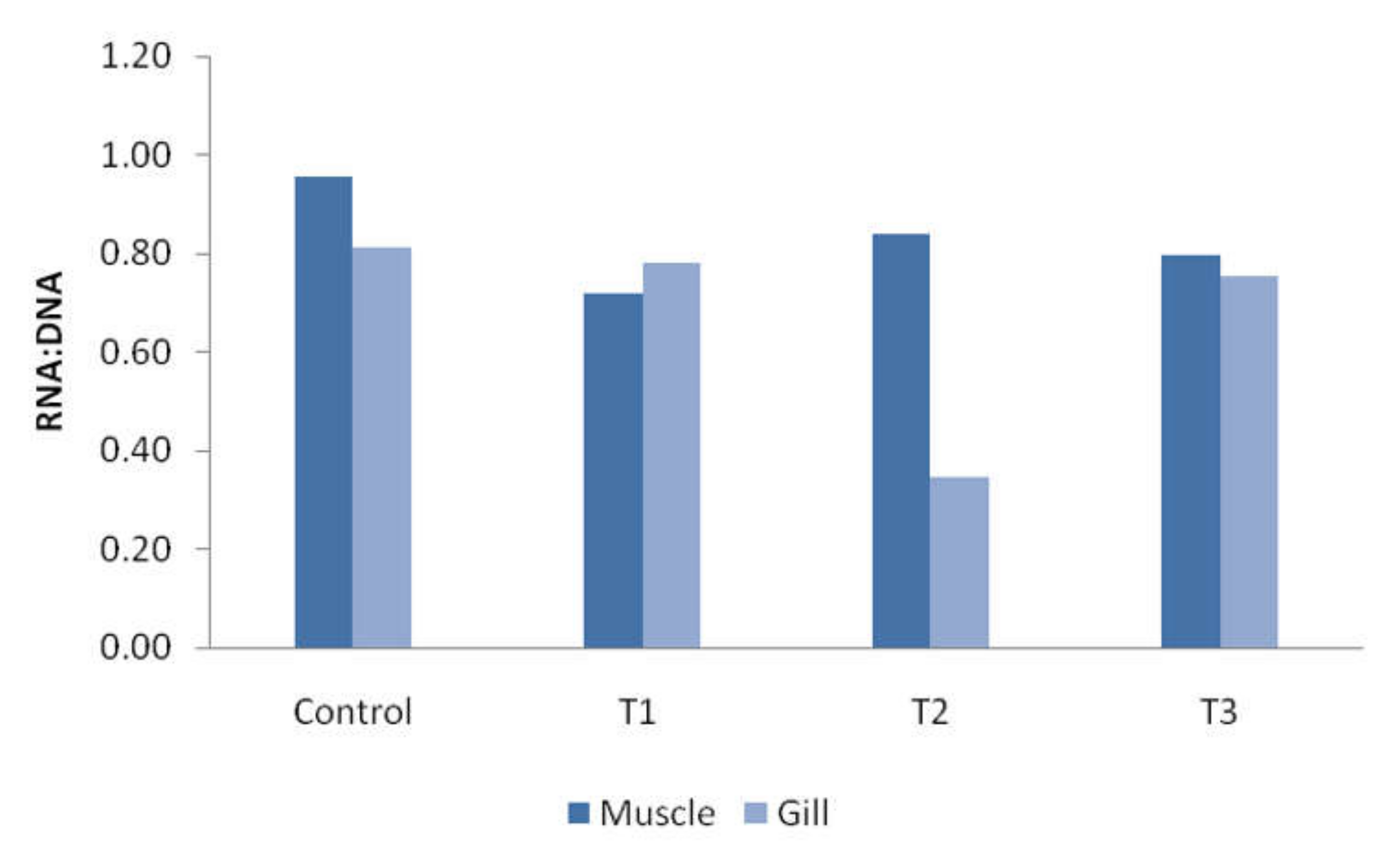
3.5. RNA and DNA Ratio
3.6. Growth Parameter Study
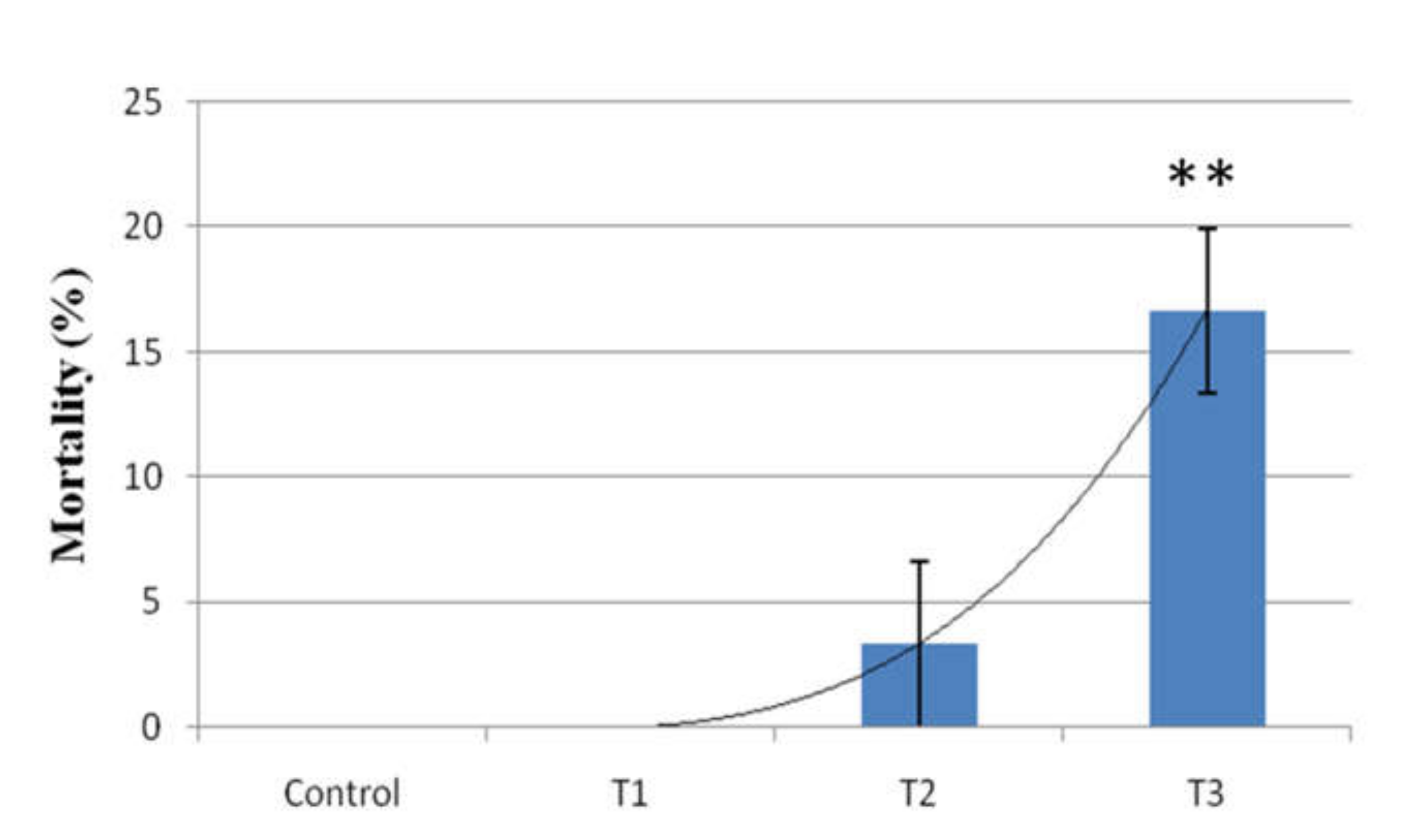
3.7. Mortality Rate Study
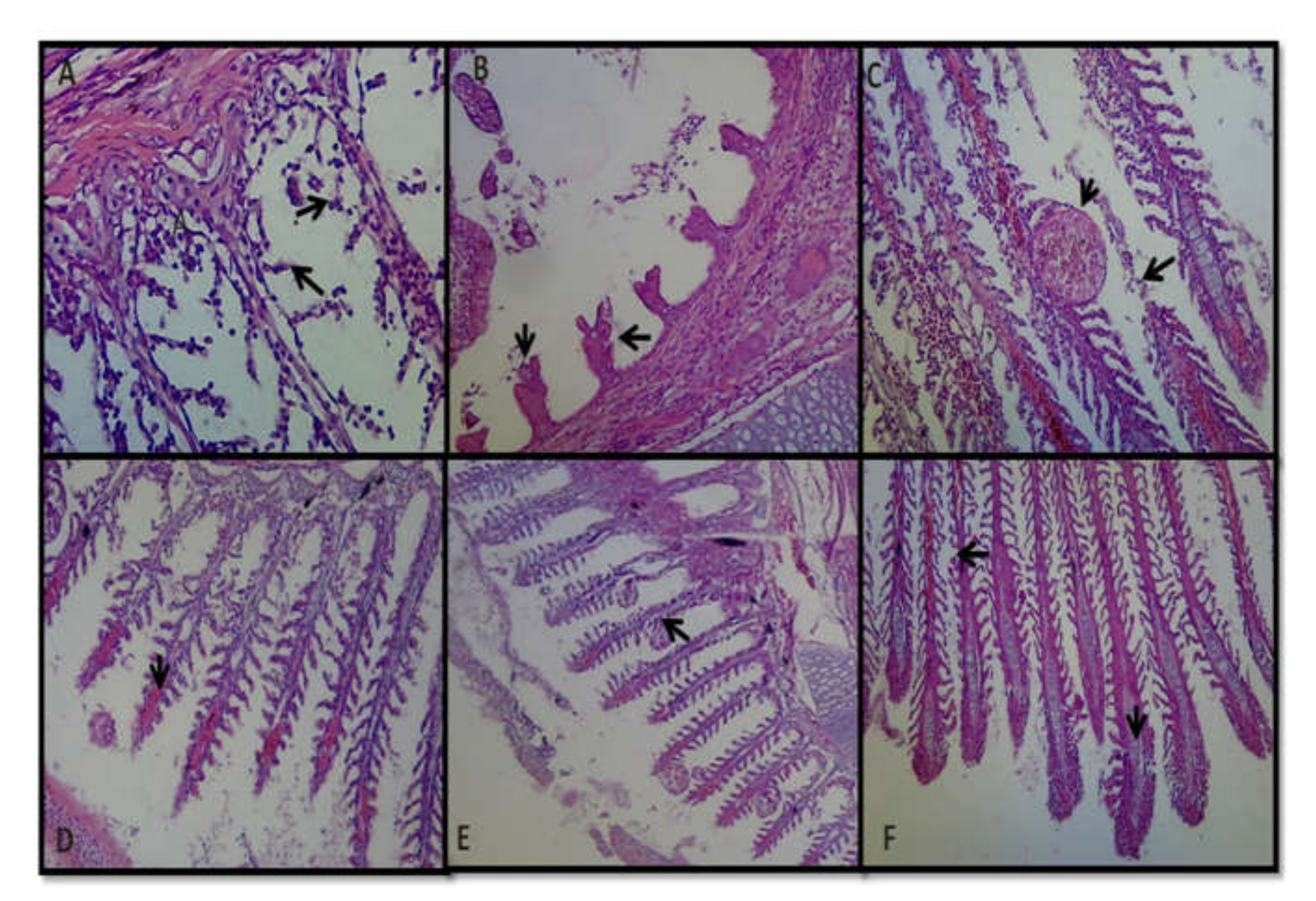
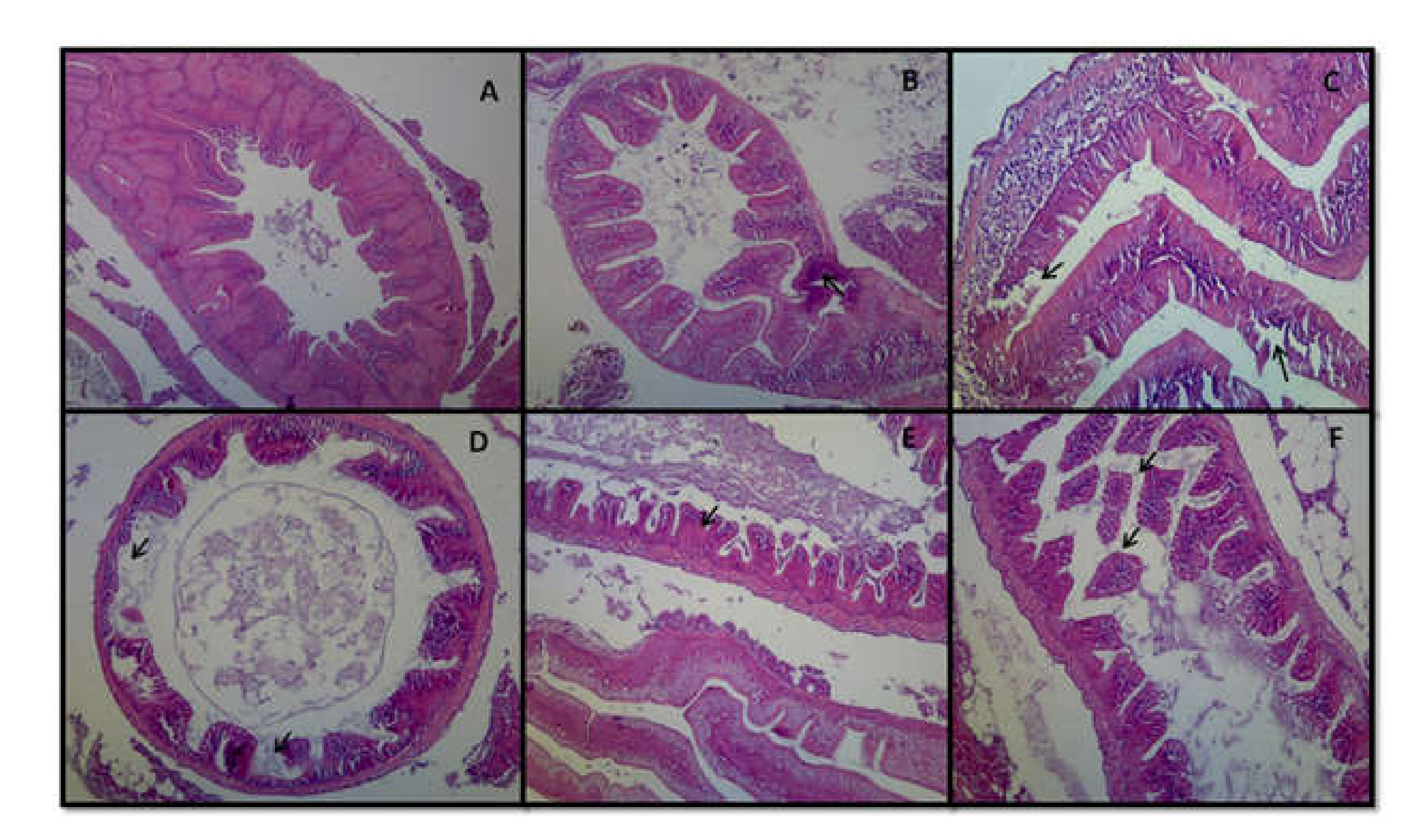
3.8. Histopathology
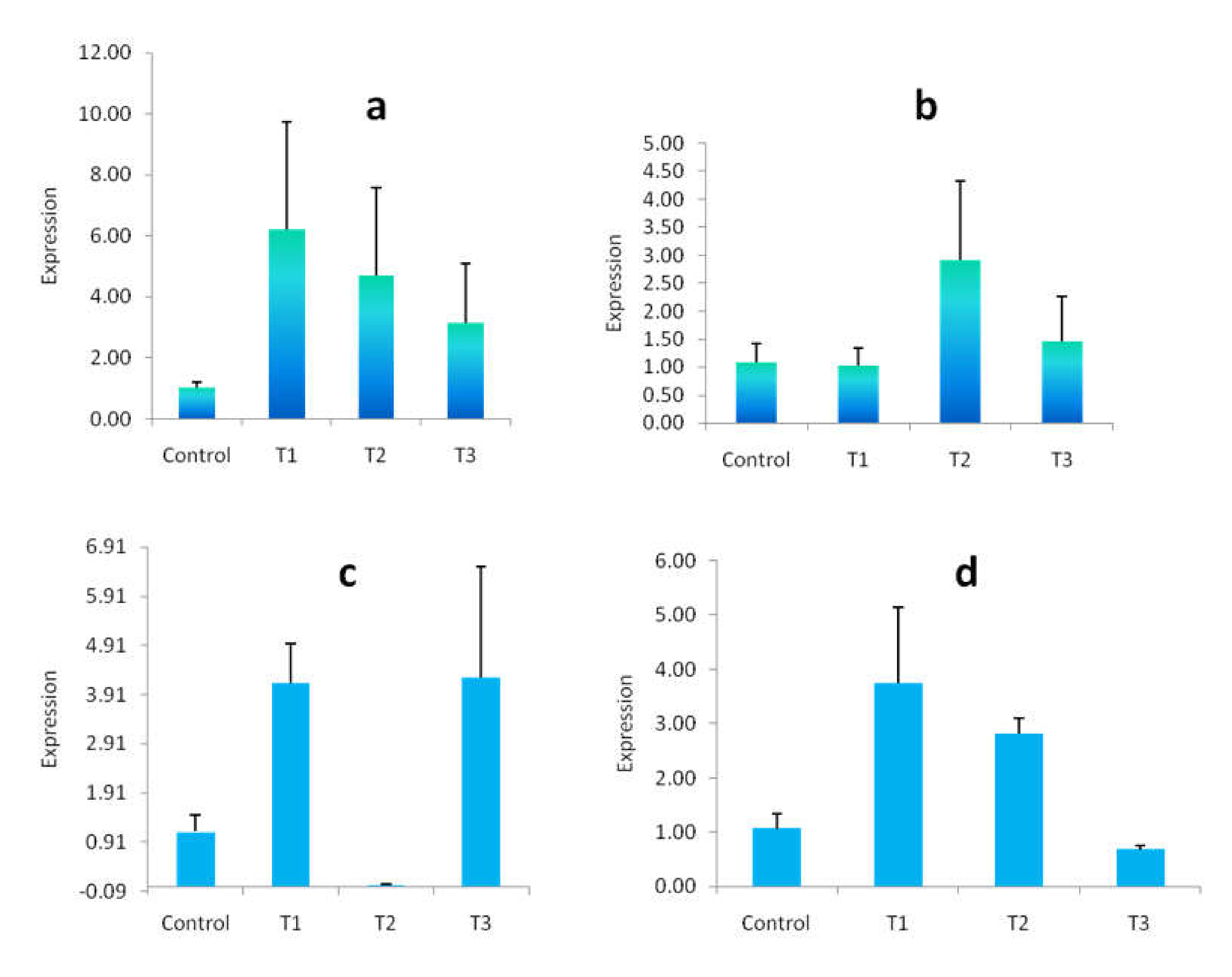
3.9. Gene Expression Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, Y.-J.; Jia, Y.-F.; Chen, N.; Bian, W.-P.; Li, Q.-K.; Ma, Y.-B.; Chen, Y.-L.; Pei, D.-S. Zebrafish as a model system to study toxicology. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Jezierska, B.; Ługowska, K.; Witeska, M. The effects of heavy metals on embryonic development of fish (a review). Fish Physiol. Biochem. 2008, 35, 625–640. [Google Scholar] [CrossRef]
- Jezierska, B.; Witeska, M. Metal Toxicity to Fish; Wydawnictwo Akademii Podlaskiej: Siedlce, Poland, 2001; 318p. [Google Scholar]
- Ratnaike, R.N. Acute and chronic arsenic toxicity. Postgrad. Med. J. 2003, 79, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S.; Stolz, J.F. The Ecology of Arsenic. Science 2003, 300, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Kumari, B.; Kumar, V.; Sinha, A.K.; Ahsan, J.; Ghosh, A.; Wang, H.; De Boeck, G. Toxicology of arsenic in fish and aquatic systems. Environ. Chem. Lett. 2016, 15, 43–64. [Google Scholar] [CrossRef]
- Babich, R.; Van Beneden, R.J. Effect of arsenic exposure on early eye development in zebrafish (Danio rerio). J. Appl. Toxicol. 2019, 39, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Naujokas, M.F.; Anderson, B.; Ahsan, H.; Aposhian, H.V.; Graziano, J.H.; Thompson, C.; Suk, W.A. The Broad Scope of Health Effects from Chronic Arsenic Exposure: Update on a Worldwide Public Health Problem. Environ. Health Perspect. 2013, 121, 295–302. [Google Scholar] [CrossRef]
- Bräuner, E.V.; Nordsborg, R.B.; Andersen, Z.J.; Tjønneland, A.; Loft, S.; Raaschou-Nielsen, O. Long-Term Exposure to Low-Level Arsenic in Drinking Water and Diabetes Incidence: A Prospective Study of the Diet, Cancer and Health Cohort. Environ. Health Perspect. 2014, 122, 1059–1065. [Google Scholar] [CrossRef]
- Gong, G.; O’Bryant, S.E. Low-level arsenic exposure, AS3MT gene polymorphism and cardiovascular diseases in rural Texas counties. Environ. Res. 2012, 113, 52–57. [Google Scholar] [CrossRef]
- Kundu, M.; Ghosh, P.; Mitra, S.; Das, J.; Sau, T.; Banerjee, S.; States, J.C.; Giri, A.K. Precancerous and non-cancer disease endpoints of chronic arsenic exposure: The level of chromosomal damage and XRCC3 T241M polymorphism. Mutat. Res. Mol. Mech. Mutagen. 2011, 706, 7–12. [Google Scholar] [CrossRef]
- Liu, J.; Waalkes, M.P. Liver is a Target of Arsenic Carcinogenesis. Toxicol. Sci. 2008, 105, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Javed, M.; Razzaq, S. Growth performance of metal stressed major carps viz. Catla catla, Labeo rohita and Cirrhina mrigala reared under semi-intensive culture system. Pak. Vet. J. 2007, 27, 8–12. [Google Scholar]
- Han, J.-M.; Park, H.-J.; Kim, J.-H.; Jeong, D.-S.; Kang, J.-C. Toxic effects of arsenic on growth, hematological parameters, and plasma components of starry flounder, Platichthys stellatus, at two water temperature conditions. Fish. Aquat. Sci. 2019, 22, 3. [Google Scholar] [CrossRef]
- Foley, C.J.; Bradley, D.L.; Höök, T.O. A review and assessment of the potential use of RNA:DNA ratios to assess the condition of entrained fish larvae. Ecol. Indic. 2016, 60, 346–357. [Google Scholar] [CrossRef]
- Ahmed, K.; Mamun, H.A.; Parvin, E.; Akter, M.S.; Khan, M.S. Arsenic induced toxicity and histopathological changes in gill and liver tissue of freshwater fish, tilapia (Oreochromis mossambicus). Exp. Toxicol. Pathol. 2013, 65, 903–909. [Google Scholar] [CrossRef]
- Banerjee, S.; Mitra, T.; Purohit, G.K.; Mohanty, S.; Mohanty, B.P. Immunomodulatory effect of arsenic on cytokine and HSP gene expression in Labeo rohita fingerlings. Fish Shellfish. Immunol. 2015, 44, 43–49. [Google Scholar] [CrossRef]
- Minatel, B.C.; Sage, A.P.; Anderson, C.; Hubaux, R.; Marshall, E.A.; Lam, W.L.; Martinez, V.D. Environmental arsenic exposure: From genetic susceptibility to pathogenesis. Environ. Int. 2017, 112, 183–197. [Google Scholar] [CrossRef]
- Beyers, D.W.; Rice, J.A.; Clements, W.H.; Henry, C.J. Estimating physiological cost of chemical exposure: Integrating energetics and stress to quantify toxic effects in fish. Can. J. Fish. Aquat. Sci. 1999, 56, 814–822. [Google Scholar] [CrossRef]
- Pedlar, R.M.; Ptashynski, M.D.; Evans, R.; Klaverkamp, J.F. Toxicological effects of dietary arsenic exposure in lake whitefish (Coregonus clupeaformis). Aquat. Toxicol. 2002, 57, 167–189. [Google Scholar] [CrossRef]
- Farag, A.M.; Stansbury, M.A.; Bergman, H.L.; Hogstrand, C.; MacConnell, E. The physiological impairment of free-ranging brown trout exposed to metals in the Clark Fork River, Montana. Can. J. Fish. Aquat. Sci. 1995, 52, 2038–2050. [Google Scholar] [CrossRef]
- Datta, S.; Ghosh, D.; Saha, D.R.; Bhattacharaya, S.; Mazumder, S. Chronic exposure to low concentration of arsenic is immunotoxic to fish: Role of head kidney macrophages as biomarkers of arsenic toxicity to Clarias batrachus. Aquat. Toxicol. 2009, 92, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.; Usmani, N. An Overview of the Adverse Effects of Heavy Metal Contamination on Fish Health. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2017, 89, 389–403. [Google Scholar] [CrossRef]
- Bose, M.J.; Ilavazhahan, M.; Tamilselvi, R.; Viswanathan, M. Effect of Heavy Metals on the Histopathology of Gills and Brain of Fresh Water Fish Catla catla. Biomed. Pharmacol. J. 2013, 6, 99–105. [Google Scholar] [CrossRef]
- Selvi, R.; Ilavazhahan, M. Histopathological Changes in Gill Tissue of the Fish Catla catla Exposed to Sublethal Concentration of Pesticide Methyl Parathion and a Heavy Metal Ferous Sulphate. Biomed. Pharmacol. J. 2012, 5, 305–312. [Google Scholar] [CrossRef]
- Chavan, V.R.; Muley, D.V. Effect of heavy metals on liver and gill of fish Cirrhinus mrigala. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 277–288. [Google Scholar]
- Padhy, G.; Mishra, C.; Barik, B.P. Histological alterations in gill tissues of Anabas testiduneus on exposure to heavy metal CdCl2. RJLBPCS 2018, 4, 328–334. [Google Scholar] [CrossRef]
- Gürcü, B.; Yildiz, S.; Koca, Y.B.G.; Koca, S. Investigation of Histopathological and Cytogenetic Effects of Heavy Metals Pollution on Cyprinus carpio (Linneaus, 1758) in the Gölmarmara Lake, Turkey. J. Anim. Vet. Adv. 2010, 9, 798–808. [Google Scholar] [CrossRef]
- Yildiz, S.; Gürcü, B.; Koca, Y.B.; Koca, S. Histopathological and Genotoxic Effects of Pollution on Anguilla anguilla in the Gediz River (Turkey). J. Anim. Vet. Adv. 2010, 9, 2890–2899. [Google Scholar] [CrossRef]
- Lam, K.L.; Ko, P.W.; Wong, J.K.-Y.; Chan, K.M. Metal toxicity and metallothionein gene expression studies in common carp and tilapia. Mar. Environ. Res. 1998, 46, 563–566. [Google Scholar] [CrossRef]
- Olsson, P.-E.; Kille, P. Functional comparison of the metal-regulated transcriptional control regions of metallothionein genes from cadmium-sensitive and tolerant fish species. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 1997, 1350, 325–334. [Google Scholar] [CrossRef]
- Kakkar, V.; Meister-Broekema, M.; Minoia, M.; Carra, S.; Kampinga, H.H. Barcoding heat shock proteins to human diseases: Looking beyond the heat shock response. Dis. Model. Mech. 2014, 7, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.K. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef]
- Purohit, G.K.; Mahanty, A.; Suar, M.; Sharma, A.P.; Mohanty, B.P.; Mohanty, S. Investigating hsp gene expression in liver of Channa striatus under heat stress for understanding the upper thermal acclimation. Biomed. Res. Int. 2014, 2014, 381719. [Google Scholar] [CrossRef]
- Rabbane, G.; Ali, Y.; Zahid, A.; Hossain, J. Diet Effects on Growth, Mortality, RNA: DNA Ratio and Gene Expression of Zebrafish Danio rerio. Genet. Aquat. Org. 2020, 4, 19–27. [Google Scholar] [CrossRef]
- Pandi Prabha, S.; Rajkumar, J.; Karthik, C. Hepatotoxic effect of lead and hepatoprotective effect of Hydrilla verticillata on hepatic transcriptional and physiological response in edible fish Labeo rohita. Drug Chem. Toxicol. 2020, 45, 1276–1283. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATSDR) 2007. Toxicological Profile for Arsenic. Atlanta, GA: U.S. Department of Health and Human Services, Public Health Service. Available online: https://www.atsdr.cdc.gov/substances/toxsubstance.asp?toxid=3 (accessed on 30 May 2021).
- Sfakianakis, D.; Renieri, E.; Kentouri, M.; Tsatsakis, A. Effect of heavy metals on fish larvae deformities: A review. Environ. Res. 2015, 137, 246–255. [Google Scholar] [CrossRef]
- Lavanya, S.; Ramesh, M.; Kavitha, C.; Malarvizhi, A. Hematological, biochemical and ionoregulatory responses of Indian major carp Catla catla during chronic sublethal exposure to inorganic arsenic. Chemosphere 2011, 82, 977–985. [Google Scholar] [CrossRef]
- Ghosh, D.; Bhattacharya, S.; Mazumder, S. Perturbations in the catfish immune responses by arsenic: Organ and cell specific effects. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Bhattacharya, S. Induction of oxidative stress by arsenic in Clarias batrachus: Involvement of peroxisomes. Ecotoxicol. Environ. Saf. 2007, 66, 178–187. [Google Scholar] [CrossRef]
- Roy, S.; Bhattacharya, S. Arsenic-induced histopathology and synthesis of stress proteins in liver and kidney of Channa punctatus. Ecotoxicol. Environ. Saf. 2006, 65, 218–229. [Google Scholar] [CrossRef]
- Akter, M.S.; Ahmed, M.K.; Akhand, M.A.A.; Islam, M.M. Acute toxicity of arsenic and mercury to fresh water climbing perch, Anabas testudineus (Bloch). World J. Zool. 2008, 3, 13–18. [Google Scholar]
- Agusa, T.; Takagi, K.; Kubota, R.; Anan, Y.; Iwata, H.; Tanabe, S. Specific accumulation of arsenic compounds in green turtles (Chelonia mydas) and hawksbill turtles (Eretmochelys imbricata) from Ishigaki Island, Japan. Environ. Pollut. 2008, 153, 127–136. [Google Scholar] [CrossRef]
- Cheng, S.H.; Wai, A.W.K.; So, C.H.; Wu, R.S.S. Cellular and molecular basis of cadmium-induced deformities in zebrafish embryos. Environ. Toxicol. Chem. 2000, 19, 3024–3031. [Google Scholar] [CrossRef]
- Hallare, A.; Schirling, M.; Luckenbach, T.; Köhler, H.-R.; Triebskorn, R. Combined effects of temperature and cadmium on developmental parameters and biomarker responses in zebrafish (Danio rerio) embryos. J. Therm. Biol. 2005, 30, 7–17. [Google Scholar] [CrossRef]
- Baldissarelli, L.A.; Capiotti, K.M.; Bogo, M.R.; Ghisleni, G.; Bonan, C.D. Arsenic alters behavioral parameters and brain ectonucleotidases activities in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Pepin, P.; Evans, G.T.; Shears, T.H. Patterns of RNA/DNA ratios in larval fish and their relationship to survival in the field. ICES J. Mar. Sci. 1999, 56, 697–706. [Google Scholar] [CrossRef]
- Gwak, W.-S.; Tanaka, Y.; Tominaga, O.; Tsusaki, T.; Tanaka, M. Field evaluation by RNA/DNA ratios on post-release nutritional status of released and wild Japanese flounder Paralichthys olivaceus juveniles. J. Exp. Mar. Biol. Ecol. 2003, 293, 107–124. [Google Scholar] [CrossRef]
- Smith, T.R.; Buckley, L.J. RNA–DNA ratio in scales from juvenile cod pro-vides a nonlethal measure of feeding condition. Trans. Am. Fish. Soc. 2003, 132, 9–17. [Google Scholar] [CrossRef]
- Kyle, M.; Watts, T.; Schade, J.; Elser, J.J. A microfluorometric method for quantifying RNA and DNA in terrestrial insects. J. Insect Sci. 2003, 3, 1. [Google Scholar] [CrossRef]
- Berdalet, E.; Dortch, Q. New double-staining technique for RNA and DNA measurement in marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 73, 295–305. [Google Scholar] [CrossRef]
- Gorokhova, E.; Kyle, M. Analysis of nucleic acids in Daphnia: Development of methods and ontogenetic variations in RNA-DNA content. J. Plankton Res. 2002, 24, 511–522. [Google Scholar] [CrossRef]
- Gorokhova, E. Relationships between nucleic acid levels and egg production rates in Acartia bifilosa: Implications for growth assessment of copepods in situ. Mar. Ecol. Prog. Ser. 2003, 262, 163–172. [Google Scholar] [CrossRef][Green Version]
- Suthers, I.; Cleary, J.; Battaglene, S.; Evans, R.; Suthers, I. Relative RNA Content as a Measure of Condition in Larval and Juvenile Fish. Mar. Freshw. Res. 1996, 47, 301–307. [Google Scholar] [CrossRef]
- Ferron, A.; Leggett, W. An Appraisal of Condition Measures for Marine Fish Larvae. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 1994; Volume 30, pp. 217–303. [Google Scholar] [CrossRef]
- Li, Z.-H.; Li, P.; Shi, Z.-C. Molecular responses in digestive tract of juvenile common carp after chronic exposure to sublethal tributyltin. Ecotoxicol. Environ. Saf. 2014, 109, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Varó, I.; Navarro, J.; Nunes, B.; Guilhermino, L. Effects of dichlorvos aquaculture treatments on selected biomarkers of gilthead sea bream (Sparus aurata L.) fingerlings. Aquaculture 2007, 266, 87–96. [Google Scholar] [CrossRef]
- Kim, S.-G.; Kang, J.-C. Effect of dietary copper exposure on accumulation, growth and hematological parameters of the juvenile rockfish, Sebastes schlegeli. Mar. Environ. Res. 2004, 58, 65–82. [Google Scholar] [CrossRef]
- De Boeck, G.; Borger, R.; Van der Linden, A.; Blust, R. Effects of sublethal copper exposure on muscle energy metabolism of common carp, measured by31P-nuclear magnetic resonance spectroscopy. Environ. Toxicol. Chem. 1997, 16, 676–684. [Google Scholar] [CrossRef]
- Miliou, H.; Zaboukas, N.; Moraitou-Apostolopoulou, M. Biochemical Composition, Growth, and Survival of the Guppy, Poecilia reticulata, During Chronic Sublethal Exposure to Cadmium. Arch. Environ. Contam. Toxicol. 1998, 35, 58–63. [Google Scholar] [CrossRef]
- Hussain, S.M.; Javed, M.; Asghar, S.; Hussain, M.; Abdullah, S.; Raza, S.A.; Javid, A. Studies on growth performance of metals mixture stressed Cirrhina mrigala in earthen ponds. Pak. J. Agric. Sci. 2010, 47, 263–270. [Google Scholar]
- Canli, M.; Atli, G. The relationships between heavy metal (Cd, Cr, Cu, Fe, Pb, Zn) levels and the size of six Mediterranean fish species. Environ. Pollut. 2003, 121, 129–136. [Google Scholar] [CrossRef]
- Bervoets, L.; Knapen, D.; De Jonge, M.; Van Campenhout, K.; Blust, R. Differential Hepatic Metal and Metallothionein Levels in Three Feral Fish Species along a Metal Pollution Gradient. PLoS ONE 2013, 8, e60805. [Google Scholar] [CrossRef] [PubMed]
- Woodward, D.F.; Brumbaugh, W.G.; DeLonay, A.J.; Little, E.E.; Smith, C.E. Effects on Rainbow Trout Fry of a Metals-Contaminated Diet of Benthic Invertebrates from the Clark Fork River, Montana. Trans. Am. Fish. Soc. 1994, 123, 51–62. [Google Scholar] [CrossRef]
- Carvalho, T.L.A.D.B.; Nascimento, A.A.D.; Gonçalves, C.F.D.S.; Dos Santos, M.A.J.; Sales, A. Assessing the histological changes in fish gills as environmental bioindicators in Paraty and Sepetiba bays in Rio de Janeiro, Brazil. Lat. Am. J. Aquat. Res. 2020, 48, 590–601. [Google Scholar] [CrossRef]
- Shah, N.; Khan, A.; Ali, R.; Marimuthu, K.; Uddin, M.N.; Rizwan, M.; Rahman, K.U.; Alam, M.; Adnan, M.; Jawad, S.M.; et al. Monitoring Bioaccumulation (in Gills and Muscle Tissues), Hematology, and Genotoxic Alteration in Ctenopharyngodon idella Exposed to Selected Heavy Metals. BioMed Res. Int. 2020, 2020, 6185231. [Google Scholar] [CrossRef] [PubMed]
- Javid, A.; Javed, M.; Abdullah, S. Nickel bio-accumulation in the bodies of Catla catla, Labeo rohita and Cirrhina mrigala during 96-hr LC50 exposures. Int. J. Agric. Biol. 2007, 9, 139–142. [Google Scholar]
- Mallatt, J. Fish Gill Structural Changes Induced by Toxicants and Other Irritants: A Statistical Review. Can. J. Fish. Aquat. Sci. 1985, 42, 630–648. [Google Scholar] [CrossRef]
- Sultana, T.; Butt, K.; Sultana, S.; Al-Ghanim, K.A.; Mubashra, R.; Bashir, N.; Ahmed, Z.; Ashraf, A.; Mahboob, S. Histopathological Changes in Liver, Gills and Intestine of Labeo rohita Inhabiting Industrial Waste Contaminated Water of River Ravi. Pak. J. Zool. 2016, 48, 1171–1177. [Google Scholar]
- Mohamed, F.A.S. Bioaccumulation of selected metals and histopathological alterations in tissues of Oreochromis niloticus and Lates niloticus from Lake Nasser, Egypt. Global Vet. 2008, 2, 205–218. [Google Scholar]
- Hanna, M.I.; Shaheed, I.B.; Elias, N.S. A contribution on chromium and lead toxicity in cultured Oreochromis niloticus. Egypt. J. aquat. Biol. Fish. 2005, 9, 177–209. [Google Scholar]
- Salim, F. Histopathological Effect of heavy metal on different organs of fresh water fish tissues from Garmat Ali River adjacent to Al- Najebyia Power Station Kufa. J. Vet. Med. Sci. 2015, 6, 141–153. [Google Scholar]
- Giari, L.; Manera, M.; Simoni, E.; Dezfuli, B. Cellular alterations in different organs of European sea bass Dicentrarchus labrax (L.) exposed to cadmium. Chemosphere 2007, 67, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Durnam, D.M.; Hoffman, J.S.; Quaife, C.J.; Benditt, E.P.; Chen, H.Y.; Brinster, R.L.; Palmiter, R.D. Induction of mouse metallothionein-I mRNA by bacterial endotoxin is independent of metals and glucocorticoid hormones. Proc. Natl. Acad. Sci. USA 1984, 81, 1053–1056. [Google Scholar] [CrossRef] [PubMed]
- DiSilvestro, R.A.; Cousins, R.J. Glucocorticoid independent mediation of interleukin-1 induced changes in serum zinc and liver metallothionein levels. Life Sci. 1984, 35, 2113–2118. [Google Scholar] [CrossRef]
- Swerdel, M.R.; Cousins, R.J. Changes in Rat Liver Metallothionein and Metallothionein mRNA Induced by Isopropanol. Exp. Biol. Med. 1984, 175, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Del Razo, L.M.; Quintanilla-Vega, B.; Brambila-Colombres, E.; Aranda, E.S.C.; Manno, M.; Albores, A. Stress Proteins Induced by Arsenic. Toxicol. Appl. Pharmacol. 2001, 177, 132–148. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Code | Primer Sequence | GC Content (%) | Tm (°C) |
|---|---|---|---|---|
| β-actin | β-actin-F | 5-GTCCCTGTACGCCTCTGGTCG-3 | 66.7 | 61.8 |
| β-actin-R | 5-GCCGGACTCATCGTACTCCTG-3 | 61.9 | 59.4 | |
| Heat shock protein 60 | HSP 60-F | 5′-AAAGATGGTGTCACAGTTGC-3′′ | 45.0 | 53.4 |
| HSP 60-R | 5′-TGTTGAGGACCAGAGTGCTG-3 | 55.0 | 57.0 | |
| Metallothionein | MT-F | 5′-CTGCAACTGCGGAGGA-3′ | 62.5 | 55.1 |
| MT-R | 5′-GGTGTCGCATGTCTTTCCTT-3′ | 50.0 | 55.5 |
| Water Quality Parameters | Acclimatization Period | LC50 Determination Period | Final Exposure Period | Test Kit/Equipment Used |
|---|---|---|---|---|
| Temperature (°C) | 29.98 ± 0.39 | 29.35 ± 0.09 | 29.18 ± 0.02 | HACH pH Meter, thermometer |
| PH | 7.7 ± 0.05 | 8.32 ± 0.45 | 8.19 ± 0.02 | HACH pH Meter |
| Dissolved oxygen (ppm) | 5.98 ± 0.21 | 7.57 ± 0.11 | 7.42 ± 0.24 | HACH HQ30d Flexi DO Meter |
| Salinity (ppt) | 0.15 ± 0.02 | 0.12 ± 0.01 | 0.15 ± 0.03 | Horiba U-50 series multiparameter |
| Conductivity (mS/cm) | 0.28 ± 0.01 | 0.28 ± 0.01 | 0.32 ± 0.05 | Horiba U-50 series multiparameter |
| Ammonia (ppm) | <0.25 | <0.25 | <0.25 | API fresh water master test kit. |
| ORP (Mv) | 105.50 ± 0.50 | 105.33 ± 0.88 | 177.33 ± 2.84 | Horiba U-50 series multiparameter |
| Nitrate (ppm) | <0.25 | <0.25 | <0.25 | API fresh water master test kit |
| TDS (g/L) | 0.20 ± 0.04 | 0.25 ± 0.13 | 0.20 ± 0.03 | Horiba U-50 series multiparameter |
| Nitrite (ppm) | <0.50 | <0.50 | <0.50 | API fresh water master test kit |
| Turbidity (NTU) | 26.3 ± 22.0 | 9.911 ± 0.06 | 26.03 ± 19.39 | Horiba U-50 series multiparameter |
| Growth Parameters | Control | T1 | T2 | T3 |
|---|---|---|---|---|
| Weight gain (g) | 0.18 ± 0.04 ab | 0.09 ± 0.05 b | 0.16 ± 0.06 b | −0.05 ± 0.07 bc |
| Length gain (cm) | 0.58 ± 0.11 a | 0.37 ± 0.11 a | 0.45 ± 0.19 a | −0.35 ± 0.12 b |
| Condition factor (initial) | 0.98 ± 0.07 a | 0.93 ± 0.05 a | 0.97 ± 0.05 a | 0.82 ± 0.03 a |
| Condition factor (final) | 0.89 ± 0.03 a | 0.86 ± 0.04 a | 0.95 ± 0.06 a | 0.92 ± 0.07 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabbane, M.G.; Kabir, M.A.; Habibullah-Al-Mamun, M.; Mustafa, M.G. Toxic Effects of Arsenic in Commercially Important Fish Rohu Carp, Labeo rohita of Bangladesh. Fishes 2022, 7, 217. https://doi.org/10.3390/fishes7050217
Rabbane MG, Kabir MA, Habibullah-Al-Mamun M, Mustafa MG. Toxic Effects of Arsenic in Commercially Important Fish Rohu Carp, Labeo rohita of Bangladesh. Fishes. 2022; 7(5):217. https://doi.org/10.3390/fishes7050217
Chicago/Turabian StyleRabbane, Md. Golam, Md. Alamgir Kabir, Md. Habibullah-Al-Mamun, and Md. Ghulam Mustafa. 2022. "Toxic Effects of Arsenic in Commercially Important Fish Rohu Carp, Labeo rohita of Bangladesh" Fishes 7, no. 5: 217. https://doi.org/10.3390/fishes7050217
APA StyleRabbane, M. G., Kabir, M. A., Habibullah-Al-Mamun, M., & Mustafa, M. G. (2022). Toxic Effects of Arsenic in Commercially Important Fish Rohu Carp, Labeo rohita of Bangladesh. Fishes, 7(5), 217. https://doi.org/10.3390/fishes7050217