
Differential Effects of Food Restriction and Warming in the Two-Spotted Goby: Impaired Reproductive Performance and Stressed Offspring

 , , , ,
, , , , 


Abstract
:1. Introduction
2. Materials and Methods
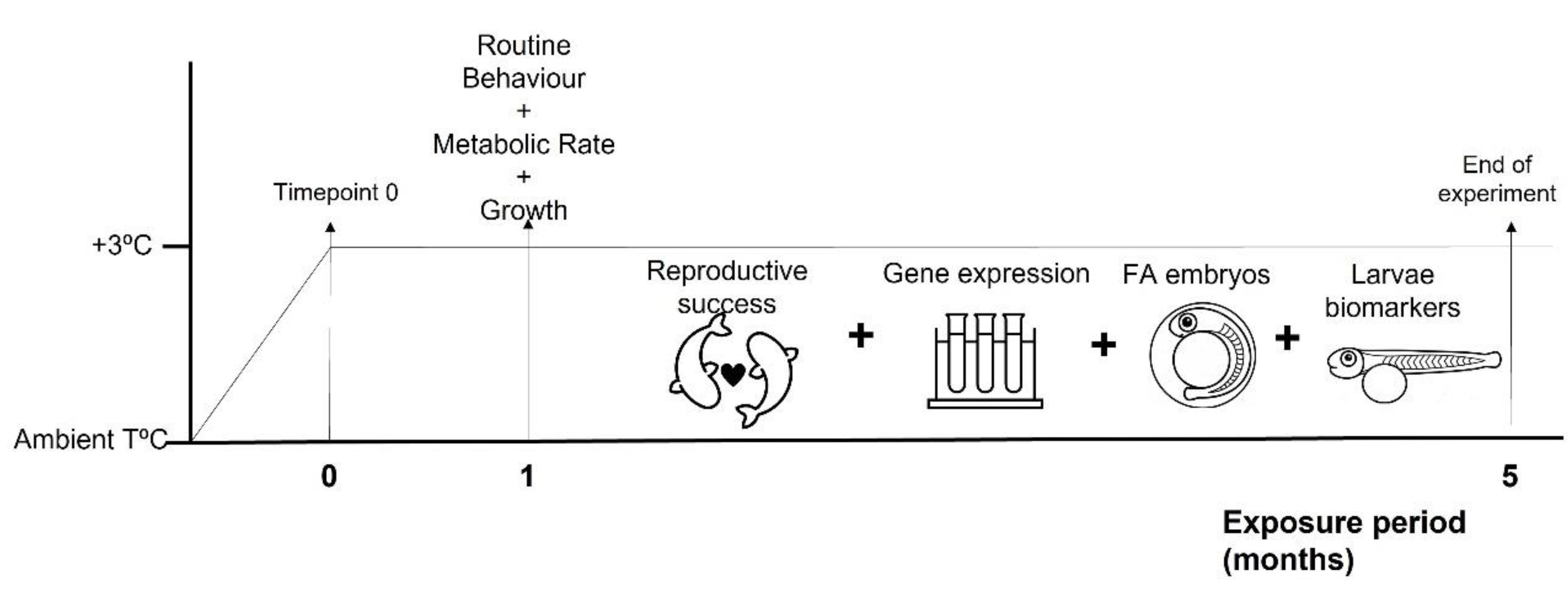
2.1. Fish Collection and Experimental Setup
2.2. Temperature and Food Availability Effects on Behaviour and Physiology
2.2.1. Routine Behaviour
2.2.2. Routine Metabolic Rate
2.3. Temperature and Food Availability Effects on Reproduction
2.3.1. Reproductive Success
2.3.2. Gene Expression of Reproduction-Related Genes
2.3.3. Fatty Acids Content in Recently Fertilized Eggs
2.3.4. Biomarker Analysis in Recently Hatched Larvae
2.4. Statistical Analysis
3. Results
3.1. Temperature and Food Availability Effects on Behaviour and Physiology
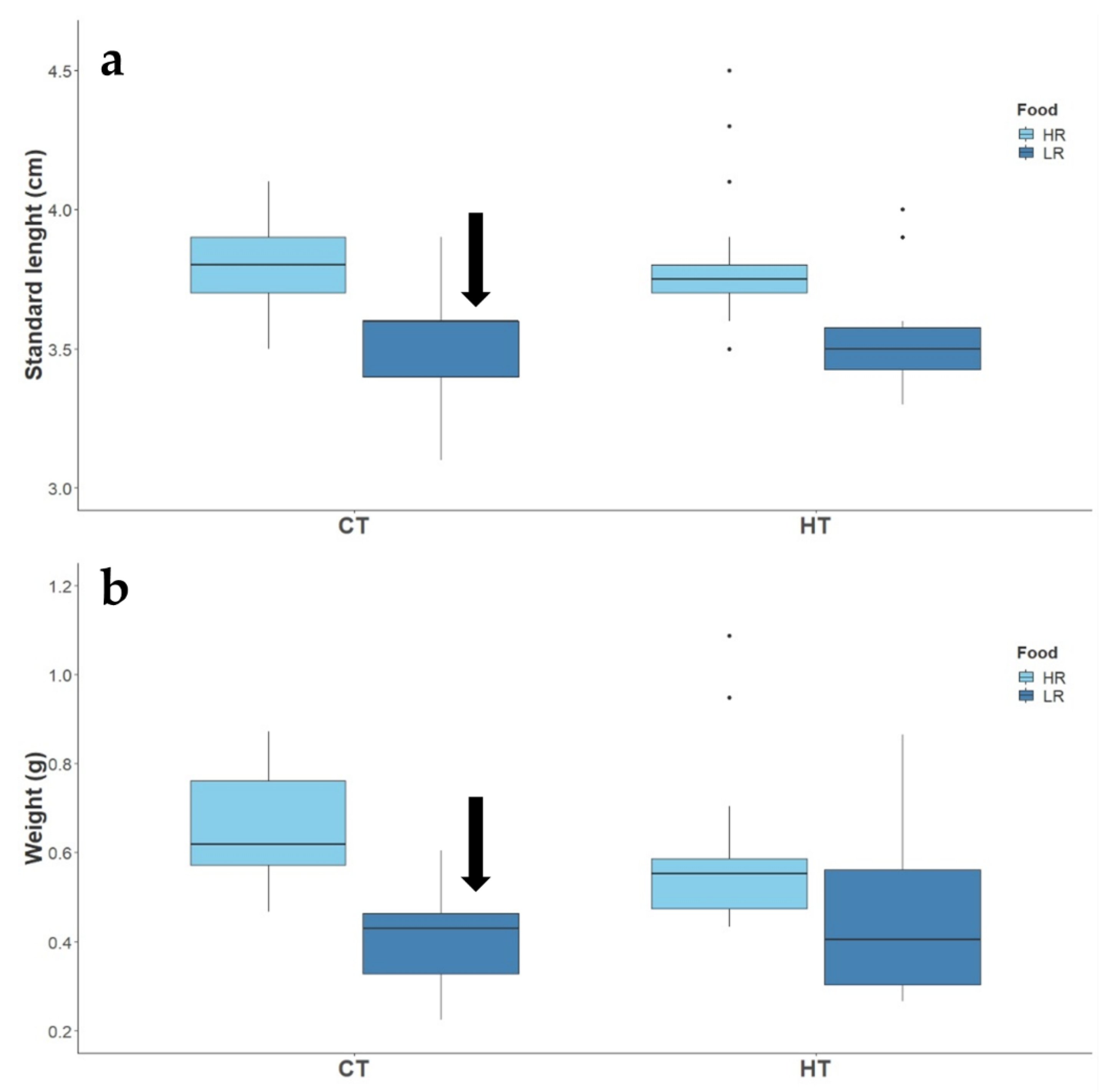
3.1.1. Routine Activity
3.1.2. Metabolic Rate
3.2. Temperature and Food Availability Effects on Reproduction
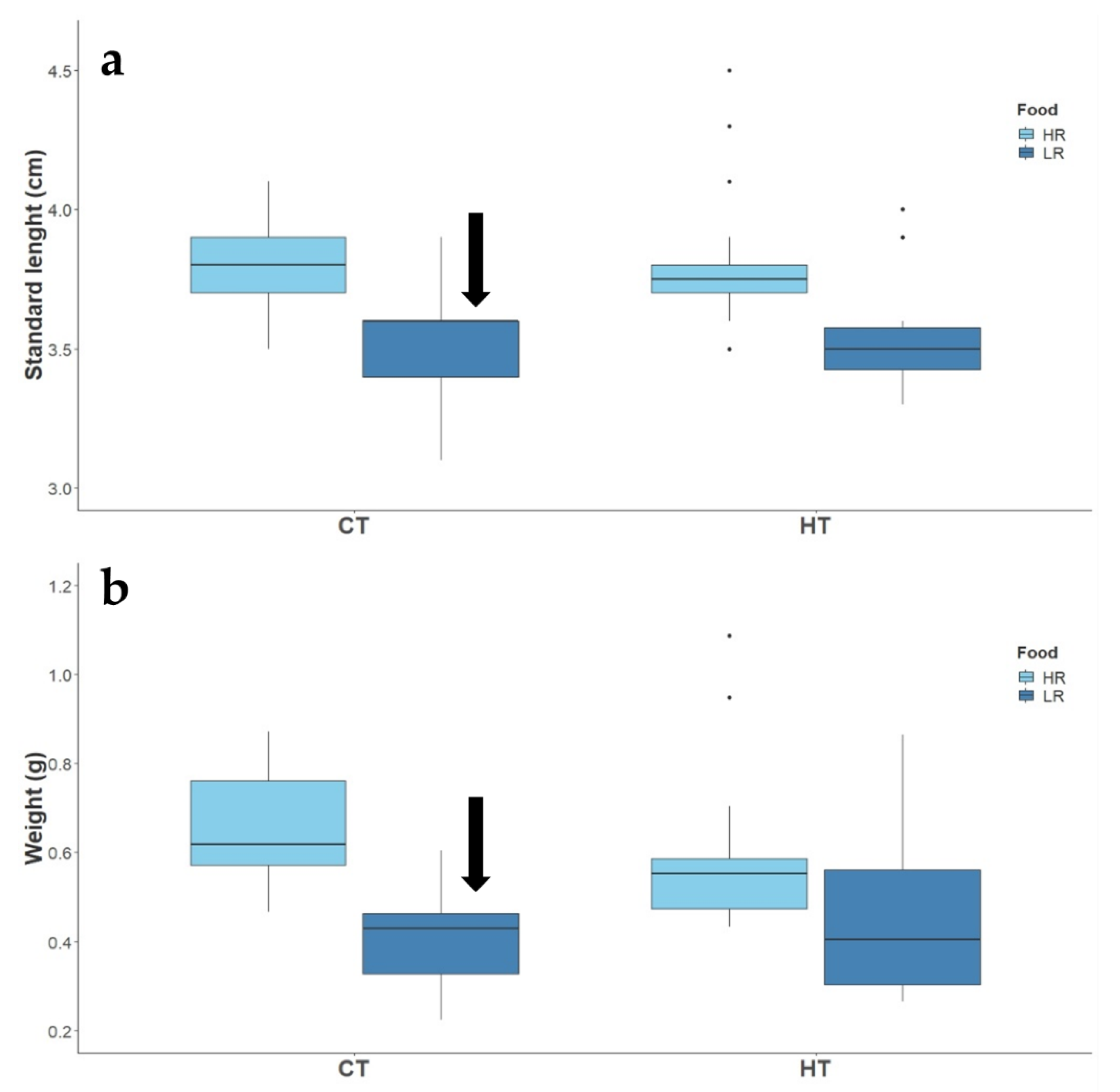
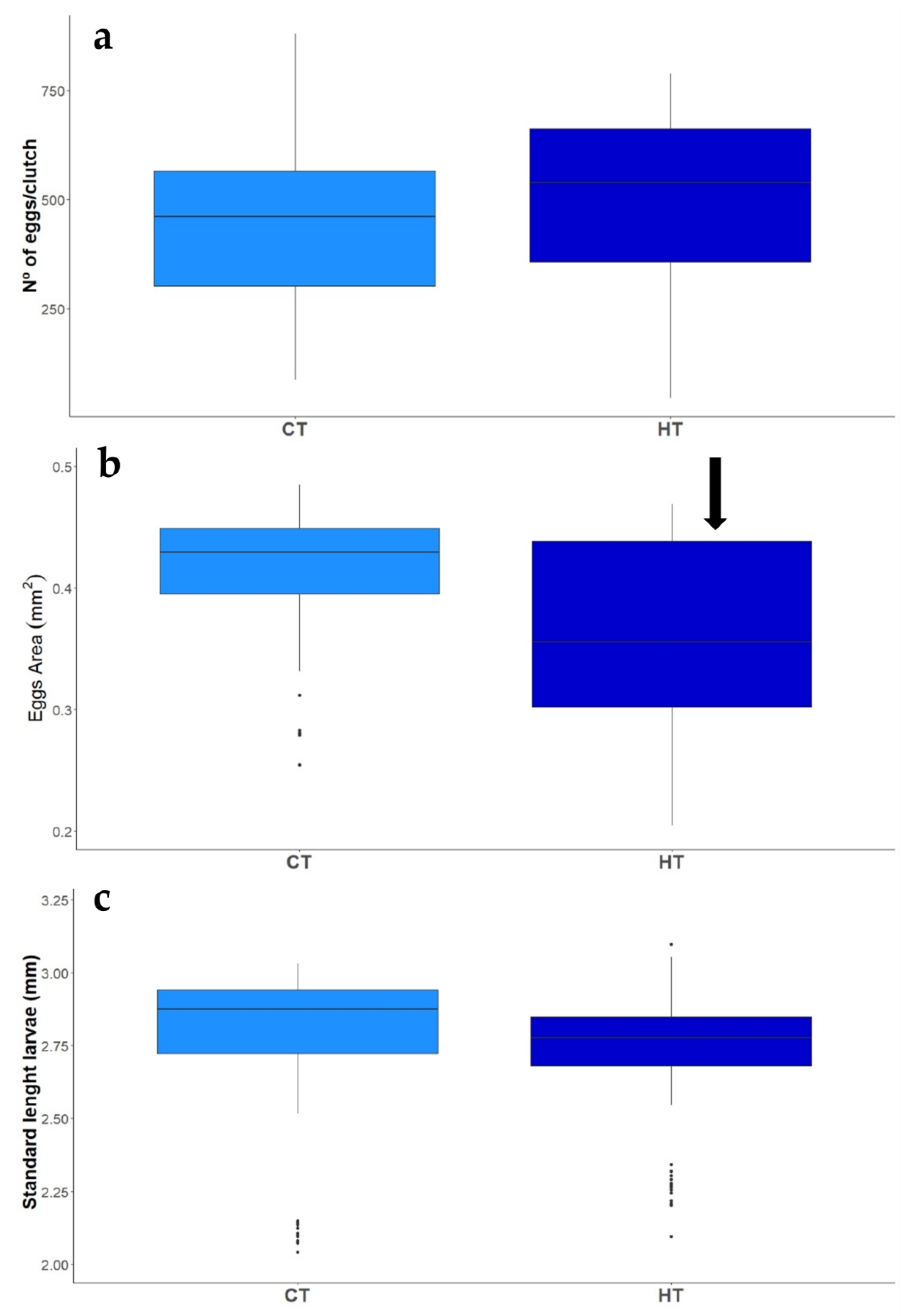
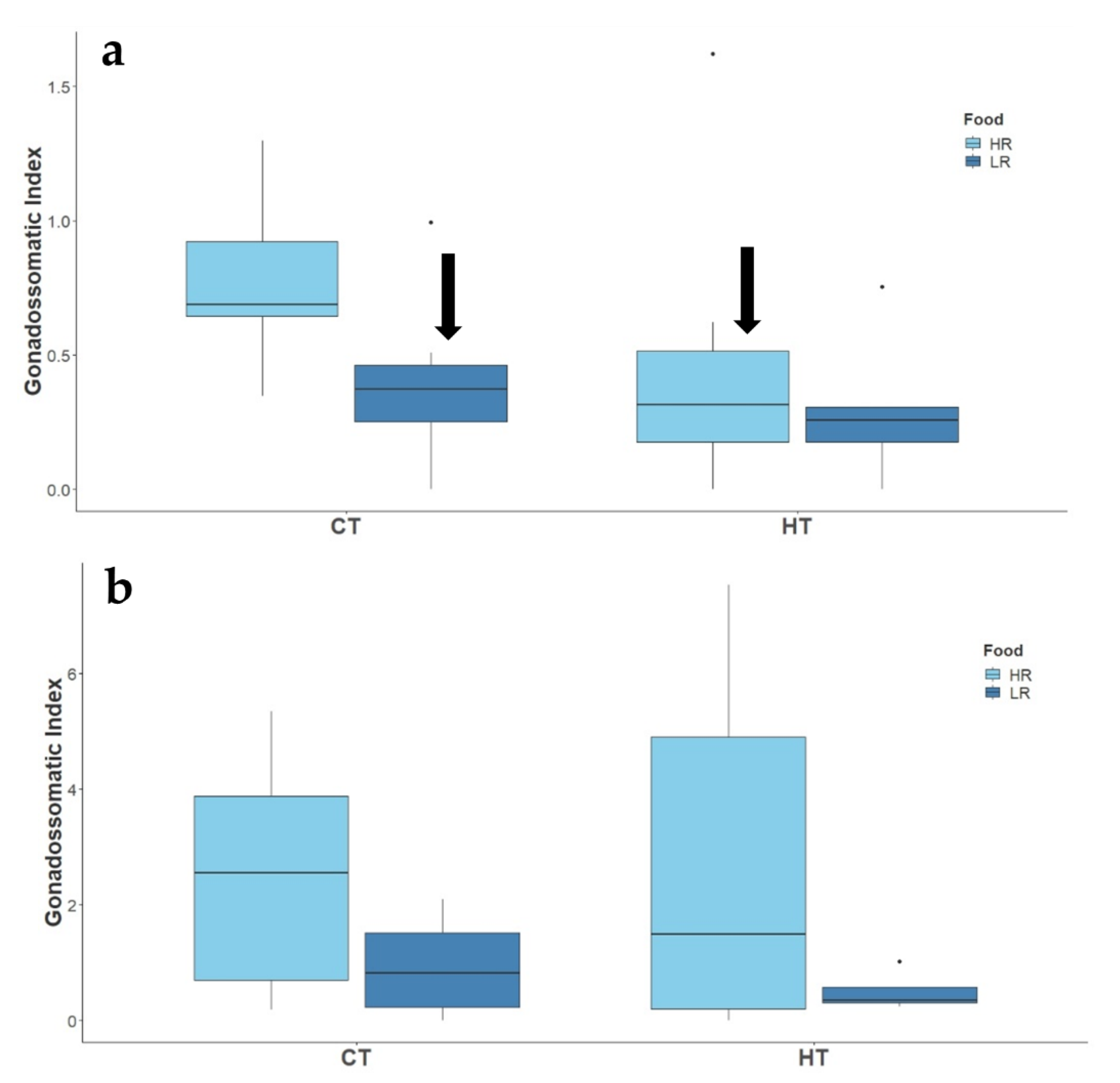
3.2.1. Reproductive Success
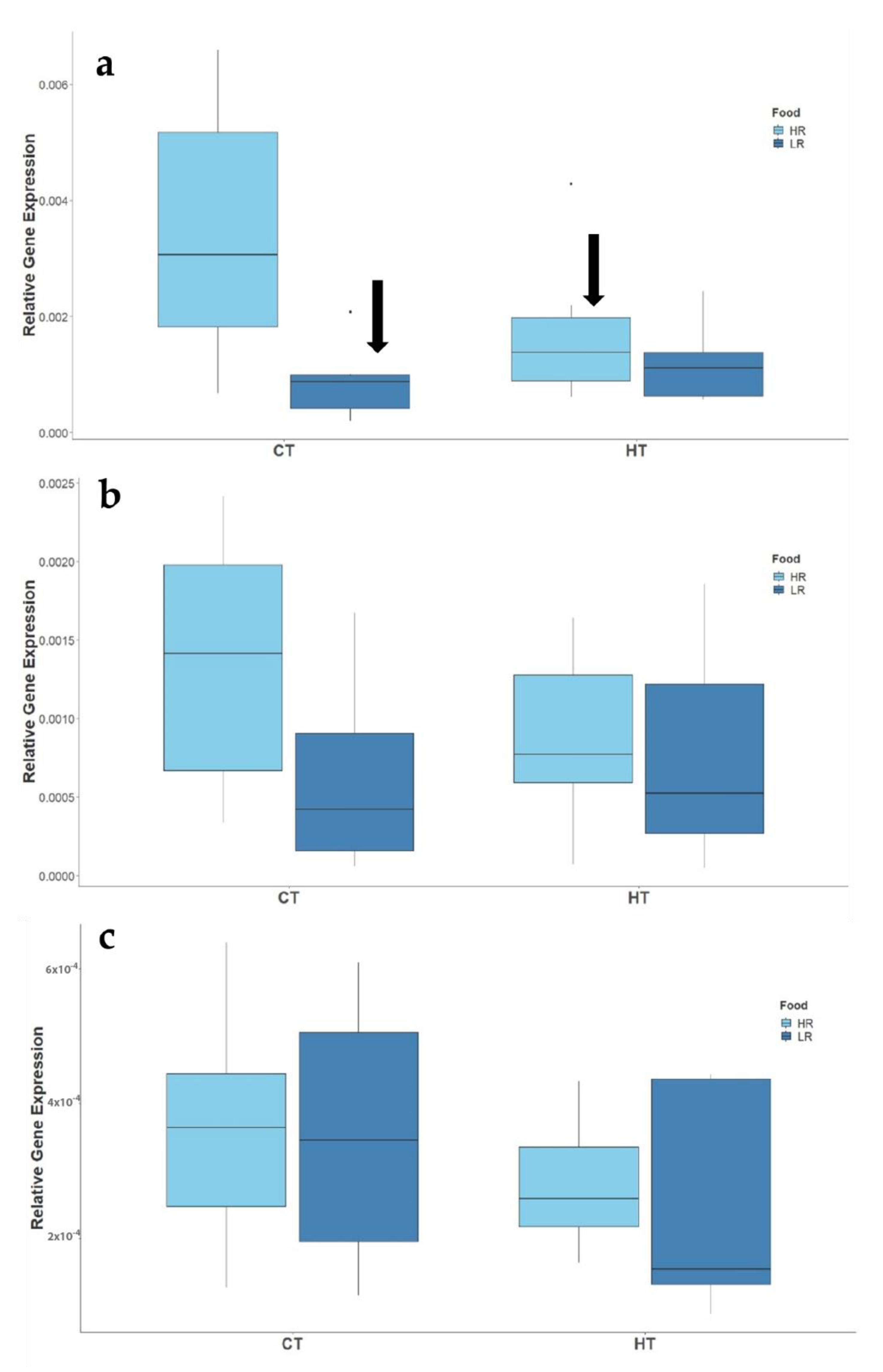
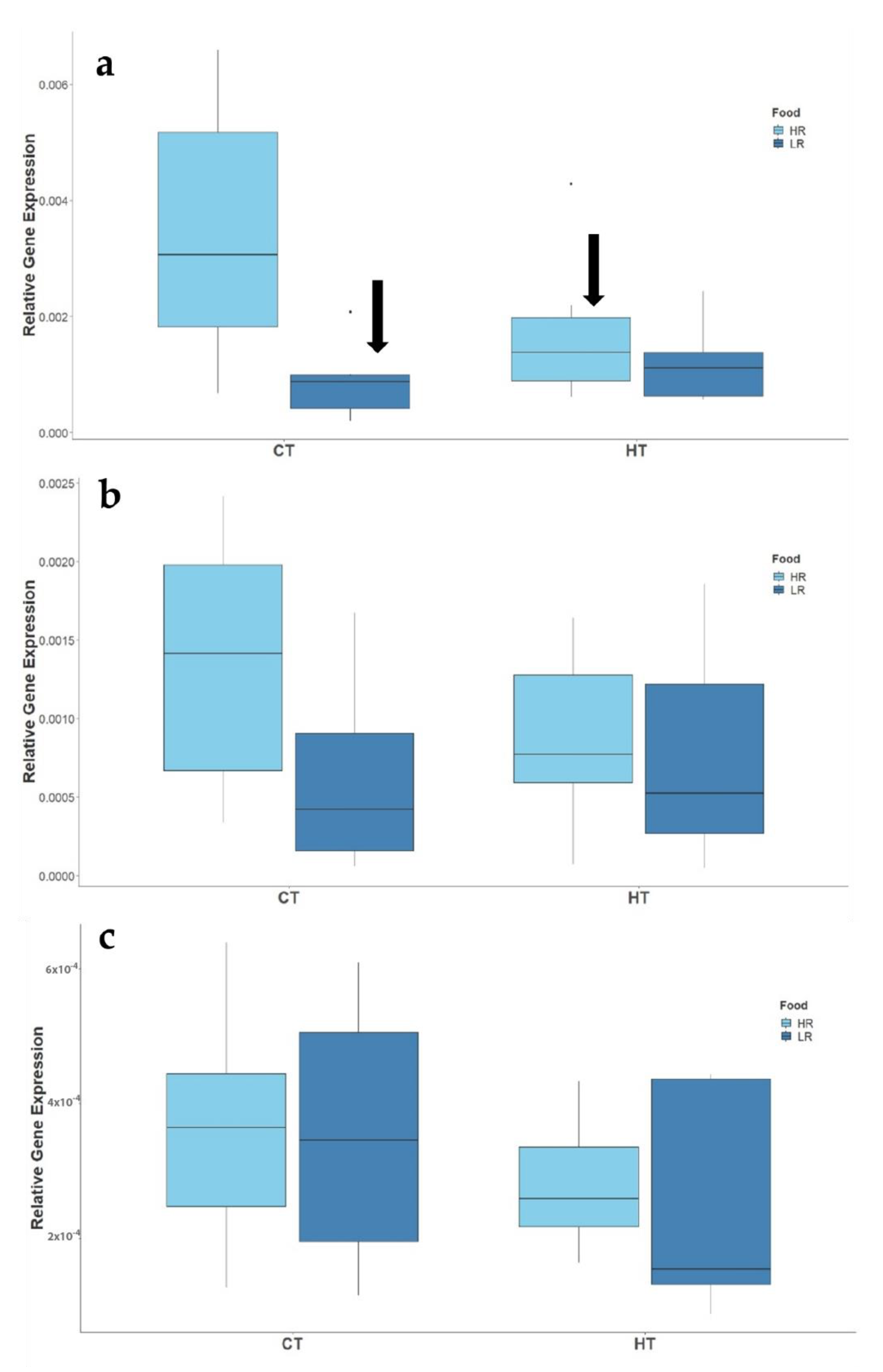
3.2.2. Expression of Reproduction-Related Genes
3.2.3. Fatty Acid Content
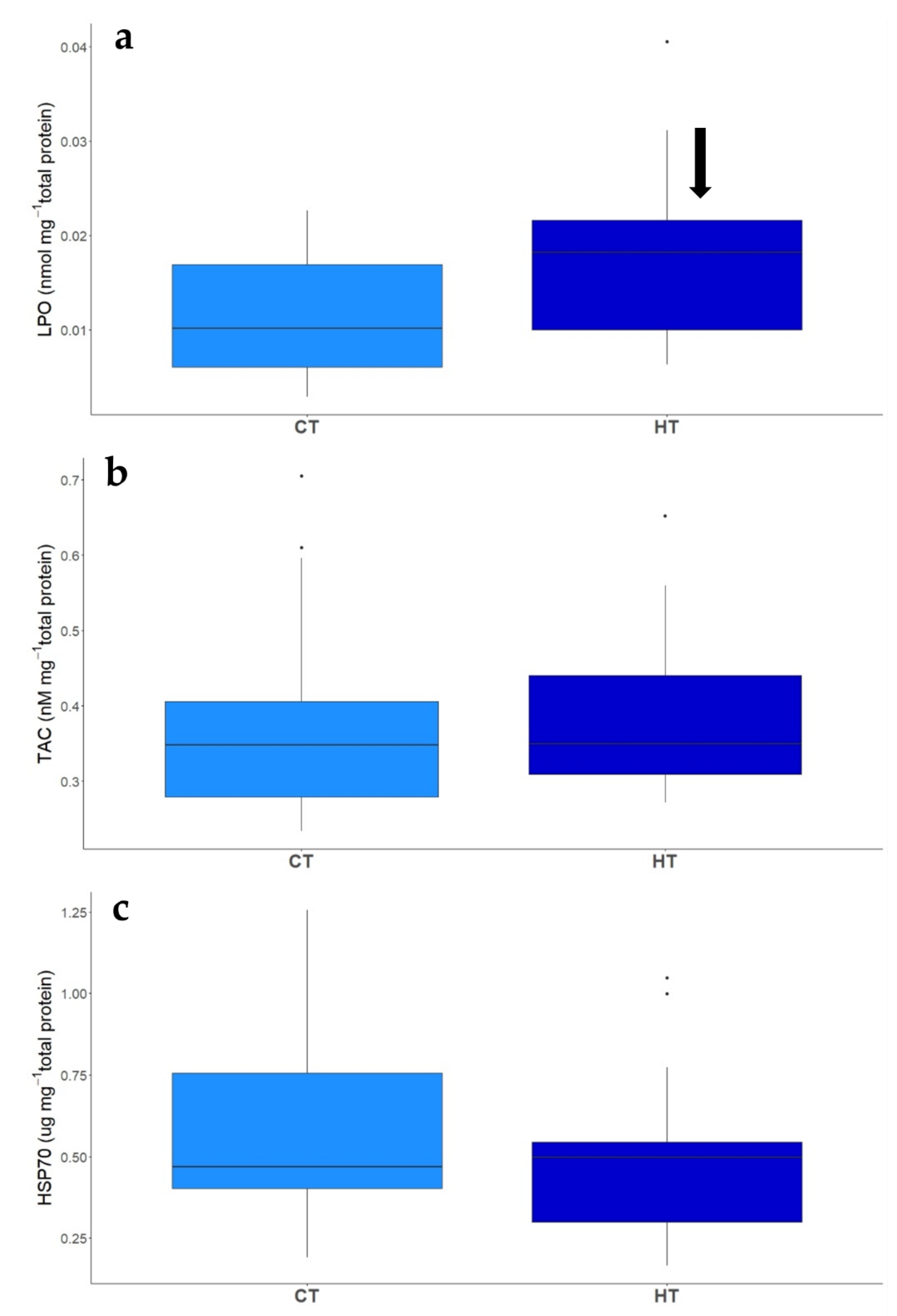
3.2.4. Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Djurichkovic, L.D.; Donelson, J.M.; Fowler, A.M.; Feary, D.A.; Booth, D.J. The Effects of Water Temperature on the Juvenile Performance of Two Tropical Damselfishes Expatriating to Temperate Reefs. Sci. Rep. 2019, 9, 13937. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.T.; Donelson, J.M.; McCormick, M.I. Extended Exposure to Elevated Temperature Affects Escape Response Behaviour in Coral Reef Fishes. PeerJ 2017, 2017, 5, e3652. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, G.E.; Crawley, N.; Lunde, I.G.; Munday, P.L. Elevated Temperature Reduces the Respiratory Scope of Coral Reef Fishes. Glob. Chang. Biol. 2009, 15, 1405–1412. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Mendonça, V.; Dias, M.; Roma, J.; Diniz, M.S. Effect of Increasing Temperature in the Differential Activity of Oxidative Stress Biomarkers in Various Tissues of the Rock Goby, Gobius Paganellus. Mar. Environ. Res. 2014, 97, 10–14. [Google Scholar] [CrossRef]
- Lopes, A.F.; Faria, A.M.; Dupont, S. Elevated Temperature, but Not Decreased PH, Impairs Reproduction in a Temperate Fish. Sci. Rep. 2020, 10, 20805. [Google Scholar] [CrossRef]
- Hopkins, K.; Moss, B.R.; Gill, A.B. Increased Ambient Temperature Alters the Parental Care Behaviour and Reproductive Success of the Three-Spined Stickleback (Gasterosteus Aculeatus). Environ. Biol. Fishes 2011, 90, 121–129. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Munday, P.L. Effects of Climate Change on Fish Reproduction and Early Life History Stages. Mar. Freshw. Res. 2011, 62, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Sswat, M.; Stiasny, M.H.; Jutfelt, F.; Riebesell, U.; Clemmesen, C. Growth Performance and Survival of Larval Atlantic Herring, under the Combined Effects of Elevated Temperatures and CO2. PLoS ONE 2018, 13, e0191947. [Google Scholar] [CrossRef] [Green Version]
- Fox-Kemper, B.; Hewitt, H.T.; Xiao, C.; Aðalgeirsdóttir, G.; Drijfhout, S.S.; Edwards, T.L.; Golledge, N.R.; Hemer, M.; Kopp, R.E.; Krinner, G.; et al. Ocean, Cryosphere and Sea Level Climate Change 2021: The Physical Science Basis Contribution Working Group I to Sixth Assessment. Rep. Intergov. Panel Clim. 2021, 2018, 1–257. [Google Scholar]
- IPCC. Assessment Report 6 Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; IPCC: Geneva, Switzerland, 2021; In Press. [Google Scholar]
- Deslauriers, D.; Kieffer, J.D. The Effects of Temperature on Swimming Performance of Juvenile Shortnose Sturgeon (Acipenser Brevirostrum). J. Appl. Ichthyol. 2012, 28, 176–181. [Google Scholar] [CrossRef]
- Milazzo, M.; Mirto, S.; Domenici, P.; Gristina, M. Climate Change Exacerbates Interspecific Interactions in Sympatric Coastal Fishes. J. Anim. Ecol. 2013, 82, 468–477. [Google Scholar] [CrossRef]
- Laubenstein, T.D.; Rummer, J.L.; Nicol, S.; Parsons, D.M.; Pether, S.M.J.; Pope, S.; Smith, N.; Munday, P.L. Correlated Effects of Ocean Acidification and Warming on Behavioral and Metabolic Traits of a Large Pelagic Fish. Diversity 2018, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.J.; Feary, D.A.; Fowler, A.M.; Madin, E.M.P.; Booth, D.J. Temperate Predators and Seasonal Water Temperatures Impact Feeding of a Range Expanding Tropical Fish. Mar. Biol. 2016, 163, 70. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Munday, P.L.; Rummer, J.L.; McCormick, M.I.; Corkill, K.; Watson, S.-A.; Allan, B.J.M.; Meekan, M.G.; Chivers, D.P. Interactive Effects of Ocean Acidification and Rising Sea Temperatures Alter Predation Rate and Predator Selectivity in Reef Fish Communities. Glob. Chang. Biol. 2015, 21.5, 1848–1855. [Google Scholar] [CrossRef]
- Allan, B.J.M.; Domenici, P.; Watson, S.A.; Munday, P.L.; McCormick, M.I. Warming Has a Greater Effect than Elevated CO2 on Predator–Prey Interactions in Coral Reef Fish. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170784. [Google Scholar] [CrossRef] [Green Version]
- Servili, A.; Canario, A.V.M.; Mouchel, O.; Muñoz-Cueto, J.A. Climate Change Impacts on Fish Reproduction Are Mediated at Multiple Levels of the Brain-Pituitary-Gonad Axis. Gen. Comp. Endocrinol. 2020, 291, 113439. [Google Scholar] [CrossRef]
- Alix, M.; Kjesbu, O.S.; Anderson, K.C. From Gametogenesis to Spawning: How Climate-Driven Warming Affects Teleost Reproductive Biology. J. Fish Biol. 2020, 97, 607–632. [Google Scholar] [CrossRef]
- Miller, G.M.; Kroon, F.J.; Metcalfe, S.; Munday, P. Temperature Is the Evil Twin: Effects of Increased Temperature and Ocean Acidification on Reproduction in a Reef Fish. Ecol. Appl. 2015, 25, 603–620. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A. The Effect of Ocean Acidification and Temperature on the Fertilization and Embryonic Development of the Sydney Rock Oyster Saccostrea Glomerata (Gould 1850). Glob. Chang. Biol. 2009, 15, 2123–2136. [Google Scholar] [CrossRef]
- Rajakumar, A.; Senthilkumaran, B. Steroidogenesis and Its Regulation in Teleost-a Review. Fish Physiol. Biochem. 2020, 46, 803–818. [Google Scholar] [CrossRef]
- Strüssmann, C.A.; Conover, D.O.; Somoza, G.M.; Miranda, L.A. Implications of Climate Change for the Reproductive Capacity and Survival of New World Silversides (Family Atherinopsidae). J. Fish Biol. 2010, 77, 1818–1834. [Google Scholar] [CrossRef]
- Sandra, G.E.; Norma, M.M. Sexual Determination and Differentiation in Teleost Fish. Rev. Fish Biol. Fish. 2010, 20, 101–121. [Google Scholar] [CrossRef]
- Díaz, N.; Piferrer, F. Lasting Effects of Early Exposure to Temperature on the Gonadal Transcriptome at the Time of Sex Differentiation in the European Sea Bass, a Fish with Mixed Genetic and Environmental Sex Determination. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Donelson, J.M.; Munday, P.L.; McCormick, M.I. Climate Change May Affect Fish through an Interaction of Parental and Juvenile Environments. Coral Reefs 2012, 31, 753–762. [Google Scholar] [CrossRef]
- Frenette, B.D.; Bruckerhoff, L.A.; Tobler, M.; Gido, K.B. Temperature Effects on Performance and Physiology of Two Prairie Stream Minnows. Conserv. Physiol. 2019, 7, coz063. [Google Scholar] [CrossRef]
- Slesinger, E.; Andres, A.; Young, R.; Seibel, B.; Saba, V.; Phelan, B.; Rosendale, J.; Wieczorek, D.; Saba, G. Correction: The effect of ocean warming on black sea bass (Centropristis striata) aerobic scope and hypoxia tolerance. PLoS ONE 2020, 15, e0244002. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative Stress in Marine Environments: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Imbs, A.B.; Yakovleva, I.M. Dynamics of Lipid and Fatty Acid Composition of Shallow-Water Corals under Thermal Stress: An Experimental Approach. Coral Reefs 2012, 31, 41–53. [Google Scholar] [CrossRef]
- Oku, H.; Yamashiro, H.; Onaga, K.; Sakai, K.; Iwasaki, H. Seasonal Changes in the Content and Composition of Lipids in the Coral Goniastrea Aspera. Coral Reefs 2003, 22, 83–85. [Google Scholar] [CrossRef]
- Parrish, C.C. Lipids in Marine Ecosystems. ISRN Oceanogr. 2013, 2013, 604045. [Google Scholar] [CrossRef] [Green Version]
- Imbs, A.B.; Latyshev, N.A.; Dautova, T.N.; Latypov, Y.Y. Distribution of Lipids and Fatty Acids in Corals by Their Taxonomic Position and Presence of Zooxanthellae. Mar. Ecol. Prog. Ser. 2010, 409, 65–75. [Google Scholar] [CrossRef]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty Acid Trophic Markers in the Pelagic Marine Environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, T. Iso- and Anteiso-Fatty Acids in Bacteria: Biosynthesis, Function, and Taxonomic Significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Mortillaro, J.M.; Pitt, K.A.; Lee, S.Y.; Meziane, T. Light Intensity Influences the Production and Translocation of Fatty Acids by Zooxanthellae in the Jellyfish Cassiopea Sp. J. Exp. Mar. Bio. Ecol. 2009, 378, 22–30. [Google Scholar] [CrossRef]
- Drazen, J.C.; Phleger, C.F.; Guest, M.A.; Nichols, P.D. Lipid, Sterols and Fatty Acid Composition of Abyssal Holothurians and Ophiuroids from the North-East Pacific Ocean: Food Web Implications. Comp. Biochem. Physiol. -B Biochem. Mol. Biol. 2008, 151, 79–87. [Google Scholar] [CrossRef]
- Patton, J.S.; Battey, J.F.; Rigler, M.W.; Porter, J.W.; Black, C.C.; Burris, J.E. A Comparison of the Metabolism of Bicarbonate 14C and Acetate 1-14C and the Variability of Species Lipid Compositions in Reef Corals. Mar. Biol. 1983, 75, 121–130. [Google Scholar] [CrossRef]
- Nyboer, E.A.; Chapman, L.J. Elevated Temperature and Acclimation Time Affect Metabolic Performance in the Heavily Exploited Nile Perch of Lake Victoria. J. Exp. Biol. 2017, 220, 3782–3793. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Richardson, A.; Robinson, C. Climate Change and Marine Plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Richardson, A.J. In Hot Water: Zooplankton and Climate Change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Garzke, J.; Ismar, S.M.H.; Sommer, U. Climate Change Affects Low Trophic Level Marine Consumers: Warming Decreases Copepod Size and Abundance. Oecologia 2015, 177, 849–860. [Google Scholar] [CrossRef]
- Hixson, S.M.; Arts, M.T. Climate Warming Is Predicted to Reduce Omega-3, Long-Chain, Polyunsaturated Fatty Acid Production in Phytoplankton. Glob. Chang. Biol. 2016, 22, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, J. An Introduction to Dynamic Energy Budget (DEB) Models with Special Emphasis on Parameter Estimation. J. Sea Res. 2006, 56, 85–102. [Google Scholar] [CrossRef]
- Green, B.S. Chapter 1 Maternal Effects in Fish Populations. Adv. Mar. Biol. 2008, 54, 1–105. [Google Scholar] [CrossRef]
- Lienart, G.D.H.; Mitchell, M.D.; Ferrari, M.C.O.; McCormick, M.I. Temperature and Food Availability Affect Risk Assessment in an Ectotherm. Anim. Behav. 2014, 89, 199–204. [Google Scholar] [CrossRef]
- Liu, S.; Fu, S.J. Correction: Effects of Food Availability on Metabolism, Behaviour, Growth and Their Relationships in a Triploid Carp. J. Exp. Biol. 2018, 221, jeb187302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donelson, J.; Munday, P.; McCormick, M.; Pankhurst, N.; Pankhurst, P. Effects of Elevated Water Temperature and Food Availability on the Reproductive Performance of a Coral Reef Fish. Mar. Ecol. Prog. Ser. 2010, 401, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.; Lopes, A.R.; Ribeiro, L.; Castanho, S.; Candeias-mendes, A.; Pousão-ferreira, P.; Faria, A.M. Effects of Exposure to Elevated Temperature and Different Food Levels on the Escape Response and Metabolism of Early Life Stages of White Seabream, Diplodus Sargus. Conserv. Physiol. 2022, 10, coac023. [Google Scholar] [CrossRef]
- Amundsen, T. Sex Roles and Sexual Selection: Lessons from a Dynamic Model System. Curr. Zool. 2018, 64, 363–392. [Google Scholar] [CrossRef]
- Borg, Å.A.; Forsgren, E.; Amundsen, T. Seasonal Change in Female Choice for Male Size in the Two-Spotted Goby. Anim. Behav. 2006, 72, 763–771. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour. In Measuring Behaviour; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar] [CrossRef]
- Killen, S.S.; Mitchell, M.D.; Rummer, J.L.; Chivers, D.P.; Ferrari, M.C.O.; Meekan, M.G.; Mccormick, M.I. Aerobic Scope Predicts Dominance during Early Life in a Tropical Damselfish. Funct. Ecol. 2014, 28, 1367–1376. [Google Scholar] [CrossRef]
- Song, Y.F.; Huang, C.; Shi, X.; Pan, Y.X.; Liu, X.; Luo, Z. Endoplasmic Reticulum Stress and Dysregulation of Calcium Homeostasis Mediate Cu-Induced Alteration in Hepatic Lipid Metabolism of Javelin Goby Synechogobius Hasta. Aquat. Toxicol. 2016, 175, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Leder, E.H.; André, C.; Le Moan, A.; Töpel, M.; Blomberg, A.; Havenhand, J.N.; Lindström, K.; Volckaert, F.A.M.; Kvarnemo, C.; Johannesson, K.; et al. Post-Glacial Establishment of Locally Adapted Fish Populations over a Steep Salinity Gradient. J. Evol. Biol. 2021, 34, 138–156. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and Modifications of Primer Design Program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Kalendar, R.; Khassenov, B.; Ramankulov, Y.; Samuilova, O.; Ivanov, K.I. FastPCR: An in Silico Tool for Fast Primer and Probe Design and Advanced Sequence Analysis. Genomics 2017, 109, 312–319. [Google Scholar] [CrossRef]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. HSP70 Production Patterns in Coastal and Estuarine Organisms Facing Increasing Temperatures. J. Sea Res. 2012, 73, 137–147. [Google Scholar] [CrossRef]
- Mukhopadhyay, I.; Nazir, A.; Saxena, D.K.; Kar Chowdhuri, D. Heat Shock Response: Hsp70 in Environmental Monitoring. J. Biochem. Mol. Toxicol. 2003, 17, 249–254. [Google Scholar] [CrossRef]
- Yamashita, M.; Yabu, T.; Ojima, N. Stress Protein HSP70 in Fish. Aqua-BioScience Monogr. 2010, 3, 111–141. [Google Scholar] [CrossRef]
- Kambayashi, Y.; Binh, N.T.; Asakura, H.W.; Hibino, Y.; Hitomi, Y.; Nakamura, H.; Ogino, K. Efficient Assay for Total Antioxidant Capacity in Human Plasma Using a 96-Well Microplte. J. Clin. Biochem. Nutr. 2009, 44, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J. A New Method for Measuring Antioxidant Activity. Biochem. Soc. Trans. 1993, 21, 95S. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, M.; Mihara, M. Determination of Malonaldehyde Precursor in Tissues by Thiobarbituric Acid Test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Jain, R.B. A Recursive Version of Grubbs’ Test for Detecting Multiple Outliers in Environmental and Chemical Data. Clin. Biochem. 2010, 43, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Ballew, N.G.; Kjelvik, M.K. Fish Behavioral Types and Their Ecological Consequences. Can. J. Fish. Aquat. Sci. 2014, 71, 927–944. [Google Scholar] [CrossRef]
- Ojanguren, A.F.; Braña, F. Thermal Dependence of Swimming Endurance in Juvenile Brown Trout. J. Fish Biol. 2000, 56, 1342–1347. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. State-Dependent Behavior and Alternative Behavioral Strategies in Brown Trout (Salmo trutta L.) Fry. Behav. Ecol. Sociobiol. 2016, 70, 2111–2125. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.T.; Donelson, J.M.; McCormick, M.I.; Ferrari, M.C.O.; Munday, P.L. Duration of Exposure to Elevated Temperature Affects Competitive Interactions in Juvenile Reef Fishes. PLoS ONE 2016, 11, e0164505. [Google Scholar] [CrossRef] [Green Version]
- Lehtiniemi, M. Swim or Hide: Predator Cues Cause Species Specific Reactions in Young Fish Larvae. J. Fish Biol. 2005, 66, 1285–1299. [Google Scholar] [CrossRef]
- Killen, S.S.; Marras, S.; Mckenzie, D.J. Fuel, Fasting, Fear: Routine Metabolic Rate and Food Deprivation Exert Synergistic Effects on Risk-Taking in Individual Juvenile European Sea Bass. J. Anim. Ecol. 2011, 80, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Aimon, C.; Le Bayon, N.; Le Floch, S.; Claireaux, G. Food Deprivation Reduces Social Interest in the European Sea Bass Dicentrarchus Labrax. J. Exp. Biol. 2019, 222, jeb190553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, R.E.; Johnson, D.W. Metabolic Responses to Temperature in a Sedentary Reef Fish, the Bluebanded Goby (Lythrypnus dalli, Gilbert). J. Exp. Mar. Bio. Ecol. 2018, 501, 83–89. [Google Scholar] [CrossRef]
- Di Santo, V.; Bennett, W.A. Effect of Rapid Temperature Change on Resting Routine Metabolic Rates of Two Benthic Elasmobranchs. Fish Physiol. Biochem. 2011, 37, 929–934. [Google Scholar] [CrossRef]
- McLeod, I.M.; Rummer, J.L.; Clark, T.D.; Jones, G.P.; McCormick, M.I.; Wenger, A.S.; Munday, P.L. Climate Change and the Performance of Larval Coral Reef Fishes: The Interaction between Temperature and Food Availability. Conserv. Physiol. 2013, 1, cot024. [Google Scholar] [CrossRef] [Green Version]
- Killen, S.S.; Marras, S.; Ryan, M.R.; Domenici, P.; Mckenzie, D.J. A Relationship between Metabolic Rate and Risk-Taking Behaviour Is Revealed during Hypoxia in Juvenile European Sea Bass. Funct. Ecol. 2012, 26, 134–143. [Google Scholar] [CrossRef]
- Donelson, J.M.; Wong, M.; Booth, D.J.; Munday, P.L. Transgenerational Plasticity of Reproduction Depends on Rate of Warming across Generations. Evol. Appl. 2016, 9, 1072–1081. [Google Scholar] [CrossRef]
- Tenugu, S.; Pranoty, A.; Mamta, S.-K.; Senthilkumaran, B. Development and Organisation of Gonadal Steroidogenesis in Bony Fishes—A Review. Aquac. Fish. 2021, 6, 223–246. [Google Scholar] [CrossRef]
- Idler, D.R.; Bitners, I.I.; Schmidt, P.J. 11-Ketotestosterone: And androgen for Sockeye Salmon Can. J. Biochem. Physioogyl. 2011, 39, 1737–1742. [Google Scholar] [CrossRef]
- Ueda, H.; Kambegawa, A.; Nagahama, Y. Involvement of Gonadotrophin and Steroid Hormones in Spermiation in the Amago Salmon, Oncorhynchus Rhodurus, and Goldfish, Carassius Auratus. Gen. Comp. Endocrinol. 1985, 59, 24–30. [Google Scholar] [CrossRef]
- Yoneda, M.; Wright, P.J. Effect of Temperature and Food Availability on Reproductive Investment of First-Time Spawning Male Atlantic Cod, Gadus Morhua. ICES J. Mar. Sci. 2005, 62, 1387–1393. [Google Scholar] [CrossRef]
- Mayer, I.; Borg, B.; Schulz, R. Seasonal Changes in and Effect of Castration/Androgen Replacement on the Plasma Levels of Five Androgens in the Male Three-Spined Stickleback, Gasterosteus Aculeatus L. Gen. Comp. Endocrinol. 1990, 79, 23–30. [Google Scholar] [CrossRef]
- Rideout, R.M.; Rose, G.A.; Burton, M.P.M. Skipped Spawning in Female Iteroparous Fishes. Fish Fish. 2005, 6, 50–72. [Google Scholar] [CrossRef]
- Rideout, R.M.; Tomkiewicz, J. Skipped Spawning in Fishes: More Common than You Might Think. Chang. Publ. Wiley 2011, 3, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Burton, M.P.M.; Penney, R.M.; Biddiscombe, S. Time Course of Gametogenesis in Northwest Atlantic Cod (Gadus morhua). Can. J. Fish. Aquat. Sci. 1997, 54, 122–131. [Google Scholar] [CrossRef]
- Jonsson, N.; Jonsson, B.; Hansen, L.P. Energetic Cost of Spawning in Male and Female Atlantic Salmon (Salmo salar L.). J. Fish Biol. 1991, 39, 739–744. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty Acid Requirements in Ontogeny of Marine and Freshwater Fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Parma, L.; Bonaldo, A.; Pirini, M.; Viroli, C.; Parmeggiani, A.; Bonvini, E.; Gatta, P.P. Fatty Acid Composition of Eggs and Its Relationships to Egg and Larval Viability from Domesticated Common Sole (Solea Solea) Breeders. Reprod. Domest. Anim. 2015, 50, 186–194. [Google Scholar] [CrossRef]
- Vasconi, M.; Aidos, L.; Di Giancamillo, A.; Bellagamba, F.; Domeneghini, C.; Moretti, V.M. Effect of Temperature on Fatty Acid Composition and Development of Unfed Siberian Sturgeon (A. Baerii) Larvae. J. Appl. Ichthyol. 2019, 35, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.R.; Baptista, M.; Rosa, I.C.; Dionísio, G.; Gomes-Pereira, J.; Paula, J.R.; Figueiredo, C.; Bandarra, N.; Calado, R.; Rosa, R. “Gone with the Wind”: Fatty Acid Biomarkers and Chemotaxonomy of Stranded Pleustonic Hydrozoans (Velella Velella and Physalia Physalis). Biochem. Syst. Ecol. 2016, 66, 297–306. [Google Scholar] [CrossRef]
- Fuiman, L.A.; Perez, K.O. Metabolic Programming Mediated by an Essential Fatty Acid Alters Body Composition and Survival Skills of a Marine Fish. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151414. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Fuiman, L.A. Maternal Diet Affects Utilization of Endogenous Lipids by Red Drum Sciaenops Ocellatus Embryos and Early Larvae. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110639. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.S.; Fernández-Palacios, H.; Tacon, A.G.J. Effect of Broodstock Nutrition on Reproductive Performance of Fish. Aquaculture 2001, 197, 25–42. [Google Scholar] [CrossRef]
- Pimentel, M.S.; Faleiro, F.; Diniz, M.; Machado, J.; Pousão-Ferreira, P.; Peck, M.A.; Pörtner, H.O.; Rosa, R. Oxidative Stress and Digestive Enzyme Activity of Flatfish Larvae in a Changing Ocean. PLoS ONE 2015, 10, e0134082. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioural Category | Behaviour |
|---|---|
| Aggression (frequency) | Charge-Quick movement towards a second fish (conspecific), while the conspecific avoids the first fish. Chase-Individual swimming after conspecific. |
| Hide (frequency) | Fish attempting to hide, either in shelter or in corner of the tank. |
| Swim (time, s) | Active swimming-Movement against current using pectoral fins and caudal body movement/Movement up and down the side of the tank using active swimming technique. Beat-and-glide-Intermittent swimming and drifting. Surface touching–fish swimming and touching the surface of the water. |
| Rest (time, s) | Motionless horizontally on bottom of aquarium. |
| Conditional Model–Time to Ignore | Estimate | z Value | Pr (<|z|) |
|---|---|---|---|
| CT + LR | −0.871 | −1.590 | 0.112 |
| HT + HR | −1.422 | −2.507 | 0.012 |
| HT + LR | 0.010 | 0.012 | 0.991 |
| Day | −0.370 | −2.068 | 0.039 |
| CT + LR × Day | 0.356 | 1.433 | 0.152 |
| HT + HR × Day | 0.469 | 1.813 | 0.070 |
| HT + LR × Day | 0.053 | 0.142 | 0.887 |
| Zero-inflation model | |||
| (Intercept) | −0.914 | −4.764 | <0.001 |
| Conditional model-Locomotion | Estimate | z value | Pr (<|z|) |
| CT + LR | 7.842 | 0.930 | 0.353 |
| HT + HR | 0.592 | 0.073 | 0.942 |
| HT + LR | −19.633 | −1.673 | 0.094 |
| Zero-inflation model | |||
| (Intercept) | −1.938 | −7.468 | <0.001 |
| Conditional model–Rest | |||
| CT + LR | −0.826 | −0.558 | 0.577 |
| HT + HR | 0.831 | 0.574 | 0.566 |
| HT + LR | −1.855 | −0.865 | 0.387 |
| Conditional model–Hide | Estimate | z value | Pr (<|z|) |
| CT + LR | −1.843 | −2.251 | 0.024 |
| HT + HR | −1.540 | −1.925 | 0.054 |
| HT + LR | 3.517 | 3.000 | 0.003 |
| Conditional model-Aggression | |||
| CT + LR | 1.464 | 1.217 | 0.224 |
| HT + HR | 1.265 | 1.052 | 0.293 |
| HT + LR | −1.795 | −1.070 | 0.284 |
| Conditional Model | Estimate | t Value | Pr (>|t|) |
|---|---|---|---|
| CT + LR | −65.68 | −0.449 | 0.656 |
| HT + HR | −203.53 | −1.392 | 0.173 |
| HT + LR | 147.99 | 0.759 | 0.453 |
| Conditional Model–SL | Estimate | t Value | Pr (>|t|) |
|---|---|---|---|
| HT + LR | −0.120 | −0.915 | 0.366 |
| HT + HR | −0.003 | −0.027 | 0.979 |
| HT + HR | 0.053 | 0.285 | 0.777 |
| Conditional model-W | |||
| HT + LR | −0.140 | −1.662 | 0.105 |
| HT + HR | 0.028 | 0.348 | 0.730 |
| HT + HR | −0.006 | −0.047 | 0.963 |
| Fatty Acids | Control Temperature | High Temperature |
|---|---|---|
| Saturated (SFA) | ||
| 14:0 | 0.59 ± 0.26 | 0.70 ± 0.07 |
| 16:0 | 11.84 ± 0.71 | 12.68 ± 0.55 |
| 18:0 | 7.32 ± 0.55 | 7.36 ± 0.29 |
| Other SFA | 4.41 ± 0.25 | 4.35 ± 0.30 |
| ∑ SFA | 24.15 ± 0.98 | 25.08 ± 0.64 |
| Monounsaturated (MUFA) | ||
| 16:1 n-9/n-7 | 5.25 ± 0.71 | 5.20 ± 0.48 |
| 18:1 n-9/n-7 | 19.61 ± 5.28 | 19.88 ± 3.21 |
| Other MUFA | 0.61 ± 0.09 | 0.64 ± 0.04 |
| ∑ MUFA | 25.47 ± 4.84 | 25.71 ± 3.47 |
| Polyunsaturated (PUFA) | ||
| 18:2 n-6 | 7.61 ± 0.67 | 6.97 ± 0.64 |
| 18:3 n-3 | 8.87 ± 3.69 | 9.00 ± 0.92 |
| 20:4 n-6 | 2.18 ± 0.13 | 2.24 ± 0.20 |
| 20:5 n-3 | 4.93 ± 0.71 | 5.68 ± 0.42 |
| 22:5 n-3 | 1.30 ± 0.39 | 2.66 ± 3.64 |
| 22:6 n-3 | 5.86 ± 2.62 | 7.29 ± 3.60 |
| Other PUFA | 6.24 ± 0.46 | 7.37 ± 4.67 |
| ∑ PUFA | 36.99 ± 4.09 | 41.21 ± 5.50 |
| ∑ n-3 | 24.42 ± 3.92 | 29.37 ± 6.05 |
| ∑ n-6 | 11.83 ± 1.01 | 11.38 ± 0.63 |
| n-3/n-6 | 2.08 ± 0.39 | 2.61 ± 0.66 |
| Total | 86.60 ± 7.57 | 91.99 ± 5.50 |
| 100-TFA | 13.40 ± 7.57 | 8.01 ± 5.50 |
| Conditional Model | Estimate | t Value | Pr (>|t|) |
|---|---|---|---|
| LPO | 0.006 | 2.239 | 0.033 |
| TAC | 0.007 | 0.155 | 0.878 |
| HSP70 | −0.094 | −0.953 | 0.348 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, A.F.; Murdoch, R.; Martins-Cardoso, S.; Madeira, C.; Costa, P.M.; Félix, A.S.; Oliveira, R.F.; Bandarra, N.M.; Vinagre, C.; Lopes, A.R.; et al. Differential Effects of Food Restriction and Warming in the Two-Spotted Goby: Impaired Reproductive Performance and Stressed Offspring. Fishes 2022, 7, 194. https://doi.org/10.3390/fishes7040194
Lopes AF, Murdoch R, Martins-Cardoso S, Madeira C, Costa PM, Félix AS, Oliveira RF, Bandarra NM, Vinagre C, Lopes AR, et al. Differential Effects of Food Restriction and Warming in the Two-Spotted Goby: Impaired Reproductive Performance and Stressed Offspring. Fishes. 2022; 7(4):194. https://doi.org/10.3390/fishes7040194
Chicago/Turabian StyleLopes, Ana F., Robyn Murdoch, Sara Martins-Cardoso, Carolina Madeira, Pedro M. Costa, Ana S. Félix, Rui F. Oliveira, Narcisa M. Bandarra, Catarina Vinagre, Ana R. Lopes, and et al. 2022. "Differential Effects of Food Restriction and Warming in the Two-Spotted Goby: Impaired Reproductive Performance and Stressed Offspring" Fishes 7, no. 4: 194. https://doi.org/10.3390/fishes7040194
APA StyleLopes, A. F., Murdoch, R., Martins-Cardoso, S., Madeira, C., Costa, P. M., Félix, A. S., Oliveira, R. F., Bandarra, N. M., Vinagre, C., Lopes, A. R., Gonçalves, E. J., & Faria, A. M. (2022). Differential Effects of Food Restriction and Warming in the Two-Spotted Goby: Impaired Reproductive Performance and Stressed Offspring. Fishes, 7(4), 194. https://doi.org/10.3390/fishes7040194