Abstract
The common carp (Cyprinus carpio) is farmed globally; as a subspecies, the Yellow River carp (C. c. haematopterus) is widely farmed in northern China. Since the Yellow River carp grows slowly in winter, it would benefit breeders to implement a breeding program targeting growth in winter. In May 2017, we established a Yellow River carp family and measured the body length, height, thickness, and weight of individuals before (October 2017) and after winter (February 2018). Based on the genetic linkage map established in our laboratory, the quantitative trait locus (QTL) mapping of these four traits led to the detection of 29 growth-related QTLs, of which 9, 7, 3, and 10 QTLs were associated with body length, height, thickness, and body weight, respectively. One consensus QTL contained seven QTLs associated with four traits. Through the two major-effect QTLs (qBL-p-1 and qBL-p-2), we screened three candidate genes (fnd3b, ghsr, and pld1), which were associated with biological activities, such as fat formation, growth, and membrane transport. These results are helpful in exploring the growth regulation mechanism and providing a reference for genetic breeding with regard to the overwintering growth of Yellow River carp.
1. Introduction
The common carp (Cyprinus carpio) has a farming history of more than 8000 years in some parts of China [1]. Nowadays, the common carp is farmed globally with a production higher than 4.41 million tons in 2019 [2]. The Yellow River carp is a subspecies of the common carp that has a large yield in northern China. The rapid development of aquaculture has led researchers to focus their work on common carp genetics and breeding [3,4,5,6].
Among the methods used in breeding research, such as selective breeding and cross-breeding, quantitative trait locus (QTL) mapping is important, as it helps screen trait-related markers and assists breeding; studies on growth, feed conversion rate, disease resistance, sex, and muscle quality have been carried out using QTL [7]. Completion of the mapping of the carp genome resulted in the determination of the QTL location with increased accuracy [8,9]. Growth is one of the most important breeding traits. In research on growth-related QTL mapping, Laghari et al. [10] analyzed three traits of common carp F1 families 300 days post hatching, detecting four major-effect QTLs that explained more than 20% of the phenotypic variance. Peng et al. [11] mapped the body weight (BW), body length (BL), and carcass weight of Yellow River carp (C. c. haematopterus) 18 months after hatching (MAH) and obtained a total of 22 QTLs and several growth-related candidate genes. Feng et al. [12] identified 21 QTLs related to BW, total length, BL, body height (BH), or head length in Yangtze River common carp at 9 MAH using microsatellite and single nucleotide polymorphism (SNP) markers. By injecting passive integrated transponder (PIT) markers into common carp at 3 MAH and measuring their growth traits at 3 and 8 MAH, Su et al. [4] not only located 18 growth-related QTLs but also found a single shared QTL that partially controlled the BL, body depth, and body width. Wang et al. [13] detected a total of 14 major-effect QTLs and 17 candidate genes associated with BL, body thickness (BT), BH, or BW in three growth stages. However, there are few reports on cold-tolerant growth.
While studying the cold tolerance of common carp, Sun et al. [14] tagged 100 common carp before overwintering and identified living and dead individuals after overwintering. In that study, a genetic linkage map was constructed, and four putative markers associated with cold tolerance were identified, but only one marker could be mapped on a linkage group (LG). Liang et al. [15] detected a large number of cold-response genes in different tissues at different temperatures by transcriptomic analysis. Sun et al. [16] researched the effect of high and low temperatures on the common carp liver transcriptome. The results showed that the change in temperature had a great influence on the energy metabolism and metabolic process of cell membranes. Ge et al. [17] revealed that FoxO-related signal pathways play an important role in the formation and acclimation of the cold tolerance of Songpu mirror carp. Long et al. [18] analyzed the low-temperature-acclimation transcriptome of three common carp strains and found that low-temperature acclimation was consistent with the upregulation of genes related to cholesterol biosynthesis, and the content of total cholesterol in the brain increased significantly under cold stress.
Although many studies have been conducted on growth-related QTL mapping and cold tolerance, studies on growth-related QTL during the overwintering period have not yet been reported. However, during winter, the growth of Yellow River carp is almost stagnant. Prolonging the growth period of the Yellow River carp into the winter would play an important role in promoting the development of its farming. It is possible to screen QTL and candidate genes related to overwintering growth by using growth-related QTL mapping of overwintering. The QTL and candidate gene may also be related to cold tolerance. The aim of this study is to provide a reference for the growth and cold-tolerant breeding of Yellow River carp. To this end, we used high-density genetic maps of Yellow River carp [13] constructed by our group to locate growth-related QTL and to screen major candidate genes according to the BL, BH, BT, and BW of Yellow River carp during overwintering.
2. Materials and Methods
2.1. Samples and Phenotypic Collection
In 2015, we established nine carp families. At 11 months after hatching (MAH), the growth was analyzed; one family with fast growth and one family with slow growth were screened. In May 2017, a female from a fast-growing and a male from a slow-growing Yellow River carp family established in 2015 were selected as parents to establish a family in the Aquaculture Base of Henan Normal University. In October 2017 (5 MAH), 300 carps were randomly selected to be anesthetized with MS-222 (tricaine methanesulfonate) and tagged with passive integrated transponder (PIT). The body length (BL), body height (BH), body thickness (BT), and body weight (BW) of the carps were measured. The carps were placed in a solution of 0.5% potassium permanganate for 15–20 min to remove potential pathogens and put into a cement pond to continue the culture. The average value, standard deviation of each trait, and Pearson correlation coefficient between traits were analyzed using SPSS 24.0 (IBM Corp, Armonk, NY, USA). The tagged carps were measured again in February 2018 (9 MAH), and approximately 0.3-g caudal fins were cut at the age of 17 MAH for the following experiments (N = 209). The genomic DNA of the carp was extracted using the phenol-chloroform DNA extraction method and checked using a NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Scientific, Waltham, MA, USA). The range and average of water temperature during overwintering was 1–20 °C and 9.53 ± 5.80 °C. For apparent errors in the growth data of some individuals before and after overwintering, we only used the information of 201 carps for this experiment.
2.2. QTL Mapping
The DNA samples were sent to Shanghai Ouyi Biotechnology Co., Ltd. (Shanghai, China) for sequencing. The libraries of the parents and progeny were prepared following Qi et al. [19]. Read processing, SNP calling, and filtering were conducted following Qi et al. [19]. Chi-square tests were used to determine the markers’ deviation from expected Mendelian segregation ratios. Population type was set as “backcross” (BC) for the maternal and paternal data sets and as “F2 intercross” for the HH data set [19]. SNPs that were heterozygous in both parents with progeny segregating as A:H:B (1:2:1) constituted the HH dataset. SNPs with genotypic score A or B in parent 1 and H in parent 2 and segregating as A:H or B:H (1:1) were combined, and B scores were converted to A scores to form the paternal (AH) dataset. Similarly, H:A and H:B markers were combined, and B scores were converted to A scores to yield the maternal (HA) dataset following Qi et al. [19].
The HA, AH, and HH genetic maps were constructed using a combination of MSTMap [20] and MAPMAKER (modified from Lander et al. [21] as described by Qi et al. [22]. The population type was set as “backcross” for the maternal and paternal datasets and as “F2 intercross” for the HH dataset. Co-segregating SNPs were added to the framework map to the same location as their representative marker using an in-house Python script. Linkage maps were drawn with MapChart [23]. Based on the genetic linkage maps of Yellow River carp established by our laboratory [13], the composite interval mapping model in WinQTLCart 2.5 [24] was used to map the QTL of net change in BL, BH, BT, and BW in Yellow River carp from 5 to 9 MAH. The population type for the QTL analysis was set as “SF2” for the HH maps and as “B2” for the maternal and paternal maps. A walk speed of 0.5 cM was used for QTL identification. The logarithm of the odds (LOD) threshold for significant QTL (p ≤ 0.05) was determined by 1000 permutations. After the QTLs were located, all QTLs located within 1 Mb of each other on the physical map were identified as being consensus QTL.
QTL naming rules: q + the abbreviation of growth traits + map type (m: maternal map, p: paternal map, h: HH map) + QTL number. For example, qBL-m-1 represents the first QTL associated with body length in the maternal map; qBW-p-1 represents the first QTL associated with body weight in the paternal map; qBH-h-1 represents the first QTL associated with body height in the HH map.
2.3. Candidate Genes
Growth-related candidate genes were filtered from these QTLs with LOD > 10. Briefly, we screened the ±50 kb genome regions surrounding the significant SNPs based on the Yellow River carp reference genome [25] (BIGD Genome Warehouse, https://bigd.big.ac.cn/gwh/Assembly/497/show, accessed on 27 October 2021) and annotated the candidate genes by conducting BLAST analysis against the Swiss-Prot database.
3. Results
3.1. Growth Traits in Yellow River Carp Aged 5–9 MAH
Table S1 shows the measurements of four growth traits in Yellow River carp at the ages of 5 MAH and 9 MAH. Based on these data, we calculated the growth trait changes in Yellow River carp at 5 to 9 MAH. The net growth of BL, BH, BT, and BW of Yellow River carp at 5–9 MAH was 0.8 ± 0.7 cm, 0.0 ± 0.2 cm, 0.4 ± 0.4 cm, and 8.2 ± 5.9 g, respectively. There were great differences in growth among individuals; the highest and lowest increases in BW were 32.8 g and −0.8 g, respectively. The correlation of the four growth traits between 5 and 9 MAH, shown in Table 1, suggests that there was a moderately positive correlation between BW and BL (r = 0.361). There was a weak positive correlation between BL and BH (r = 0.187), while the correlation between other traits was not significant (p > 0.05). The results suggest that the increase in the BW of Yellow River carp at the age of 5–9 MAH was mainly associated with the increase in BL.

Table 1.
Pearson correlations between the growth traits of Yellow River carp at 5–9 months after hatching.
3.2. BL-Related QTL of Yellow River Carp at 5–9 MAH
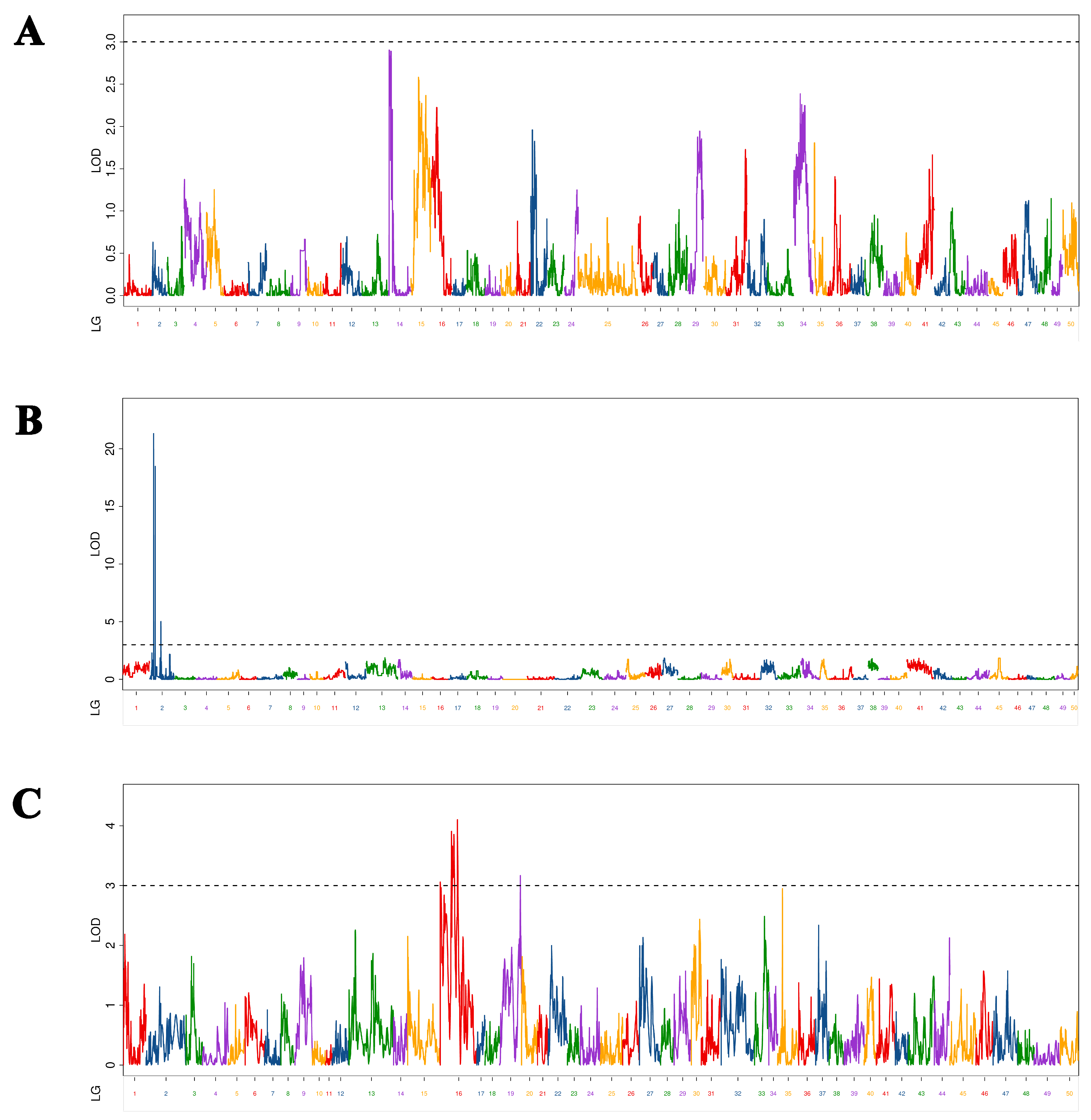
We identified nine BL-associated QTLs of Yellow River carp at 5–9 MAH (Figure 1). As shown in Table 2, the LOD values range from 3.06 to 21.32, with a PVE (phenotypic variation explained) of 2.52% to 29.13% and additive effect of −3.49 to 0.59. Nine QTLs were located on three chromosomes (three QTLs on chromosome 4, five QTLs on chromosome 12, and one QTL on chromosome 13). There were two QTLs with LOD values greater than 10%: qBL-p-1 (LOD = 21.32, PVE = 29.13%) and qBL-p-2 (LOD = 18.47, PVE = 24.34%), located on chromosome 4. These QTLs were located at 20,559,100 bp and 12,719,902 bp on chromosome 4, respectively. Of the nine markers, only one marker (qBL-h-5) was located in an exonic region, while the others were located at intergenic regions.
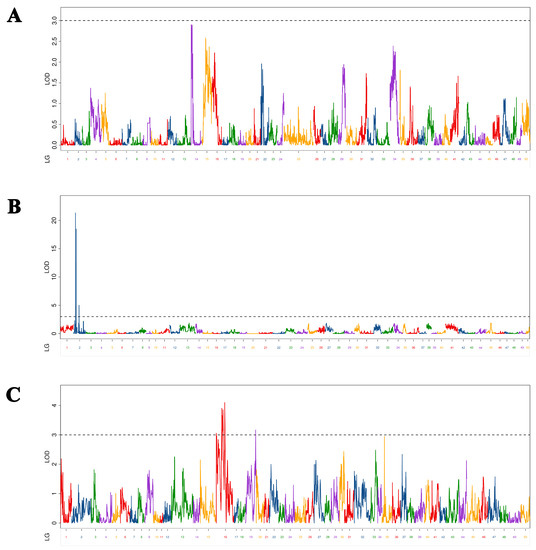
Figure 1.
Distribution of body length-related QTLs on 50 linkage groups of Yellow River carp in the maternal (A), paternal (B), and HH maps (C). Different color indicates different linkage group.
Table 2.
Information on body length-related QTLs in Yellow River carp at 5–9 months after hatching.
3.3. BH-Related QTLs in Yellow River Carp at 5–9 MAH
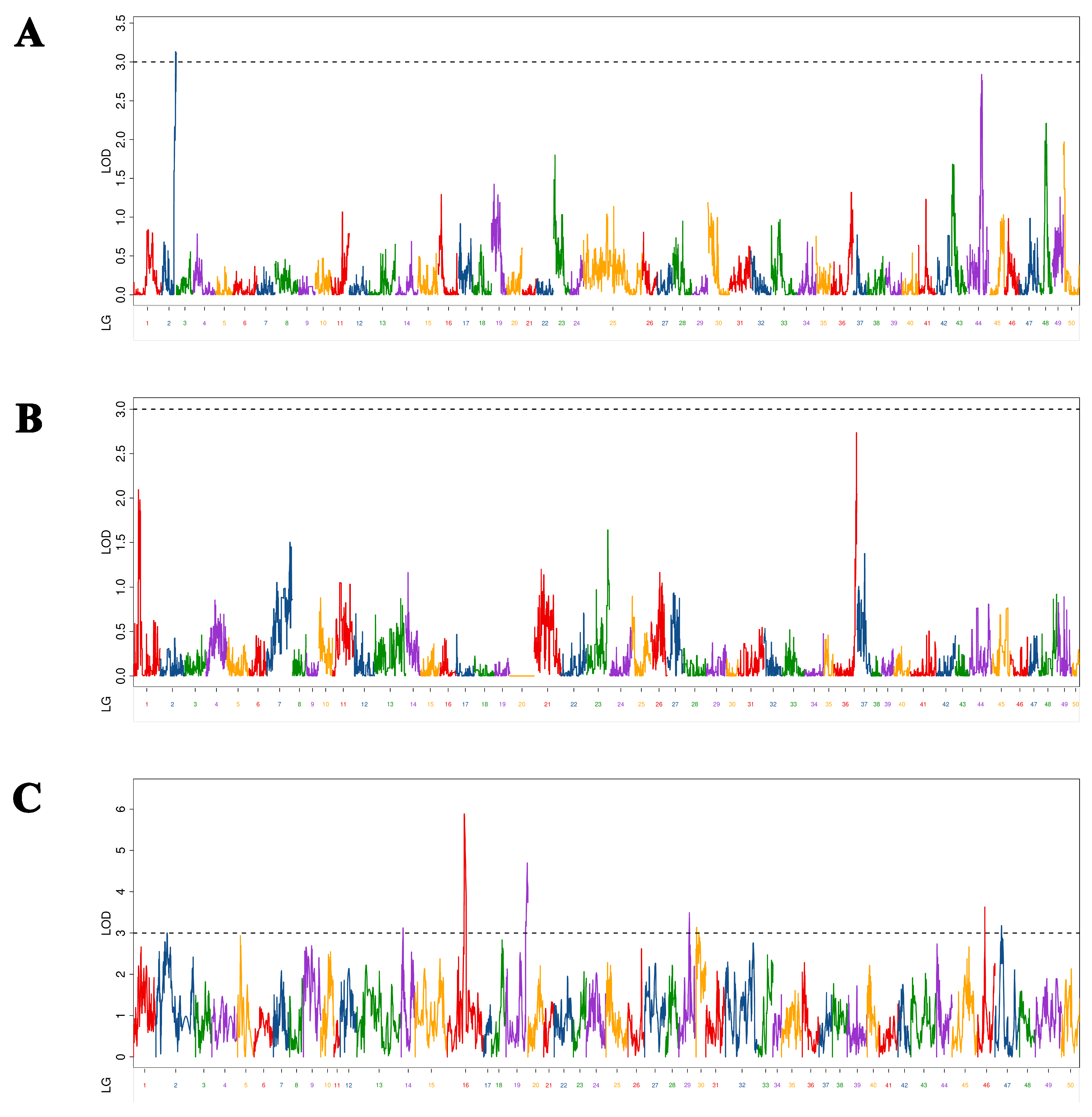
In this study, seven BH-associated QTLs were identified in Yellow River carp at 5–9 MAH (Figure 2). As shown in Table 3, LOD values ranged from 3.12 to 5.89, with a PVE of 0.38% to 11.20% and additive effect of −0.33 to 0.26. Seven QTLs were located on seven chromosomes (chromosomes 2, 12, 13, 18, 24, 29, and 47, respectively). Among them, qBH-h-2 had the maximum LOD value (5.89), and PVE (11.20%) was mapped at 25,680,671 bp on chromosome 12 in the physical map. The QTL of qBH-h-1 with the minimum LOD value (3.12) and PVE (0.38%) was mapped at 13,961,785 bp on chromosome 18 in the physical map. Of these seven markers, two markers (qBH-h-2 and qBH-h-5) were located in exonic regions; qBH-h-6 was located in an intronic region; the other four markers were located in intergenic regions.
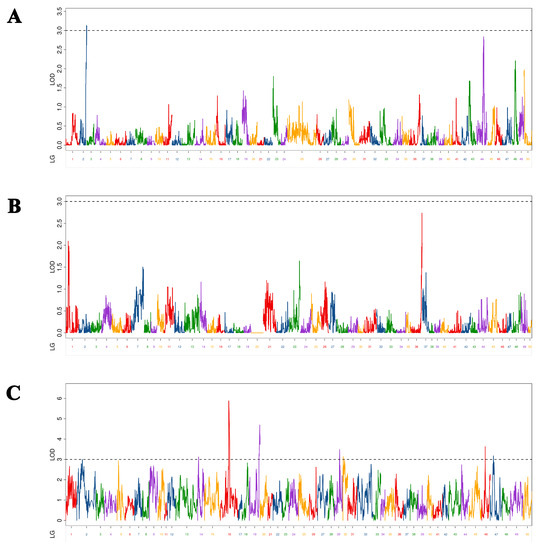
Figure 2.
Distribution of body height-related QTLs on 50 linkage groups of Yellow River carp in the maternal (A), paternal (B), and HH maps (C). Different color indicates different linkage group.
Table 3.
Information on body height-related QTLs in Yellow River carp at 5–9 months after hatching.
3.4. BT-Related QTLs in Yellow River Carp at 5–9 MAH
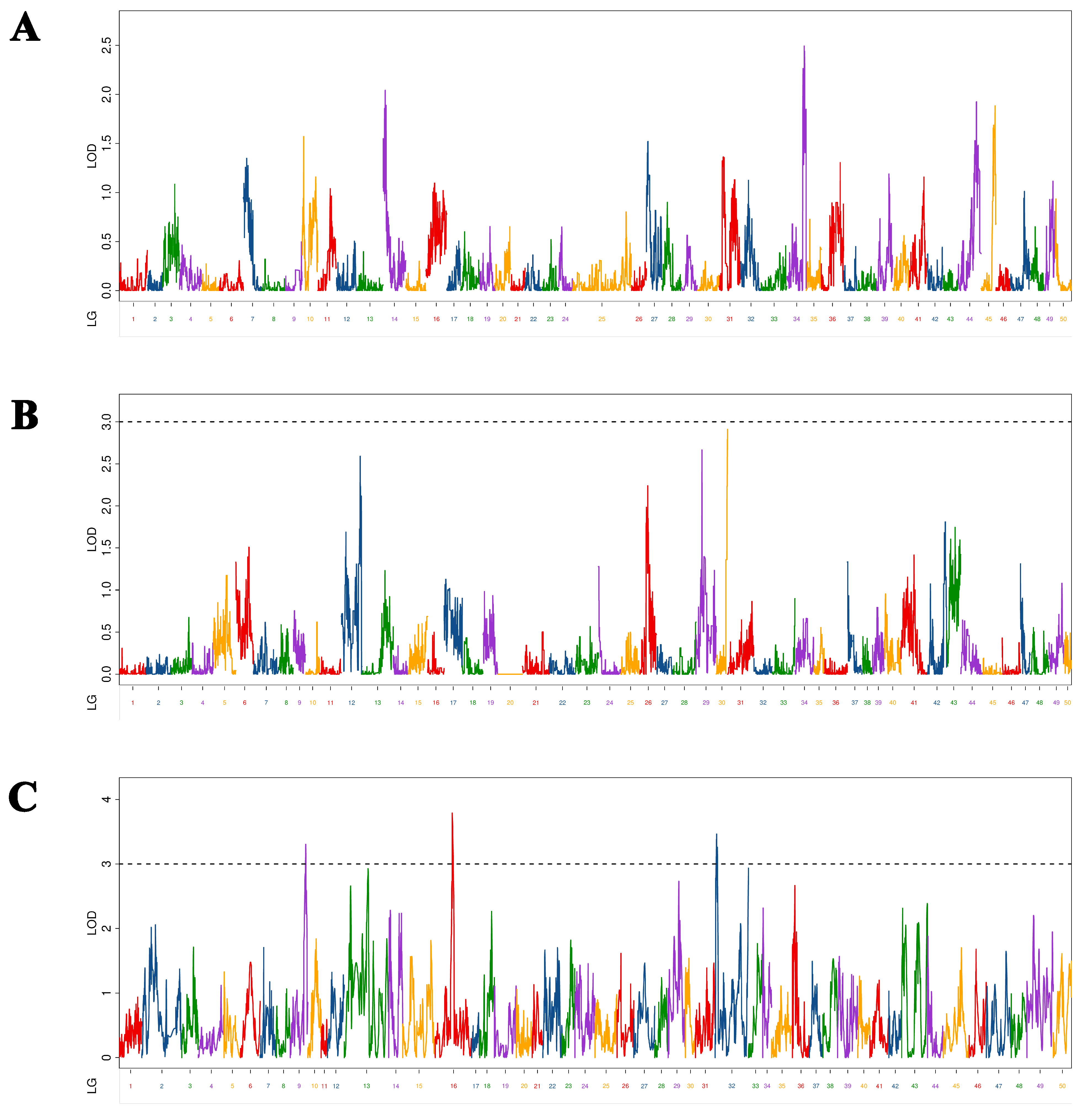
We identified three BT-associated QTLs in Yellow River carp at 5–9 MAH (Figure 3). As shown in Table 4, the LOD values range from 3.26 to 3.78, with a PVE of 1.88% to 7.26% and an additive effect of −0.19 to 0.18. Three QTLs were located on three chromosomes (chromosomes 4, 12, and 31). Among the QTLs, qBT-h-2 had the maximum LOD value (3.79), and PVE (7.26%) was mapped at 25,680,671 bp on chromosome 12 in the physical map. Meanwhile, qBT-h-1, qBT-h-2, and qBT-h-1 were located in intronic, exonic, and intergenic regions, respectively.
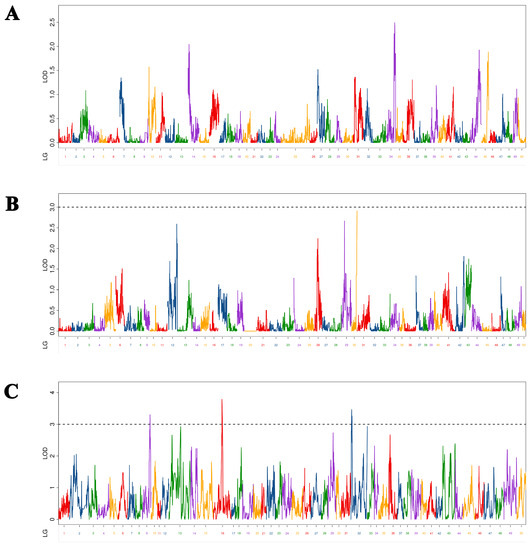
Figure 3.
Distribution of body thickness-related QTLs in 50 linkage groups of Yellow River carp in the maternal (A), paternal (B), and HH maps (C). Different color indicates different linkage group.
Table 4.
Information on body thickness-related QTLs in Yellow River carp at 5–9 months after hatching.
3.5. BW-Related QTLs in Yellow River Carp at 5–9 MAH
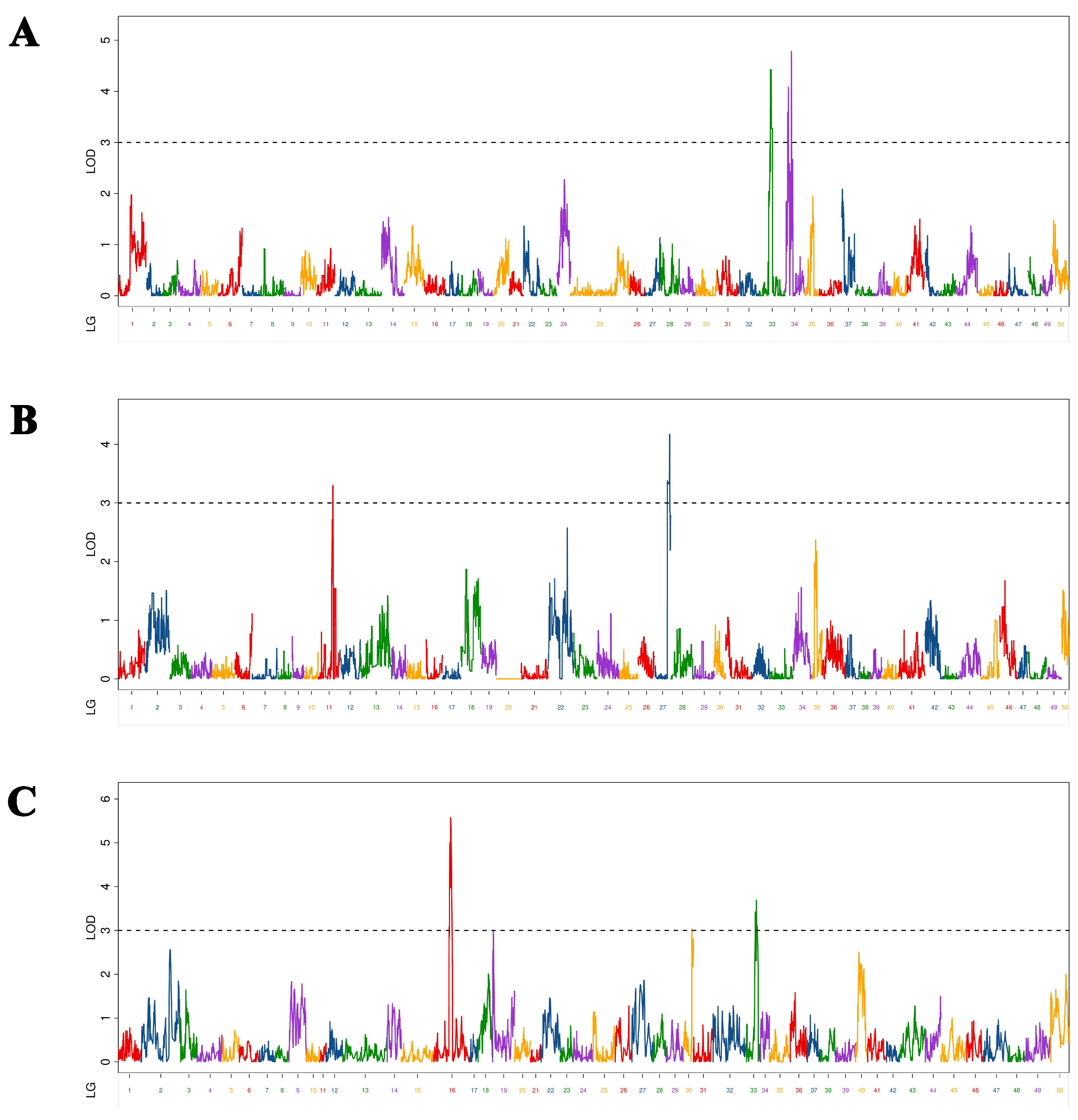
We identified 10 BW-associated QTLs in Yellow River carp at 5–9 MAH (Figure 4). As shown in Table 5, the LOD values range from 3.00 to 5.58, with a PVE of 2.07% to 8.85% and an additive effect of −52.80 to 52.62. These QTLs were located on three chromosomes (one QTL on chromosomes 13 and 33 and two QTLs on chromosomes 12, 29, 34, and 39). Among them, qBW-h-1 with the maximum LOD value (5.58) was mapped at 25,680,671 bp on chromosome 12 in the physical map. Of the ten markers, qBW-h-3 was located in exonic regions; there were four markers (qBW-p-1, qBW-h-1, qBW-h-2 and qBW-h-5) located in intronic regions; the other five markers were located in intergenic regions.
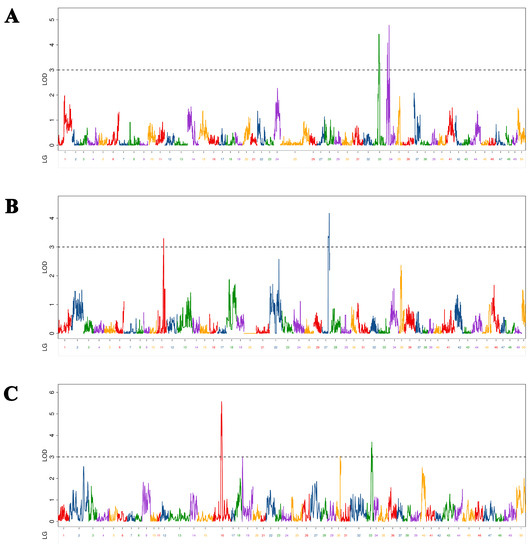
Figure 4.
Distribution of body weight-related QTLs on 50 linkage groups of Yellow River carp in the maternal (A), paternal (B), and HH maps (C). Different color indicates different linkage group.
Table 5.
Information on body weight-related QTLs in Yellow River carp at 5–9 months after hatching.
3.6. Consensus QTL at 5–9 MAH
We screened one consensus QTL (C-QTL) containing seven QTLs (qBL-h-1, qBL-h-2, qBL-h-5, qBH-h-2, qBT-h-2, qBW-p-1, and qBW-h-1). Among these, the nearest marker position of qBL-h-5, qBH-h-2, qBT-h-2, and qBW-h-1 was the same at 25,680,671 bp on chromosome 12. The marker positions of qBL-h-1, qBL-h-2, and qBW-p-1 were 25,507,539 bp, 25,495,788 bp, and 24,883,274 bp, respectively. As shown in Table 6, C-QTL may control four growth traits (BL, BH, BT, and BW) at the same time, thereby being pleiotropic.
Table 6.
Information on the consensus QTL in Yellow River carp at 5–9 months after hatching.
3.7. Candidate Genes
According to the standard of candidate gene screening, three candidate genes (Table 7) related to growth were screened at 5–9 MAH. These three candidate genes on chromosome 4 are closely related to qBL-p-2. Among them, Fibronectin type III domain-containing protein 3B (Fnd3b), also termed factor for adipocyte differentiation 104 (Fad104), may be a positive regulator of adipogenesis [26]. Growth hormone secretagogue receptor type 1 (Ghsr) is a receptor for ghrelin; coupled to G-alpha-11 proteins, it can stimulate growth hormone secretion. The gene also binds other growth hormone-releasing peptides (Ghrp) (e.g., Met-enkephalin and Ghrp-6) as well as non-peptide, low-molecular-weight secretagogues (e.g., L-692,429, MK-0677, and adenosine) [27,28]. Phospholipase D1 (Pld1) functions as phospholipase selective for phosphatidylcholine, which is implicated as a critical step in numerous cellular pathways, including signal transduction, membrane trafficking, and the regulation of mitosis [29,30].
Table 7.
Candidate genes related to major effect QTLs in Yellow River carp at 5–9 months after hatching.
4. Discussion
Our study on the growth-related QTLs of Yellow River carp during overwintering showed that the family grew slowly during the overwintering period. There were significant differences in the added value of BW among individuals, and the increase in BW resulted mainly from the increase in BL. At present, there are some reports about the growth-related QTL of fish at a certain growth stage. For example, Sauvage et al. [31] tagged brook charr (Salvelinus fontinalis) with PIT at the age of 5 months, collected the growth data of four different growth periods (May 2009, July 2009, August 2009, and November 2009), and mapped the growth rate-related QTLs in different growth periods. Zhang et al. [32] measured the growth rate of common carp at three time points (3, 4, and 5 MAH) and identified 15 genomic regions associated with growth rate. Wang et al. [13] carried out a QTL mapping study on four growth traits of Yellow River carp at the age of 13, 17, and 13–17 months. However, there is no research on the QTL location related to the growth of fish during overwintering. This may be due to the slow growth of carp during overwintering; some scholars think that the research is of little significance. Although carp grow slowly during overwintering, there are great differences among individuals. Thus, we can screen the major-effect QTL during overwintering by detecting the QTL related to growth traits. These QTL may also be related to the cold tolerance of carp.
At present, the research on the cold tolerance of fish is mainly focused on the transcriptomic analysis of the common carp [18,20], Nile tilapia (Oreochromis niloticus) [33], yellow drum (Nibea albiflora) [34], Onychostoma macrolepis [35], etc. However, there are few studies on cold-tolerance-related QTL. At present, there is only one report on the location of cold-tolerance-related QTL in common carp [14]. In this study, the dead and surviving fish were classified into two groups after overwintering. Four RAPD markers that may be associated with cold tolerance were screened, but only one was located in the linkage group. In this study, 300 fish were initially marked, but only 198 fish were used because the samples were collected at the age of 17 MAH for QTL mapping. There were still 207 fish at the age of 17 MAH, but the growth data of nine fish were obviously wrong during the overwintering period, so these growth data were excluded. In future research, we should develop devices that can automatically and accurately obtain the phenotypic traits of fish and so improve the accuracy of the results [36].
In this study, as different maps (maternal, paternal, and HH) are constructed with different markers, the QTL mapping results of the same trait on these maps are very different. Twenty-nine QTLs related to overwintering growth were located and distributed on 12 chromosomes. The characteristics of a large number and dispersion of QTL were similar to those mentioned by other studies on common carp growth-related QTL [10,11,12,37]. The largest number of growth-related QTLs in common carp was 165, distributed in 30 linkage groups [38], indicating that common carp growth-related traits were determined by multiple genes. The results of the present study show that these growth-related, complex traits are controlled by a large number of genes. It is generally assumed that the higher the LOD value, the higher the confidence, and the higher the PVE, the greater the role of QTL. In this study, there were only two BL-related QTLs that had LOD > 10 and PVE > 20%, which were values similar to those obtained by a study on the Yangtze River common carp growth-related QTL [12]. In that study, the authors found that one BW-related QTL had LOD > 10 and PVE > 20%. However, although several QTLs have been found in studies of Yellow River carp [11] and common carp [10] with PVE >20%, none had LOD >10%. Therefore, assumptions regarding whether there was a QTL that had a significant effect on different growth traits of common carp at different growth stages were quite different. However, these QTLs with significant effects are the ones we should focus on, as they can be used in molecular-assisted breeding, similar to the two QTLs that had significant effects and were screened in this study. The molecular markers and genes will help increase overwintering growth and prolong the growth period. In this study, C-QTL contained seven QTLs associated with four growth traits and had obvious pleiotropy. In previous studies, certain QTLs appeared pleiotropic in different periods [13] or different families [38]. The consensus QTLs may be related to housekeeping genes if their PVE is very low. In this study, the PVE of C-QTL was low, which may have little effect on overwintering growth. Although many markers were located in genes, the major QTLs were located in intergenic regions.
Among the three overwintering growth-related genes screened in this study, fnd3b is a protooncogene involved in the occurrence and development of cancer [39,40]. The protein Fnd3b expressed in this gene is a positive regulator of adipogenesis, related to adipocyte differentiation, and is an RNA-binding protein [41,42]. Other studies have shown that Fnd3b affects human craniofacial development [43] and is related to the domestication of chickens [44]. Ghsr, the receptor of growth hormone secretory hormone (Ghrelin) and growth hormone-releasing peptide, is a G protein coupling protein [45]. The Ghsr/Ghrelin system is an important pathway for the regulation of growth hormone secretion [46,47]. Ghsr has been found to play an important role in energy balance and metabolism as well as digestion and GH secretion in fish [48]. However, in the study of fish QTL, no QTL related to Ghsr was found. Pld1 is an enzyme widely distributed in bacteria, fungi, animals, and plants [49]. It plays a key role in signal transduction, membrane transport, and cytoskeletal regulation [50,51,52]. Lipid catabolism plays an important role in enhancing the cold tolerance of fish [53]. Zheng et al. [54] studied the QTL related to muscle fat content and abdominal fat in common carp and located 18 genome-wide QTLs with suggestive significance. This result was different from the genes found in this study, which might be due to the different traits examined in the two studies. These three genes could be used as important candidates to study the growth and metabolism of Yellow River carp during overwintering.
In conclusion, this is the first study on the growth-related QTL mapping in overwintering of Yellow River carp. In this study, 29 QTLs, including two major-effect QLTs and one C-QTL related to overwintering growth traits, were detected. Additionally, we screened three genes related to overwintering growth (fnd3b, ghsr, and pld1), which were related to fat formation, growth, membrane transport, etc. The results of this study provide information for the breeding of Yellow River carp to improve growth and cold tolerance during overwintering.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/fishes7040166/s1, Table S1: Body length, height, thickness, and weight of the family at the age of 5, 9, and 5–9 months after hatching (MAH).
Author Contributions
Conceptualization, L.W. and X.L.; methodology, L.W. and L.Z.; validation, L.W.; formal analysis, L.Z. and J.C.; investigation, L.W.; resources, L.W. and X.L.; writing—original draft preparation, L.W. and L.Z.; writing—review and editing, L.W. and L.Z.; visualization, Y.C., M.Z., M.Y. and H.J.; supervision, X.L. and Z.Q.; project administration, L.W.; funding acquisition, L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant 31602149) and major public welfare projects in Henan Province (201300311300).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Academic Committee of Henan Normal University (HNSD-SCXY-2116BS1029).
Data Availability Statement
GBS reads were submitted to NCBI-SRA (Acc. PRJNA788161).
Conflicts of Interest
The authors declare no conflict of interest. The authors declare that they have no known competing financial interest or personal relationship that could have appeared to influence the work reported in this paper.
References
- Nakajima, T.; Hudson, M.J.; Uchiyama, J.; Makibayashi, K.; Zhang, J. Common carp aquaculture in Neolithic China dates back 8000 years. Nat. Ecol. Evol. 2019, 3, 1415–1418. [Google Scholar] [CrossRef]
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2019; FAO: Roma, Italy, 2021. [Google Scholar]
- Dong, Z.; Nguyen, N.H.; Zhu, W. Genetic evaluation of a selective breeding program for common carp Cyprinus carpio conducted from 2004 to 2014. BMC Genet. 2015, 16, 94. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Li, H.; Du, F.; Zhang, C.; Li, X.; Jing, X.; Liu, L.; Li, Z.; Yang, X.; Xu, P.; et al. Combined QTL and genome scan analyses with the help of 2b-RAD identify Growth-Associated genetic markers in a new Fast-Growing carp strain. Front. Genet. 2018, 9, 592. [Google Scholar] [CrossRef]
- Tadmor-Levi, R.; Hulata, G.; David, L. Multiple interacting QTLs affect disease challenge survival in common carp (Cyprinus carpio). Heredity 2019, 123, 565–578. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, P.; Huang, X.; Liu, Q.; Lin, B.; Fu, Y.; Gu, Q.; Hu, F.; Luo, K.; Zhang, C.; et al. The establishment of an autotetraploid fish lineage produced by female allotetraploid hybrids × male homodiploid hybrids derived from Cyprinus carpio (♀) × Megalobrama amblycephala (♂). Aquaculture 2020, 515, 734583. [Google Scholar] [CrossRef]
- Chen, L.; Xu, J.; Sun, X.; Xu, P. Research advances and future perspectives of genomics and genetic improvement in allotetraploid common carp. Rev. Aquacult. 2022, 14, 957–978. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, X.; Wang, X.; Li, J.; Liu, G.; Kuang, Y.; Xu, J.; Zheng, X.; Ren, L.; Wang, G.; et al. Genome sequence and genetic diversity of the common carp, Cyprinus carpio. Nat. Genet. 2014, 46, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, Q.; Yang, M.-D.H.; Li, Q.; Cui, M.; Dong, Z.; Wang, H.; Yu, J.; Zhao, Y.; Yang, C.; et al. Parallel subgenome structure and divergent expression evolution of allo-tetraploid common carp and goldfish. Nat. Genet. 2021, 53, 1493–1503. [Google Scholar] [CrossRef]
- Laghari, M.Y.; Lashari, P.; Zhang, X.; Xu, P.; Narejo, N.T.; Xin, B.; Zhang, Y.; Sun, X. QTL mapping for economically important traits of common carp (Cyprinus carpio L.). J. Appl. Genet. 2015, 56, 65–75. [Google Scholar] [CrossRef]
- Peng, W.; Xu, J.; Zhang, Y.; Feng, J.; Dong, C.; Jiang, L.; Feng, J.; Chen, B.; Gong, Y.; Chen, L.; et al. An ultra-high density linkage map and QTL mapping for sex and growth-related traits of common carp (Cyprinus carpio). Sci. Rep. 2016, 6, 26693. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Yu, X.; Fu, B.; Wang, X.; Liu, H.; Pang, M.; Tong, J. A high-resolution genetic linkage map and QTL fine mapping for growth-related traits and sex in the Yangtze River common carp (Cyprinus carpio haematopterus). BMC Genom. 2018, 19, 230. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jia, S.; Zhang, Y.; Jiang, S.; Chen, Y.; Chen, J.; Yu, M.; Zhang, L.; Qiao, Z.; Li, X. Construction of three High-Density genetic linkage maps and dynamic QTL mapping of growth traits in yellow river carp (Cyprinus carpio haematopterus). Curr. Issues Mol. Biol. 2021, 43, 160. [Google Scholar] [CrossRef]
- Sun, X.; Liang, L. A genetic linkage map of common carp (Cyprinus carpio L.) and mapping of a locus associated with cold tolerance. Aquaculture 2004, 238, 165–172. [Google Scholar] [CrossRef]
- Liang, L.; Chang, Y.; He, X.; Tang, R. Transcriptome Analysis to Identify Cold-Responsive Genes in Amur Carp (Cyprinus carpio haematopterus). PLoS ONE 2015, 10, e130526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhao, L.; Wu, H.; Lian, W.; Cui, C.; Du, Z.; Luo, W.; Li, M.; Yang, S. Analysis of miRNA-seq in the liver of common carp (Cyprinus carpio L.) in response to different environmental temperatures. Funct. Integr. Genom. 2019, 19, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.; Long, Y.; Shi, L.; Ren, J.; Yan, J.; Li, C.; Li, Q.; Cui, Z. Transcriptomic profiling revealed key signaling pathways for cold tolerance and acclimation of two carp species. BMC Genom. 2020, 21, 539. [Google Scholar] [CrossRef]
- Long, Y.; Li, X.; Li, F.; Ge, G.; Liu, R.; Song, G.; Li, Q.; Qiao, Z.; Cui, Z. Transcriptional Programs Underlying Cold Acclimation of Common Carp (Cyprinus carpio L.). Front. Genet. 2020, 11, 556418. [Google Scholar] [CrossRef]
- Qi, P.; Eudy, D.; Schnable, J.C.; Schmutz, J.; Raymer, P.L.; Devos, K.M. High Density Genetic Maps of Seashore Paspalum Using Genotyping-By-Sequencing and Their Relationship to The Sorghum Bicolor Genome. Sci. Rep. 2019, 9, 12183. [Google Scholar] [CrossRef]
- Wu, Y.; Bhat, P.R.; Close, T.J.; Lonardi, S. Efficient and accurate construction of genetic linkage maps from the minimum spanning tree of a graph. PLoS Genet. 2008, 4, e1000212. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Qi, P.; Gimode, D.; Saha, D.; Schröder, S.; Chakraborty, D.; Wang, X.; Dida, M.M.; Malmberg, R.L.; Devos, K.M. UGbS-Flex, a novel bioinformatics pipeline for imputation-free SNP discovery in polyploids without a reference genome: Finger millet as a case study. BMC Plant Biol. 2018, 18, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. HERED 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.C.J.B. Windows QTL Cartographer 2.5; North Carolina State University: Raleigh, NC, USA, 2012. [Google Scholar]
- Xu, P.; Xu, J.; Liu, G.; Chen, L.; Zhou, Z.; Peng, W.; Jiang, Y.; Zhao, Z.; Jia, Z.; Sun, Y.; et al. The allotetraploid origin and asymmetrical genome evolution of the common carp Cyprinus carpio. Nat. Commun. 2019, 10, 4625. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, K.; Kondo, C.; Johmura, Y.; Nishizuka, M.; Imagawa, M. The novel genefad 104, containing a fibronectin type III domain, has a significant role in adipogenesis. FEBS Lett. 2004, 577, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Date, Y.; Kojima, M.; Matsuo, H.; Hosoda, H.; Kangawa, K.; Nakazato, M. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar]
- Smith, R.G.; Leonard, R.; Bailey, A.R.T.; Palyha, O.; Feighner, S.; Tan, C.; Mckee, K.K.; Pong, S.; Griffin, P.; Howard, A. Growth hormone secretagogue receptor family members and ligands. Endocrine 2001, 14, 9–14. [Google Scholar] [CrossRef]
- Hammond, S.M.; Altshuller, Y.M.; Sung, T.C.; Rudge, S.A.; Rose, K.; Engebrecht, J.; Morris, A.J.; Frohman, M.A. Human ADP-ribosylation factor-activated phosphatidylcholine-specific phospholipase D defines a new and highly conserved gene family. J. Biol. Chem. 1995, 270, 29640–29643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, I.; Arnold, R.S.; Lambeth, J.D. Cloning and initial characterization of a human phospholipase D2 (hPLD2). ADP-ribosylation factor regulates hPLD2. J. Biol. Chem. 1998, 273, 12846–12852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvage, C.; Vagner, M.; Derome, N.; Audet, C.; Bernatchez, L. Coding gene SNP mapping reveals QTL linked to growth and stress response in brook charr (Salvelinus fontinalis). G3 2012, 2, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, C.; Lu, C.; Cao, D. Elite allele mining for growth rate traits in common carp (Cyprinus carpio) by association analysis. Aquac. Res. 2021, 52, 1192–1200. [Google Scholar] [CrossRef]
- Zhou, T.; Gui, L.; Liu, M.; Li, W.; Hu, P.; Duarte, D.F.C.; Niu, H.; Chen, L. Transcriptomic responses to low temperature stress in the Nile tilapia, Oreochromis niloticus. Fish Shellfish Immun. 2019, 84, 1145–1156. [Google Scholar] [CrossRef]
- Jiao, S.; Nie, M.; Song, H.; Xu, D.; You, F. Physiological responses to cold and starvation stresses in the liver of yellow drum (Nibea albiflora) revealed by LC-MS metabolomics. Sci. Total Environ. 2020, 715, 136940. [Google Scholar] [CrossRef]
- Gou, N.; Ji, H.; Wu, W.; Zhong, M.; Zhang, B. Transcriptional response to cold and fasting acclimation in Onychostoma macrolepis during the overwintering stage. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100901. [Google Scholar] [CrossRef]
- Fu, G.; Yuna, Y. Phenotyping and phenomics in aquaculture breeding. Aquac. Fish. 2022, 7, 140–146. [Google Scholar] [CrossRef]
- Wang, X.; Fu, B.; Yu, X.; Qu, C.; Zhang, Q.; Tong, J. Fine mapping of growth-related quantitative trait loci in Yellow River carp (Cyprinus carpio haematoperus). Aquaculture 2018, 484, 277–285. [Google Scholar] [CrossRef]
- Lv, W.; Zheng, X.; Kuang, Y.; Cao, D.; Yan, Y.; Sun, X. QTL variations for growth-related traits in eight distinct families of common carp (Cyprinus carpio). BMC Genet. 2016, 17, 65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, S.; Hu, T.; Liu, S.; He, Y.; Sun, S. Up-regulated microRNA-143 transcribed by nuclear factor kappa B enhances hepatocarcinoma metastasis by repressing fibronectin expression. Hepatology 2009, 50, 490–499. [Google Scholar] [CrossRef]
- Cai, C.; Rajaram, M.; Zhou, X.; Liu, Q.; Marchica, J.; Li, J.; Powers, R.S. Activation of multiple cancer pathways and tumor maintenance function of the 3q amplified oncogene FNDC3B. Cell Cycle 2012, 11, 1773–1781. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Li, H.; Li, X.; Yu, S.; Xiang, H.; Peng, J.; Jiang, S. MicroRNA-215 impairs adipocyte differentiation and co-represses FNDC3B and CTNNBIP1. Int. J. Biochem. Cell Biol. 2016, 79, 104–112. [Google Scholar] [CrossRef]
- Tominaga, K.; Johmura, Y.; Nishizuka, M.; Imagawa, M. Fad24, a mammalian homolog of Noc3p, is a positive regulator in adipocyte differentiation. J. Cell Sci. 2004, 117, 6217–6226. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Mitchell, E.B.; Gorski, J.L.; Hollinger, C.; Hoppman, N.L. Two cases with de novo 3q26.31 microdeletion suggest a role for FNDC3B in human craniofacial development. Am. J. Med. Genet. A 2016, 170, 3276–3281. [Google Scholar] [CrossRef]
- Li, Q.; Wang, N.; Du, Z.; Hu, X.; Chen, L.; Fei, J.; Wang, Y.; Li, N. Gastrocnemius transcriptome analysis reveals domestication induced gene expression changes between wild and domestic chickens. Genomics 2012, 100, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef]
- Bowers, C.Y.; Momany, F.A.; Reynolds, G.A.; Hong, A. On the in vitro and in vivo activity of a new synthetic hexapeptide that acts on the pituitary to specifically release growth hormone. Endocrinology 1984, 114, 1537–1545. [Google Scholar] [CrossRef]
- Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Ghrelin and des-acyl ghrelin: Two major forms of rat ghrelin peptide in gastrointestinal tissue. Biochem. Biophys. Res. Commun. 2000, 279, 909–913. [Google Scholar] [CrossRef]
- Jönsson, E. The role of ghrelin in energy balance regulation in fish. Gen. Comp. Endocr. 2013, 187, 79–85. [Google Scholar] [CrossRef]
- Exton, J.H. New developments in phospholipase d. J. Biol. Chem. 1997, 272, 15579–15582. [Google Scholar] [CrossRef] [Green Version]
- Freyberg, Z.; Siddhanta, A.; Shields, D. “Slip, sliding away”: Phospholipase D and the Golgi apparatus. Trends Cell Biol. 2003, 13, 540–546. [Google Scholar] [CrossRef]
- Kodaki, T.; Yamashita, S. Cloning, expression, and characterization of a novel phospholipase D complementary DNA from rat brain. J. Biol. Chem. 1997, 272, 11408–11413. [Google Scholar] [CrossRef] [Green Version]
- Mcdermott, M.I.; Wang, Y.; Wakelam, M.J.O.; Bankaitis, V.A. Mammalian phospholipase D: Function, and therapeutics. Prog. Lipid Res. 2020, 78, 101018. [Google Scholar] [CrossRef]
- Lu, D.L.; Ma, Q.; Wang, J.; Li, L.Y.; Han, S.L.; Limbu, S.M.; Li, D.L.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. Fasting enhances cold resistance in fish through stimulating lipid catabolism and autophagy. J. Physiol. 2019, 597, 1585–1603. [Google Scholar] [CrossRef]
- Zheng, X.; Kuang, Y.; Lv, W.; Cao, D.; Sun, Z.; Sun, X. Genome-Wide association study for muscle fat content and abdominal fat traits in common carp (Cyprinus carpio). PLoS ONE 2016, 11, e169127. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).