
Identification of Proteins Responsible for High Activity of Cysteine Proteinase Inhibitor in the Blood of Nile Tilapia Oreochromis niloticus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Samples
2.2. Chemicals
2.3. Papain Inhibitory Activity in Different Tissues
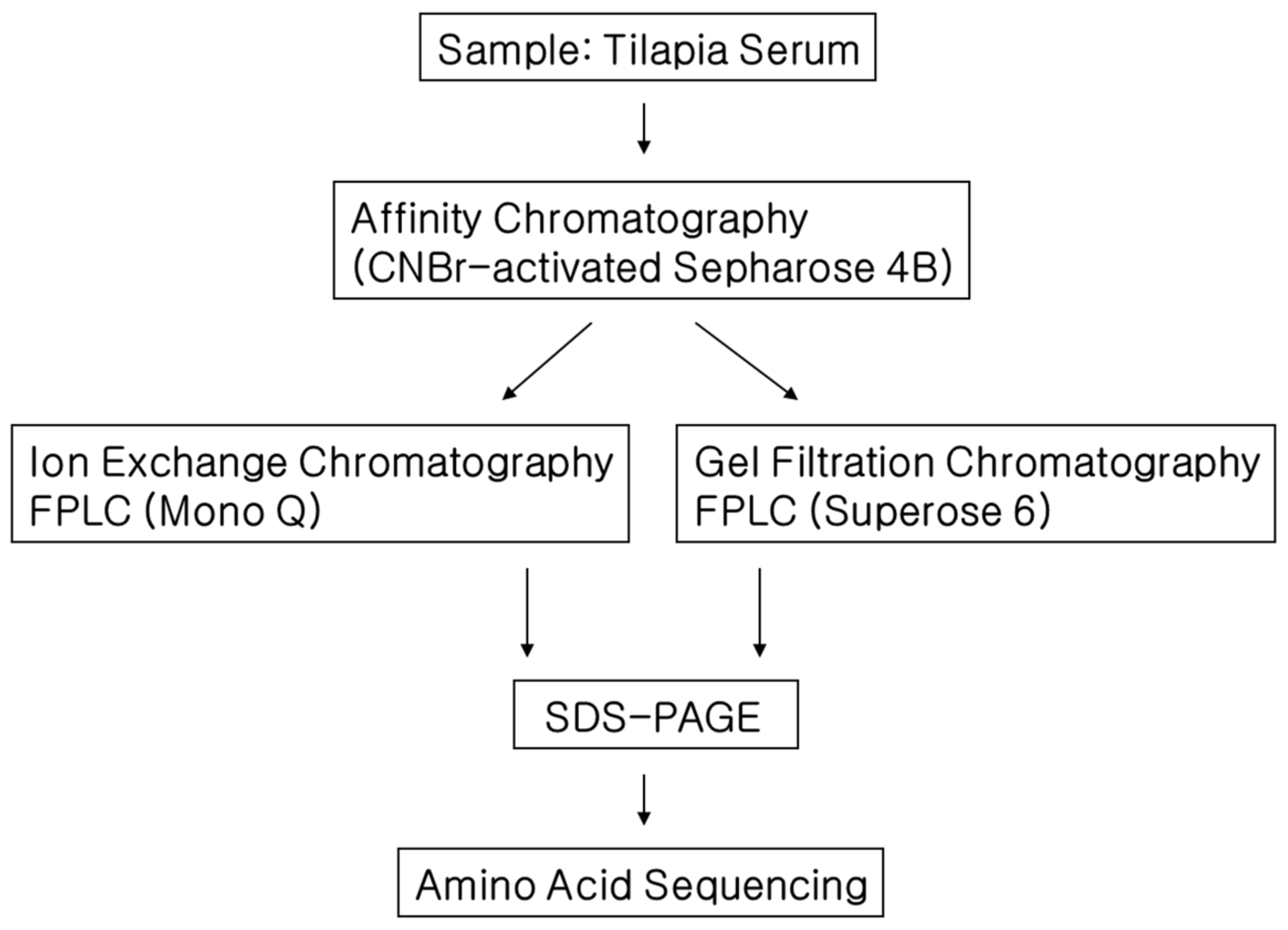
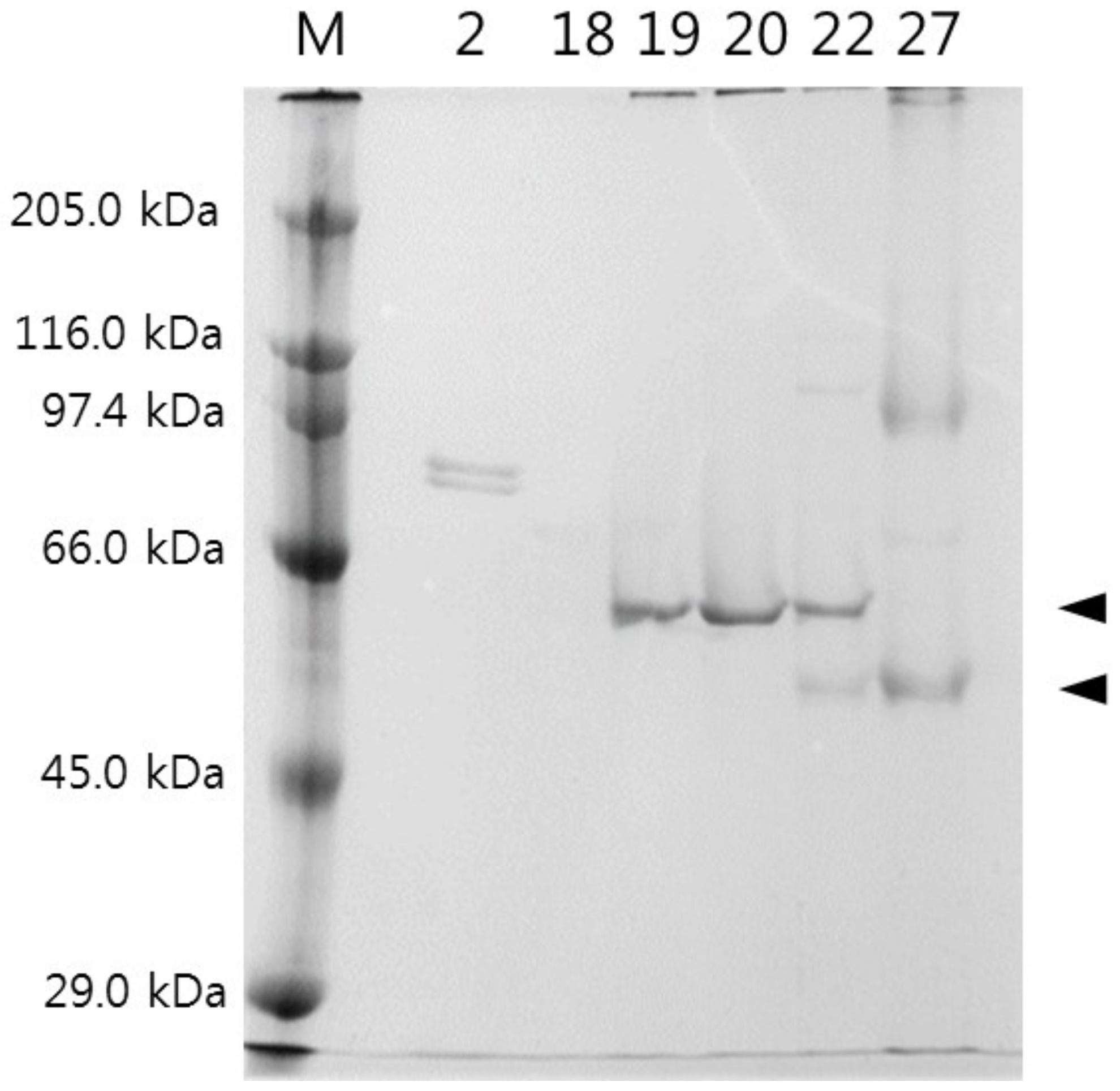
2.4. Isolation of CPIs from Tilapia Serum
2.5. Amino Acid Sequencing
3. Results and Discussion
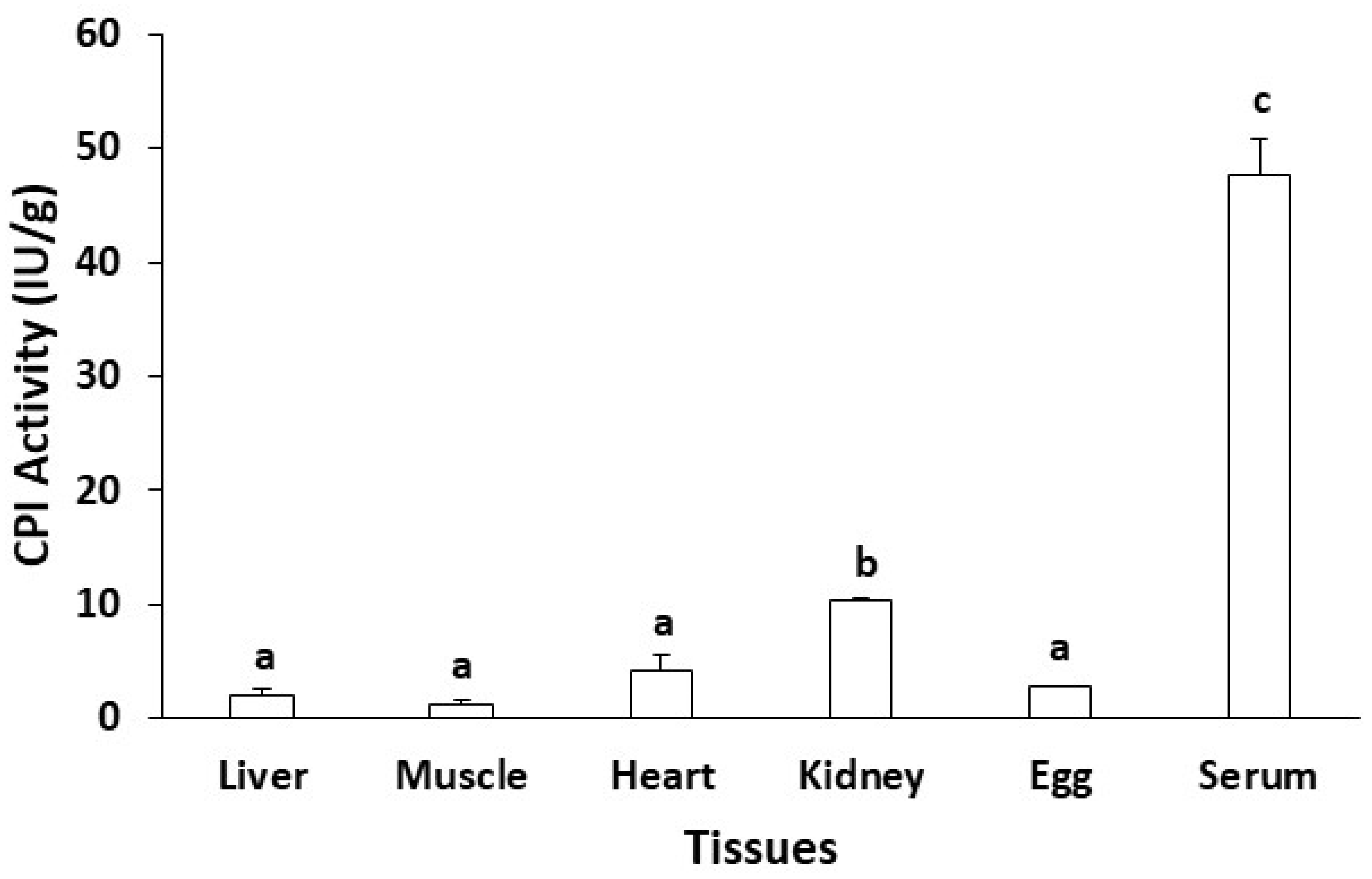
3.1. Tissue Distribution of CPI Activity
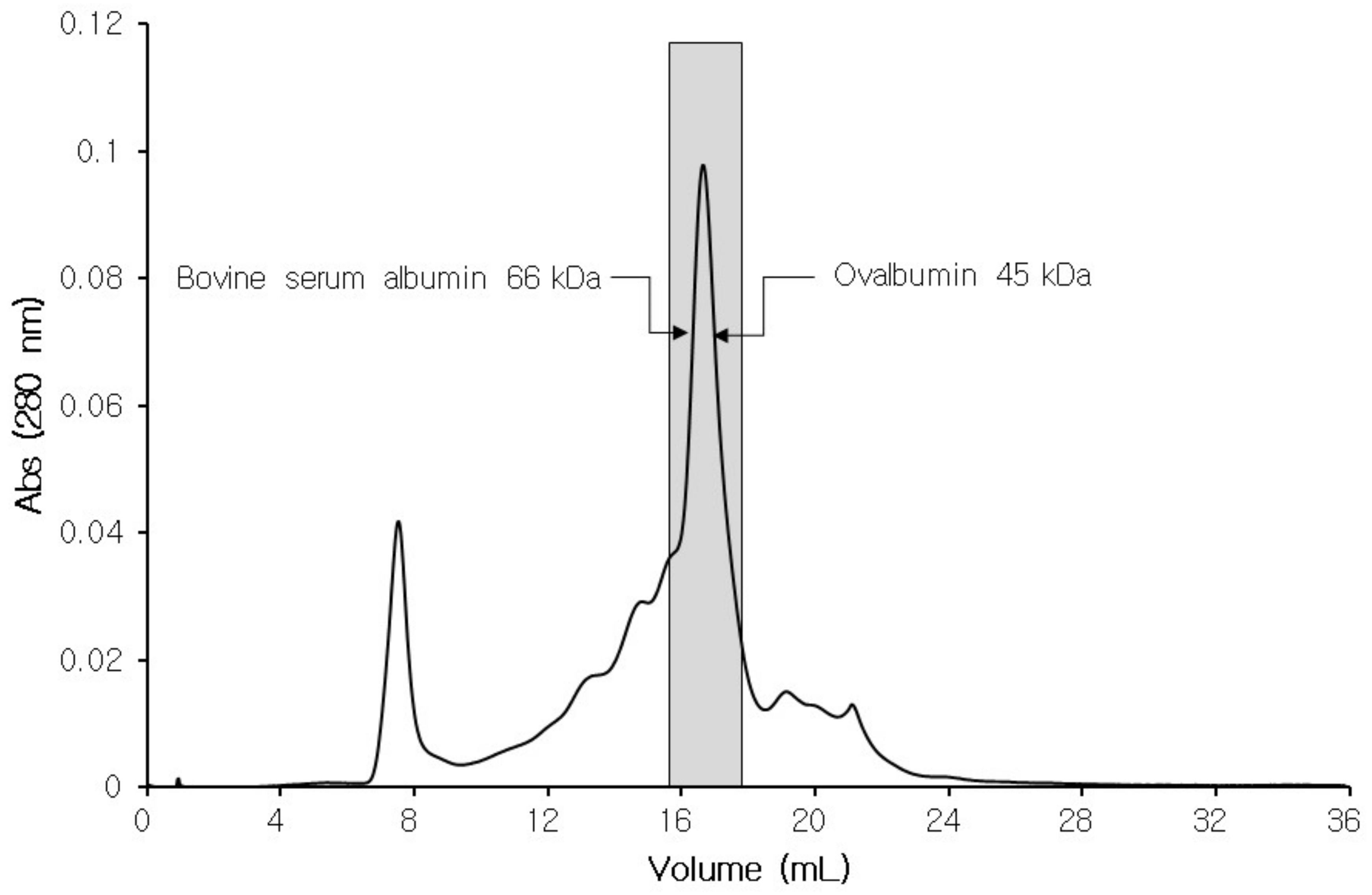
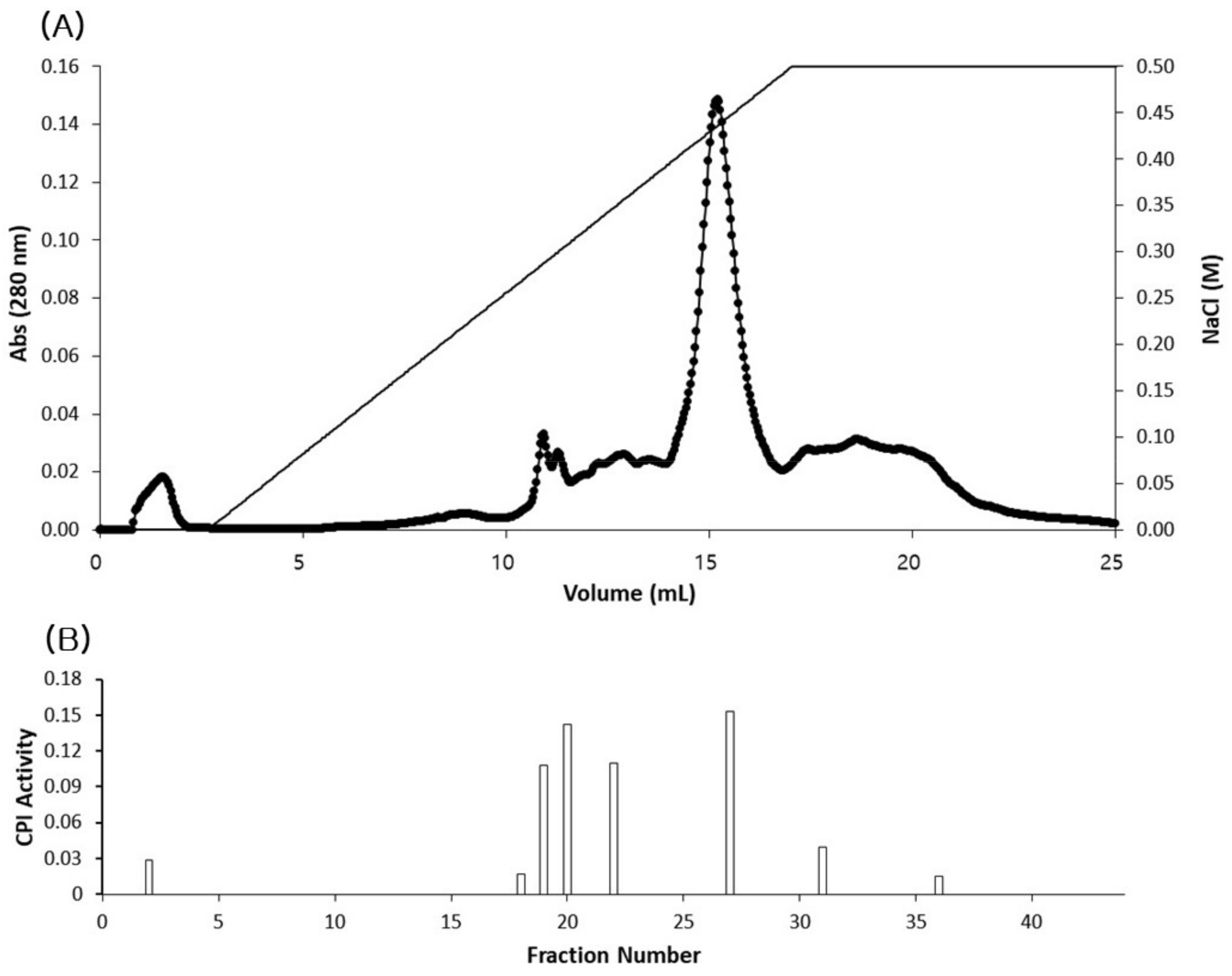
3.2. Isolation of CPIs from Tilapia Serum
3.3. Amino Acid Sequencing and Identification of Major Blood CPIs in Nile Tilapia
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bah, C.S.F.; Bekhit, A.E.A.; Carne, A.; McConnell, M.A. Slaughterhouse blood: An emerging source of bioactive compounds. Compr. Rev. Food Sci. Food Saf. 2013, 12, 314–331. [Google Scholar] [CrossRef]
- Li, D.K.; Lin, H.; Kim, S.M. Effect of rainbow trout (Oncorhynchus mykiss) plasma protein on the gelation of Alaska pollock (Theragra chalcogramma) surimi. J. Food Sci. 2008, 73, C227–C234. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.R.; Park, J.W. Effect of salmon plasma protein on Pacific whiting surimi gelation under various ohmic heating conditions. LWT-Food Sci. Technol. 2015, 61, 309–315. [Google Scholar] [CrossRef]
- Fowler, M.R.; Park, J.W. Salmon blood plasma: Effective inhibitor of protease-laden Pacific whiting surimi and salmon mince. Food Chem. 2015, 176, 448–454. [Google Scholar] [CrossRef]
- Rawdkuen, S.; Lanier, T.C.; Visessanguan, W.; Benjakul, S. Cysteine proteinase inhibitor from chicken plasma: Fractionation, characterization and autolysis inhibition of fish myofibrillar proteins. Food Chem. 2007, 101, 1647–1657. [Google Scholar] [CrossRef]
- Brown, W.M.; Dziegielewska, K.M. Friends and relations of the cystatin superfamily-new members and their evolution. Protein Sci. 1997, 6, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Kwon, H.C.; Kwon, J.Y. Thermal stability of cysteine proteinase inhibitor of tilapia (Oreochromis niloticus) egg and serum. Dev. Reprod. 2006, 10, 263–269. [Google Scholar]
- Nopianti, R.; Baehaki, A.; Ridhowati, S.; Suhartono, M.T. Protease Inhibitory activity and protein analysis of catfish (Pangasius hypothalamus) and swamp eel (Monopterus albus) blood plasma. Pertanika J. Trop. Agric. Sci. 2019, 42, 155–164. [Google Scholar]
- Taglieri, N.; Koenig, W.; Kaski, J.C. Cystatin C and cardiovascular risk. Clin. Chem. 2009, 55, 1932–1943. [Google Scholar] [CrossRef] [Green Version]
- Mashiko, H.; Takahashi, H. Bullfrog plasma cysteine proteinase inhibitor (CPI). Immunophamacology 1996, 32, 91–93. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Lu, Y.; Lu, C. Transcriptome profiling of zebrafish infected with Streptococcus suis. Microb. Pathog. 2010, 48, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.A.; Arnold, G.J.; Nynca, J.; Frohlich, T.; Otte, K.; Ciereszko, A. Characterization of carp seminal plasma proteome in relation to blood plasma. J. Proteomics 2014, 98, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Nakharuthai, C.; Rodrigues, P.M.; Schrama, D.; Kumkhong, S.; Boonanuntanasarn, S. Effects of different dietary vegetable lipid sources on health status in Nile tilapia (Oreochromis niloticus): Haematological indices, immune response parameters and plasma proteome. Animals 2020, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- Dubin, G. Proteinaceous cysteine protease inhibitors. Cell Mol. Life Sci. 2005, 62, 653–669. [Google Scholar] [CrossRef]
- Ghys, L.; Paepe, D.; Smets, P.; Lefebvre, H.; Delanghe, J.; Daminet, S. Cystatin C: A new renal marker and its potential use in small animal medicine. J. Vet. Intern. Med. 2014, 28, 1152–1164. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; An, H.; Seymour, T.A.; Bradford, C.S.; Morrissey, M.T.; Bailey, G.S.; Helmrich, A.; Barnes, D.W. Molecular cloning, sequence analysis and expression distribution of rainbow trout (Oncorhynchus mykiss) cystatin C. Comp. Biochem. Physiol. Part B 1998, 121, 135–143. [Google Scholar] [CrossRef]
- Li, D.K.; Lin, H.; Kim, S.M. Purification and characterization of a cysteine protease inhibitor from chum salmon (Oncorhynchus keta) plasma. J. Agric. Food Chem. 2008, 56, 106–111. [Google Scholar] [CrossRef]
- Li, C.; Gao, C.; Fu, Q.; Su, B.; Chen, J. Identification and expression analysis of fetuin B (FETUB) in turbot (Scophthalmus maximus L.) mucosal barriers following bacterial challenge. Fish Shellfish Immunol. 2017, 68, 386–394. [Google Scholar] [CrossRef]
- Synnes, M. Purification and characterization of two cysteine proteinase inhibitors from the skin of Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Part B 1998, 121, 257–264. [Google Scholar] [CrossRef]
- Yamashita, M.; Konagaya, S. A novel cysteine protease inhibitor of the egg of chum salmon, containing a cysteine-rich thyroglobulin-motif. J. Biol. Chem. 1996, 271, 1282–1284. [Google Scholar] [CrossRef] [Green Version]
- Nurhayati, T.; Rusyadi, S.; Suwandi, R.; Nugraha, R. Purification and characterization of a cathepsin inhibitor from catfish (Pangasius sp.) of Indonesian water. Int. Food Res. J. 2013, 20, 941–946. [Google Scholar]
- Li, F.; An, H.; Seymour, T.A.; Barnes, D.W. Rainbow trout (Oncorhynchus mykiss) cystatin C: Expression in Escherichia coli and properties of the recombinant protease inhibitor. Comp. Biochem. Physiol. Part B 2000, 123, 493–502. [Google Scholar] [CrossRef]
- Barrett, A.J. A new assay for cathepsin B1 and other thiol proteinases. Anal. Biochem. 1972, 47, 280–293. [Google Scholar] [CrossRef]
- Barrett, A.J.; Kirschke, H. Cathepsin B, cathepsin H, and cathepsin L. Meth. Enzymol. 1981, 80, 535–561. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Hakansson, K.; Huh, C.; Grubb, A.; Karlsson, S.; Abrahamson, M. Mouse and rat cystatin C: Escherichia coli Production, characterization and tissue distribution. Comp. Biochem. Physiol. 1996, 114B, 303–311. [Google Scholar] [CrossRef]
- Li, R.; Tan, X.; Li, S.; Jin, Y.; Li, S.; Li, S.; Takala, T.M.; Saris, P.E.J. Cloning, expression, characterization, and tissue distribution of cystatin C from silver carp (Hypophthalmichthys molitrix). J. Agric. Food Chem. 2021, 69, 5144–5154. [Google Scholar] [CrossRef]
- Xiao, P.-P.; Hu, Y.-H.; Sun, L. Scophthalmus maximus Cystatin B enhances head kidney macrophage-mediated bacterial killing. Dev. Comp. Immunol. 2010, 34, 1237–1241. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Coulis, G.; Ouali, A. Role of muscle endopeptidases and their inhibitors in meat tenderness. Trends Food Sci. Technol. 2002, 13, 400–421. [Google Scholar] [CrossRef]
- Elzanowski, A.; Barker, W.C.; Hunt, L.T.; Seibel-Ross, E. Cystatin domains in alpha-2-HS glycoprotein and fetuin. FEBS Lett. 1988, 227, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Jerala, R.; Trstenjak-Prebanda, M.; Kroon-Zitko, L.; Lenarcic, B.; Turk, V. Mutations in the QVVAG region of the cysteine proteinase inhibitor stefin B. Biol. Chem. Hoppe Seyler 1990, 371, 157–160. [Google Scholar] [PubMed]
- Wong, M.K.S.; Takei, Y. Lack of plasma kallikrein-kinin system cascade in teleosts. PLoS ONE 2013, 8, e81057. [Google Scholar]
- Brown, W.M.; Saunders, N.R.; Mollgard, K.; Diegielewska, K.M. Fetuin–an old friend revisited. BioEssays 1992, 14, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Nynca, J.; Slowinska, M.; Dietrich, M.A.; Bilinska, B.; Kotula-Balak, M.; Ciereszko, A. Isolation and identification if fetuin-B-like protein from rainbow trout seminal plasma and its localization in the reproductive system. Comp. Biochem. Physio Part B 2011, 158, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Hibbetts, K.; Hines, B.; Williams, D. An overview of proteinase inhibitors. J. Vet. Intern. Med. 1999, 13, 302–308. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide No. | Sequences |
|---|---|
| 20-1 | ASLFQPLWGEGK * |
| 20-2 | ATAQVVAGSVYR * |
| 20-3 | ADATQDVGTLMANDPFMAAPK ** |
| 27-1 | ADATQDVGTLMANDPFMAAPK ** |
| 27-2 | EGYLFSLHR * |
| 27-3 | YNSFSDSTHLFSLHNLDR *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mun, S.H.; Kwon, J.Y. Identification of Proteins Responsible for High Activity of Cysteine Proteinase Inhibitor in the Blood of Nile Tilapia Oreochromis niloticus. Fishes 2022, 7, 161. https://doi.org/10.3390/fishes7040161
Mun SH, Kwon JY. Identification of Proteins Responsible for High Activity of Cysteine Proteinase Inhibitor in the Blood of Nile Tilapia Oreochromis niloticus. Fishes. 2022; 7(4):161. https://doi.org/10.3390/fishes7040161
Chicago/Turabian StyleMun, Seong Hee, and Joon Yeong Kwon. 2022. "Identification of Proteins Responsible for High Activity of Cysteine Proteinase Inhibitor in the Blood of Nile Tilapia Oreochromis niloticus" Fishes 7, no. 4: 161. https://doi.org/10.3390/fishes7040161
APA StyleMun, S. H., & Kwon, J. Y. (2022). Identification of Proteins Responsible for High Activity of Cysteine Proteinase Inhibitor in the Blood of Nile Tilapia Oreochromis niloticus. Fishes, 7(4), 161. https://doi.org/10.3390/fishes7040161