
Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Diets
2.3. Bioassay
2.4. Growth Indices
2.5. Digestive Enzyme Activities
2.6. RNA Extraction and cDNA Synthesis
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
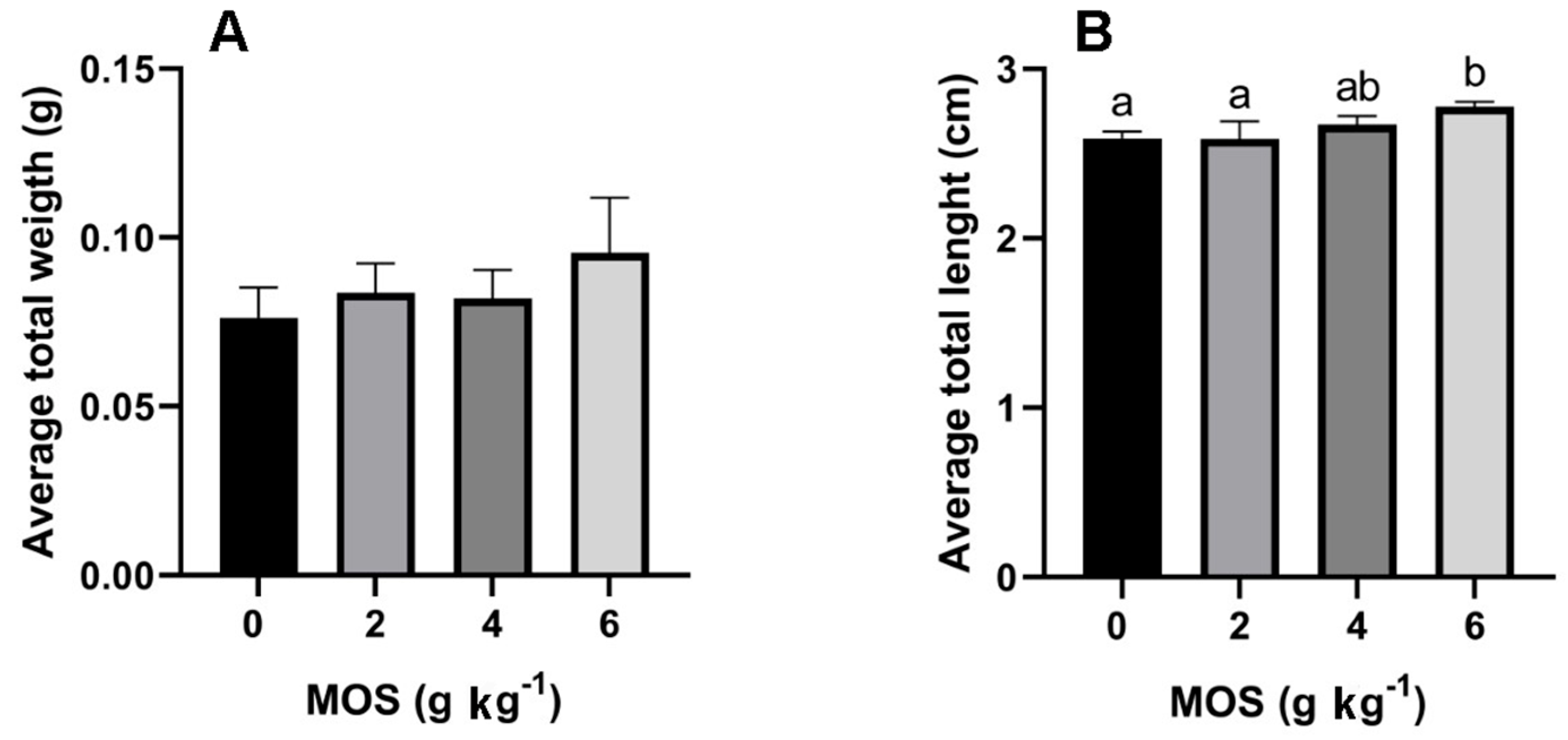
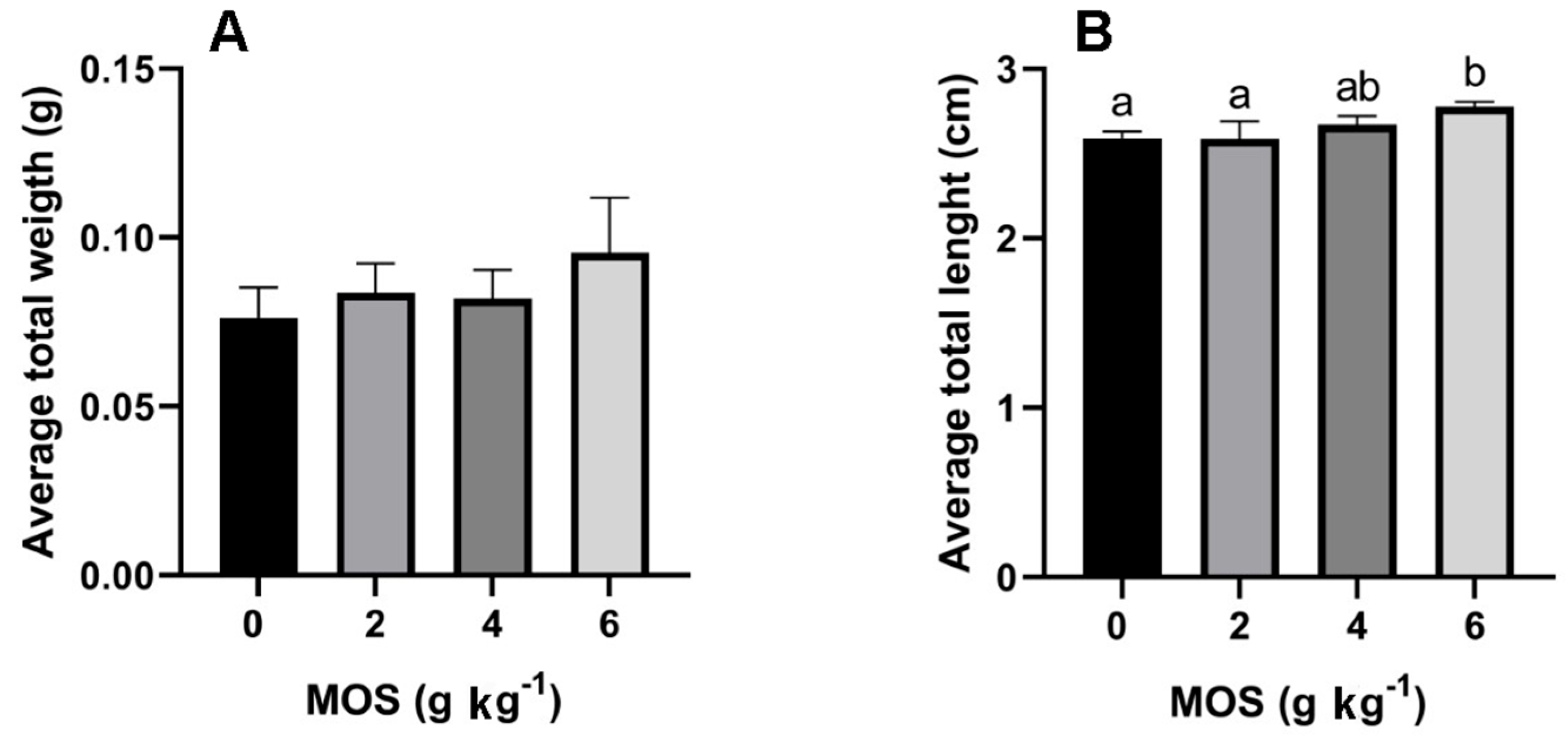
3.1. Growth Parameters Influenced by MOS Supplementation
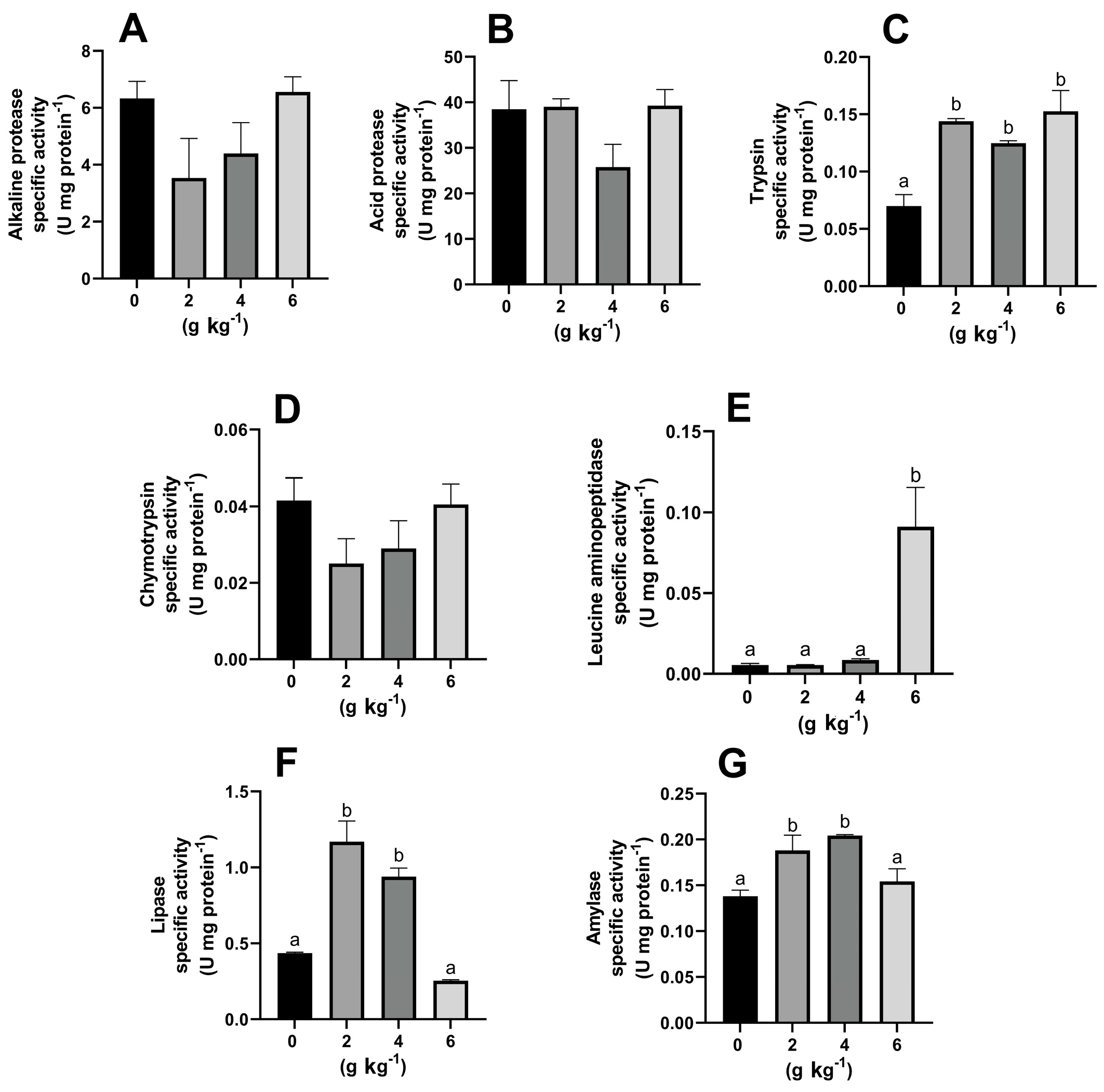
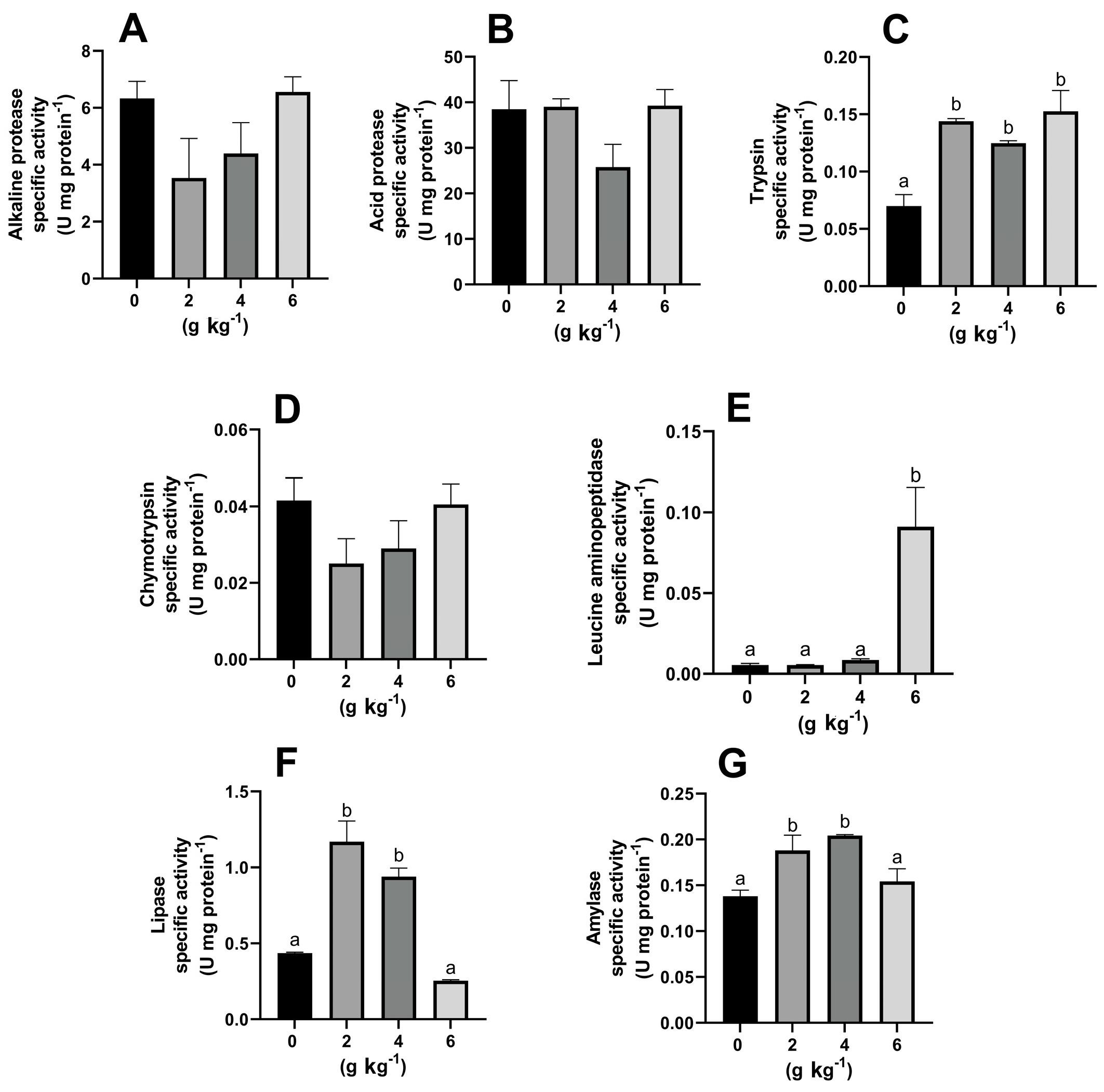
3.2. Effect of MOS on the Activity of Digestive Enzymes
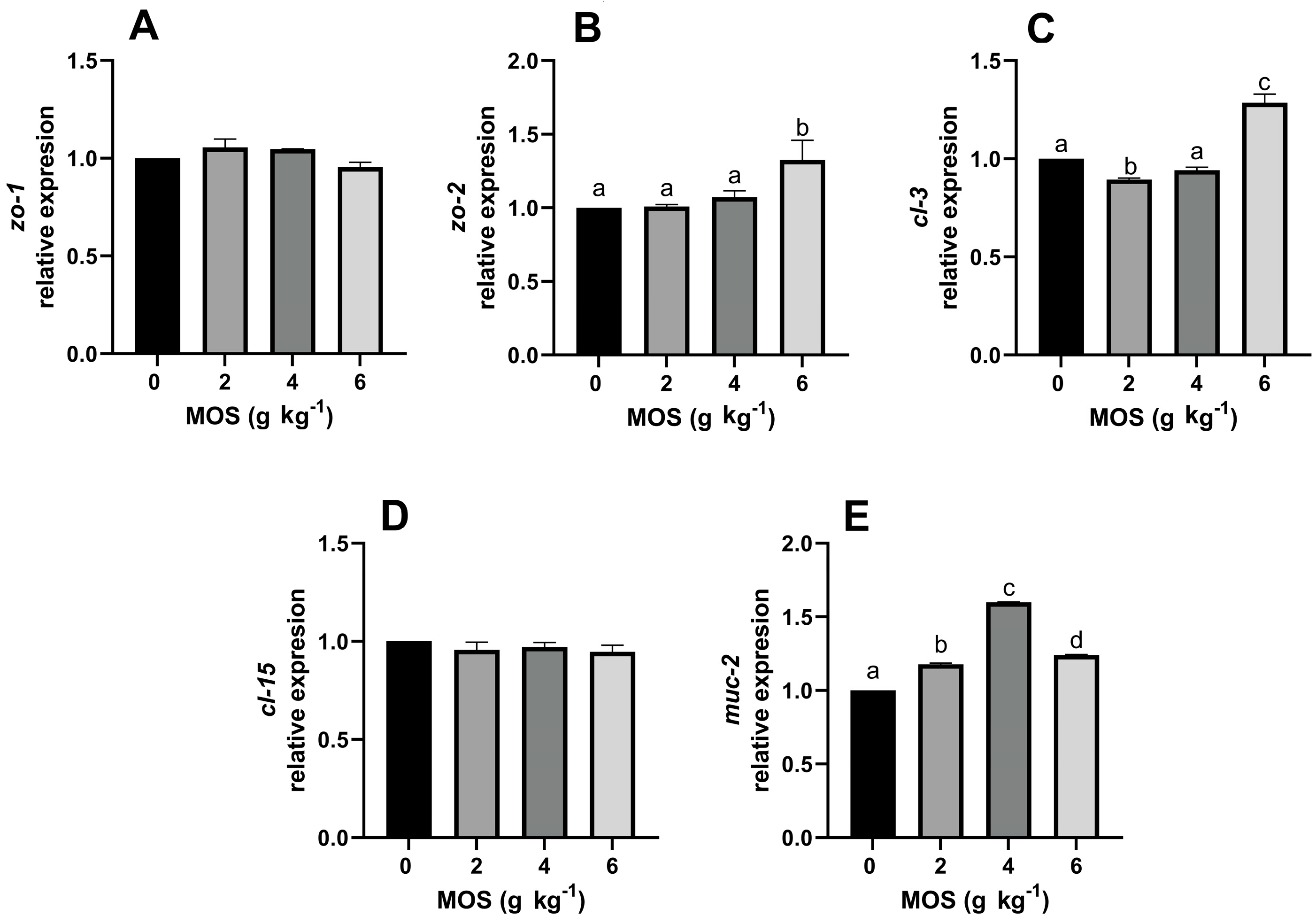
3.3. Effect of MOS on Gene Expression of Intestinal Barrier Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jobling, M. Fish nutrition research: Past, present and future. Aquac. Int. 2016, 24, 767–786. [Google Scholar] [CrossRef]
- Montalto, M.; D’Onofrio, F.D.; Gallo, A.; Cazzato, A.; Gasbarrini, G. Intestinal microbiota and its functions. Dig. Liver Dis. 2009, 3, 30–34. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Gainza, O.; Romero, J. Effect of mannan oligosaccharides on the microbiota and productivity parameters of Litopenaeus vannamei shrimp under intensive cultivation in Ecuador. Sci. Rep. 2020, 10, 2719. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Olsen, R.E.; Gifstad, T.Ø.; Dalmo, R.A.; Amlund, H.; Hemre, G.-I.; Bakke, A.M. Prebiotics in aquaculture: A review. Aquac. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Lu, J.; Qi, C.; Limbu, S.M.; Han, F.; Yang, L.; Wang, X.; Qin, J.G.; Chen, L. Dietary mannan oligosaccharide (MOS) improves growth performance, antioxidant capacity, non-specific immunity and intestinal histology of juvenile Chinese mitten crabs (Eriocheir sinensis). Aquaculture 2019, 510, 337–346. [Google Scholar] [CrossRef]
- Ren, Z.; Wang, S.; Cai, Y.; Wu, Y.; Tian, L.; Wang, S.; Jiang, L.; Guo, W.; Sun, Y.; Zhou, Y. Effects of dietary mannan oligosaccharide supplementation on growth performance, antioxidant capacity, non-specific immunity and immune- related gene expression of juvenile hybrid grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀). Aquaculture 2020, 523, 735195. [Google Scholar] [CrossRef]
- Mansour, M.R.; Akrami, R.; Ghobadi, S.H.; Denji, K.A.; Ezatrahimi, N.; Gharaei, A. Effect of dietary mannan oligosaccharide (MOS) on growth performance, survival, body composition, and some hematological parameters in giant sturgeon juvenile (Huso huso Linnaeus, 1754). Fish Physiol. Biochem. 2012, 38, 829–835. [Google Scholar] [CrossRef]
- Márquez-Couturier, G.; Vázquez-Navarrete, C.J. Estado del arte de la biología y cultivo de pejelagarto (Atractosteus tropicus). Agroproductividad 2015, 8, 44–51. [Google Scholar]
- Frías-Quintana, C.A.; Domínguez-Lorenzo, J.; Álvarez-González, C.A.; Tovar-Ramírez, D.; Martínez-García, R. Using cornstarch in microparticulate diets for larvicultured tropical gar (Atractosteus tropicus). Fish Physiol. Biochem. 2016, 42, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Rodríguez, K.; Álvarez-González, C.; Peña-Marín, E.; Vega-Villasante, F.; Martínez-García, R.; Camarillo-Coop, S. Effect of β-Glucans in diets on growth, survival, digestive enzyme activity, and immune system and intestinal barrier gene expression for Tropical Gar (Atractosteus tropicus) juveniles. Fishes 2018, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Sepúlveda-Quiroz, C.A.; Peña-Marín, E.S.; Pérez-Morales, A.; Martínez-García, R.; Alvarez-Villagomez, C.S.; Maytorena-Verdugo, C.I.; Camarillo-Coop, S.; Vissio, P.G.; Pérez Sirkin, D.; Tovar-Ramírez, D.; et al. Fructooligosaccharide supplementation in diets for tropical gar (Atractosteus tropicus) juvenile: Effects on morphophysiology and intestinal barrier function. Aquac. Res. 2020, 52, 37–50. [Google Scholar] [CrossRef]
- Nájera-Arzola, I.C.; Álvarez-González, C.A.; Frías-Quintana, C.A.; Peña, E.; Martínez-García, R.; Camarillo-Coop, S.; Méndez-Marín, O.; Gisbert, E. Evaluation of Mannan oligosaccharides (MOS) in balanced diets for tropical gar juveniles (Atractosteus tropicus). Hidrobiológica 2018, 28, 239–246. [Google Scholar] [CrossRef]
- Mexican Standard NOM-062-ZOO-1999; Diario Oficial de la Federación. Ministry of Agriculture, Livestock, Rural Development, Fisheries and Food: Ciudad de Mexico, Mexico, 1999. Available online: http://publico.senasica.gob.mx/?doc=743 (accessed on 15 May 2022).
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- Frías-Quintana, C.A.; Márquez-Couturier, G.; Alvarez-González, C.A.; Nolasco-Soria, H.; Galaviz-Espinosa, M.A.; Martínez-García, R.; Camarillo-Coop, S.; Martínez-Yañez, R.; Gisbert, E. Development of digestive tract and enzyme activities during the early ontogeny of the tropical gar Atractosteus tropicus. Fish Physiol. Biochem. 2015, 41, 1075–1091. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Walter, H.E. Proteinases: Methods with hemoglobin, casein and azocoll as substrates. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1984; Volume 5, pp. 270–277. [Google Scholar]
- García-Carreño, F.L.; Hernández-Cortés, M.; Haard, N.F. Enzymes with peptidase and proteinase activity from the digestive systems of a freshwater and a marine decapod. J. Agric. Food Chem. 1994, 42, 1456–1461. [Google Scholar] [CrossRef]
- Maroux, S.; Louvard, D.; Barath, J. The aminopeptidase from hog intestinal brush border. Biochim. Biophys. Acta (BBA)-Enzymol. 1973, 321, 282–295. [Google Scholar] [CrossRef]
- Versaw, W.; Cuppett, S.L.; Winters, D.D.; Williams, L.E. An improved colorimetric assay for bacterial lipase in nonfat dry milk. J. Food Sci. 1989, 54, 232–254. [Google Scholar] [CrossRef]
- Robyt, J.F.; Whelan, W. Amylases. In Starch and Its Derivates; Radley, J.A., Ed.; Chapman and Hall: London, UK, 1968. [Google Scholar]
- Pérez-Jiménez, G.M. Efecto del Prebiótico Fructooligosacárido (FOS) Sobre la Fisiología Digestiva y Barrera Intestinal en Larvas de Pejelagarto (Atractosteus tropicus). Master’s Thesis, División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, Mexico, 2020. [Google Scholar]
- Martínez-Burguete, T.; Peña-Marín, E.S.; García-Gasca, A.; Álvarez-González, C.A.; Llera-Herrera, R. Nutrigenomic marker discovery by de novo transcriptomic sequencing during early development of the tropical gar (Atractosteus tropicus). Aquac. Res. 2021, 52, 3829–3842. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2(−delta delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gatlin, D.M., III; Peredo, A.M. Prebiotics and Probiotics: Definitions and Applications; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2012; Volume 4711, pp. 1–8. [Google Scholar]
- Roberfroid, M. Prebiotics: The Concept Revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, P.Y.; Kim, W.K. Review: Roles of Prebiotics in Intestinal Ecosystem of Broilers. Front. Vet. Sci. 2018, 5, 245. [Google Scholar] [CrossRef]
- Torrecillas, S.; Montero, D.; Izquierdo, M. Improved health and growth of fish fed mannan oligosaccharides: Potential mode of action. Fish Shellfish Immunol. 2014, 36, 525–544. [Google Scholar] [CrossRef]
- Grisdale-Helland, B.; Helland, S.J.; Gatlin, D.M., III. The effects of dietary supplementation with mannanoligosaccharide, fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon (Salmo salar). Aquaculture 2008, 283, 163–167. [Google Scholar] [CrossRef]
- Salze, G.; McLean, E.; Schwarz, M.H.; Craig, S.R. Dietary mannan oligosaccharide enhances salinity tolerance and gut development of larval cobia. Aquaculture 2008, 274, 148–152. [Google Scholar] [CrossRef]
- Talpur, A.D.; Munir, M.B.; Mary, A.; Hashim, R. Dietary probiotics and prebiotics improved food acceptability, growth performance, haematology and immunological parameters and disease resistance against Aeromonas hydrophila in snakehead (Channa striata) fingerlings. Aquaculture 2014, 426–427, 14–20. [Google Scholar] [CrossRef]
- Lu, Z.Y.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Mannan oligosaccharides improved growth performance and antioxidant capacity in the intestine of on-growing grass carp (Ctenopharyngodon idella). Aquac. Rep. 2020, 17, 100313. [Google Scholar] [CrossRef]
- Méndez-Pérez, R.; García-López, R.; Bautista-López, J.S.; Vázquez-Castellanos, J.; Álvarez-Gonzales, C.; Peña-Marín, E.; Baltierra-Trejo, E.; Adams-Schroeder, R.; Domínguez-Rodríguez, V.; Melgar-Valdés, C.; et al. High-throughput sequencing of the 16S rRNA gene to analyze the gut microbiome in juvenile and adult tropical gar (Atractosteus tropicus). Lat. Am. J. Aquat. Res. 2020, 48, 456–479. [Google Scholar] [CrossRef]
- Mohammadian, T.; Nasirpour, M.; Tabandeh, M.R.; Mesbah, M. Synbiotic effects of β-glucan, mannan oligosaccharide and Lactobacillus casei on growth performance, intestine enzymes activities, immune-hematological parameters and immune-related gene expression in common carp, Cyprinus carpio: An experimental infection with Aeromonas hydrophila. Aquaculture 2019, 511, 634197. [Google Scholar] [CrossRef]
- Ye, J.D.; Wang, K.; Li, F.D.; Sun, Y.Z. Single or combined effects of fructo- and mannan oligosaccharide supplements and Bacillus clausii on the growth, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquac. Nutr. 2011, 17, e902e11. [Google Scholar] [CrossRef]
- Wang, J.; Lei, P.; Gamil, A.A.A.; Lagos, L.; Yue, Y.; Schirmer, K.; Mydland, L.T.; Øverland, M.; Krogdahl, Å.; Kortner, T.M. Rainbow Trout (Oncorhynchus Mykiss) Intestinal Epithelial Cells as a Model for Studying Gut Immune Function and Effects of Functional Feed Ingredients. Front. Immunol. 2019, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- David-Ruales, C.A.; Machado-Fracalossi, D.; Vásquez-Torres, W. Early development in fish larvae, key for starting exogenous feeding. Rev. Lasallista Investig. 2018, 15, 180–194. [Google Scholar] [CrossRef]
- Kokot, A.M.; Jarocka-Cyrta, E.; Drabińska, N. Overview of the Importance of Biotics in Gut Barrier Integrity. Int. J. Mol. Sci. 2022, 23, 2896. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Heazlewood, C.; Cook, M.; Eri, R.; Price, G.; Tauro, S.; Taupin, D.; Thornton, D.; Png, C.; Crockford, T.; Cornall, R.; et al. Aberrant mucin assembly in mice causes endoplasmic reticulum stress and spontaneous inflammation resembling ulcerative colitis. PLoS Med. 2008, 5, e54. [Google Scholar] [CrossRef] [Green Version]
- Findley, M.K.; Koval, M. Regulation and roles for claudin-family tight junction proteins. IUBMB Life 2010, 61, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, X.; Wang, Y.; Huang, Y.; Wang, C. Effects of alternate feeding between fish meal and novel protein diets on the intestinal health of juvenile largemouth bass (Micropterus salmoides). Aquac. Rep. 2022, 23, 101023. [Google Scholar] [CrossRef]
- González-Marisal, L.; Miranda, J.; Raya-Sandino, A.; Domínguez-Calderón, A.; Cuellar-Perez, F. ZO-2, a tight junction protein involved in gene expression, proliferation, apoptosis, and cell size regulation. Ann. N. Y. Acad. Sci. 2017, 1397, 35–53. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Ingredients (g kg−1)/MOS (g kg−1) | 0 | 2 | 4 | 6 |
|---|---|---|---|---|
| Fish meal a | 305.4 | 305.4 | 305.4 | 305.4 |
| Poultry meal a | 150 | 150 | 150 | 150 |
| Pork meal a | 150 | 150 | 150 | 150 |
| Soybean meal a | 150 | 150 | 150 | 150 |
| Starch b | 123.7 | 121.7 | 119.7 | 117.7 |
| Soybean oil c | 79.9 | 79.9 | 79.9 | 79.9 |
| MOS d | 0 | 2 | 4 | 6 |
| Mineral premix e | 5 | 5 | 5 | 5 |
| Vitamin premix e | 10 | 10 | 10 | 10 |
| Grenetin f | 20 | 20 | 20 | 20 |
| Vitamin C g | 5 | 5 | 5 | 5 |
| Vitamin E g | 1 | 1 | 1 | 1 |
| Proximal composition g/100g dry matter | ||||
| Protein | 44.20 | 44.00 | 44.15 | 44.05 |
| Lipids | 14.90 | 15.10 | 15.10 | 15.00 |
| Ash | 12.44 | 12.40 | 12.43 | 12.38 |
| NFE 1 | 28.46 | 28.5 | 28.32 | 28.57 |
| Target Gene | Primer Sequence (5′-3′) | R2 | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| zo-1 | FW: TGTGCCTCAGATCACTCCACRV: AAAGGCAGAGGGTTGGCTTC | 0.95 | 123 | [23] |
| zo-2 | FW: TACCCATGGAAAATGTGCCTCARV: CGGGGTCTCTTCACGGTAAT | 0.98 | 88 | [23] |
| claudin-3 | FW: CCTGTATATCGGCTGGGCTGRV: TGCAAGCTAACGACTACGCA | 0.91 | 285 | [23] |
| claudin-15 | FW: ATCCCGGGACAAAGTACGAGRV:CAGATCGCTAGCAAGGCAGA | 0.93 | 70 | [23] |
| muc-2 | FW: GGCCTCCTCAAGAGCACGGTGRV:TCTGCACGCTGGAGCACTCAATG | - | 100 | [13] |
| β-actin | FW: GGACTTTGAGCAGGAGATGGRV:GACGGAGTATTTACGCTCTGG | - | 355 | [13] |
| Parameter/MOS (g kg−1) | 0 | 2 | 4 | 6 |
|---|---|---|---|---|
| AWG (g fish−1) | 0.042 ± 0.009 | 0.056 ± 0.009 | 0.055 ± 0.009 | 0.067 ± 0.016 |
| WG (%) | 122.83 ± 26.28 a | 196.97 ± 30.74 a | 202.29 ± 31.62 a | 232.69 ± 57.30 b |
| SGR (% day−1) | 3.98 ± 0.60 a | 5.42 ± 0.53 a | 5.51 ± 0.54 a | 5.96 ± 0.84 b |
| FCR | 3.94 ± 0.92 | 2.93 ± 0.49 | 2.97 ± 0.51 | 2.49 ± 0.58 |
| K | 0.47 | 0.40 | 0.33 | 0.37 |
| S (%) | 24.87 ± 1.26 a | 15.14 ± 2.57 b | 13.16 ± 0.72 b | 25.28 ± 1.67 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maytorena-Verdugo, C.I.; Peña-Marín, E.S.; Alvarez-Villagómez, C.S.; Pérez-Jiménez, G.M.; Sepúlveda-Quiroz, C.A.; Alvarez-González, C.A. Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes. Fishes 2022, 7, 127. https://doi.org/10.3390/fishes7030127
Maytorena-Verdugo CI, Peña-Marín ES, Alvarez-Villagómez CS, Pérez-Jiménez GM, Sepúlveda-Quiroz CA, Alvarez-González CA. Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes. Fishes. 2022; 7(3):127. https://doi.org/10.3390/fishes7030127
Chicago/Turabian StyleMaytorena-Verdugo, Claudia I., Emyr S. Peña-Marín, Carina S. Alvarez-Villagómez, Graciela M. Pérez-Jiménez, César A. Sepúlveda-Quiroz, and Carlos A. Alvarez-González. 2022. "Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes" Fishes 7, no. 3: 127. https://doi.org/10.3390/fishes7030127
APA StyleMaytorena-Verdugo, C. I., Peña-Marín, E. S., Alvarez-Villagómez, C. S., Pérez-Jiménez, G. M., Sepúlveda-Quiroz, C. A., & Alvarez-González, C. A. (2022). Inclusion of Mannan-Oligosaccharides in Diets for Tropical Gar Atractosteus tropicus Larvae: Effects on Growth, Digestive Enzymes, and Expression of Intestinal Barrier Genes. Fishes, 7(3), 127. https://doi.org/10.3390/fishes7030127