
Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus labrax Juvenile Reared in Tanks

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Settings
2.2. Group-Based Risk-Taking Test
2.3. Statistical Analyses
2.4. Ethical Statement
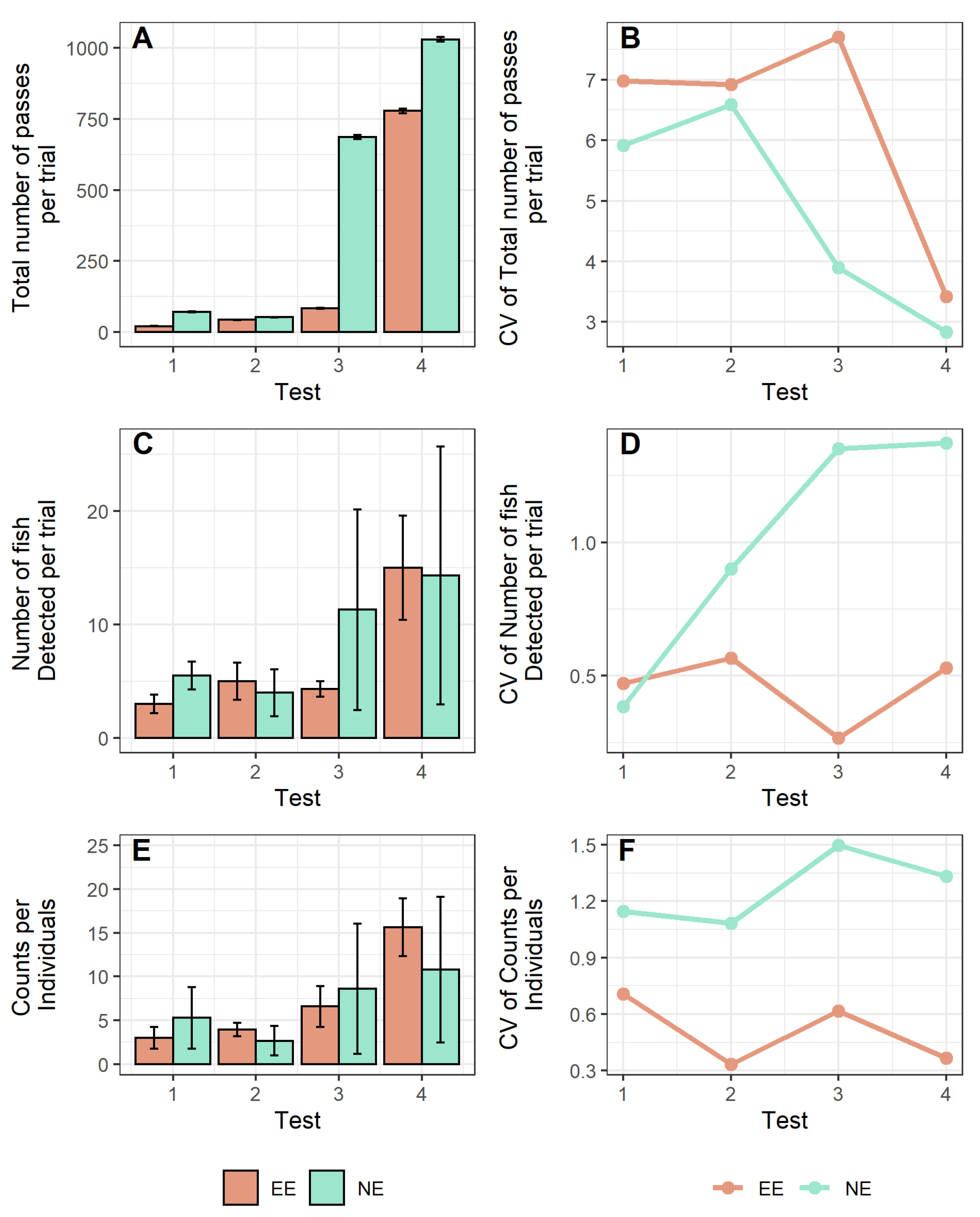
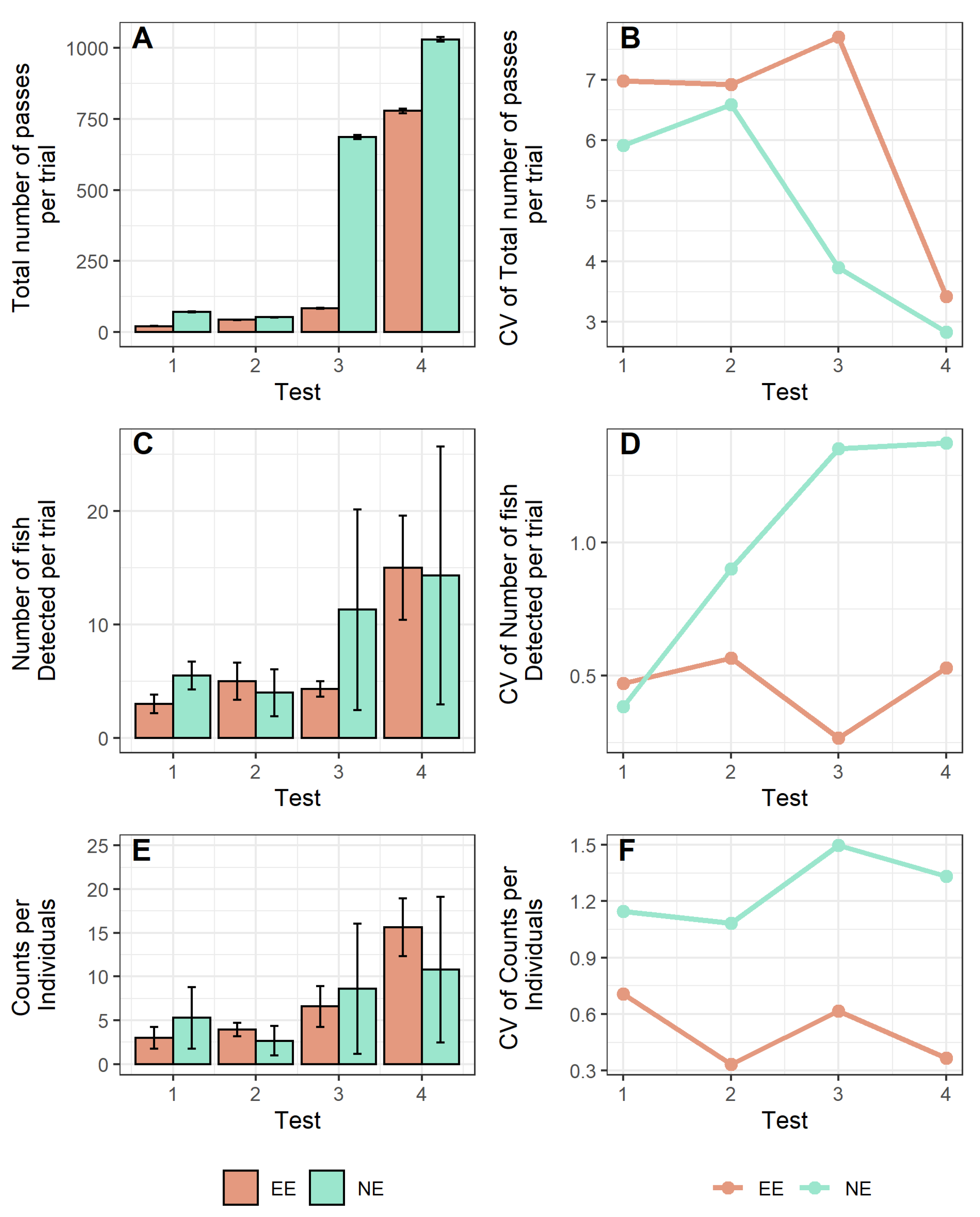
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergqvist, J.; Gunnarsson, S. Finfish Aquaculture: Animal Welfare, the Environment, and Ethical Implications. J. Agric. Environ. Ethic 2011, 26, 75–99. [Google Scholar] [CrossRef]
- Huntingford, F.; Kadri, S.; Jobling, M. Introduction: Aquaculture and Behaviour; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 1–35. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2021, 14, 704–728. [Google Scholar] [CrossRef]
- Brydges, N.; Braithwaite, V.A. Does environmental enrichment affect the behaviour of fish commonly used in laboratory work? Appl. Anim. Behav. Sci. 2009, 118, 137–143. [Google Scholar] [CrossRef]
- Jones, N.A.R.; Webster, M.M.; Salvanes, A.G.V. Physical enrichment research for captive fish: Time to focus on the DETAILS. J. Fish Biol. 2021, 99, 704–725. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2014, 17, 1–30. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Díaz-Gil, C.; Saraiva, J.L.; Moranta, D.; Castanheira, M.F.; Nuñez-Velázquez, S.; Ledesma-Corvi, S.; Mora-Ruiz, M.; Grau, A. Effects of structural environmental enrichment on welfare of juvenile seabream (Sparus aurata). Aquac. Rep. 2019, 15, 100224. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Caballero-Froilán, J.C.; Jiménez-García, M.; Capó, X.; Tejada, S.; Saraiva, J.; Sureda, A.; Moranta, D. Enriched environments enhance cognition, exploratory behaviour and brain physiological functions of Sparus aurata. Sci. Rep. 2020, 10, 11252. [Google Scholar] [CrossRef]
- Muñoz, L.; Aspillaga, E.; Palmer, M.; Saraiva, J.; Arechavala-Lopez, P. Acoustic Telemetry: A Tool to Monitor Fish Swimming Behavior in Sea-Cage Aquaculture. Front. Mar. Sci. 2020, 7, 645. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Lee, J.S.F.; Berejikian, B.A. Effects of the rearing environment on average behaviour and behavioural variation in steelhead. J. Fish Biol. 2008, 72, 1736–1749. [Google Scholar] [CrossRef]
- Øverli, Ø.; Sørensen, C.; Pulman, K.G.; Pottinger, T.; Korzan, W.; Summers, C.H.; Nilsson, G.E. Evolutionary background for stress-coping styles: Relationships between physiological, behavioral, and cognitive traits in non-mammalian vertebrates. Neurosci. Biobehav. Rev. 2007, 31, 396–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscolo, C.N.P.; Morais, R.N.; de Freitas, E.G. Same-sized fish groups increase aggressive interaction of sex-reversed males Nile tilapia GIFT strain. Appl. Anim. Behav. Sci. 2011, 135, 154–159. [Google Scholar] [CrossRef]
- Dara, M.; Dioguardi, M.; Vazzana, M.; Vazzana, I.; Accardi, D.; Carbonara, P.; Alfonso, S.; Cammarata, M. Effects of Social Hierarchy Establishment on Stress Response and Cell Phagocytosis in Gilt-Head Sea Bream (Sparus aurata). Fishes 2022, 7, 75. [Google Scholar] [CrossRef]
- Maguire, S.M.; DeAngelis, R.; Dijkstra, P.D.; Jordan, A.; Hofmann, H.A. Social network dynamics predict hormone levels and behavior in a highly social cichlid fish. Horm. Behav. 2021, 132, 104994. [Google Scholar] [CrossRef]
- Galhardo, L.; Almeida, O.; Oliveira, R.F. Preference for the presence of substrate in male cichlid fish: Effects of social dominance and context. Appl. Anim. Behav. Sci. 2009, 120, 224–230. [Google Scholar] [CrossRef]
- Castanheira, M.F.; Conceição, L.E.; Millot, S.; Rey, S.; Bégout, M.-L.; Damsgård, B.; Kristiansen, T.; Höglund, E.; Øverli, Ø.; Martins, C.I. Coping styles in farmed fish: Consequences for aquaculture. Rev. Aquac. 2015, 9, 23–41. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, S.; Millot, S.; Leguay, D.; Chatain, B.; Bégout, M.-L. Consistency in European seabass coping styles: A life-history approach. Appl. Anim. Behav. Sci. 2015, 167, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Huntingford, F.A.; Andrew, G.; Mackenzie, S.; Morera, D.; Coyle, S.M.; Pilarczyk, M.; Kadri, S. Coping strategies in a strongly schooling fish, the common carp Cyprinus carpio. J. Fish Biol. 2010, 76, 1576–1591. [Google Scholar] [CrossRef]
- Millot, S.; Bégout, M.-L.; Chatain, B. Risk-taking behaviour variation over time in sea bass Dicentrarchus labrax: Effects of day-night alternation, fish phenotypic characteristics and selection for growth. J. Fish Biol. 2009, 75, 1733–1749. [Google Scholar] [CrossRef] [Green Version]
- Leal, E.; Fernández-Durán, B.; Guillot, R.; Ríos, D.; Cerdá-Reverter, J.M. Stress-induced effects on feeding behavior and growth performance of the sea bass (Dicentrarchus labrax): A self-feeding approach. J. Comp. Physiol. B 2011, 181, 1035–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varsamos, S.; Flik, G.; Pepin, J.; Bonga, S.W.; Breuil, G. Husbandry stress during early life stages affects the stress response and health status of juvenile sea bass, Dicentrarchus labrax. Fish Shellfish. Immunol. 2006, 20, 83–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.F.; Herrera, M.; Costas, B.; Conceição, L.E.; Martins, C.I. Linking cortisol responsiveness and aggressive behaviour in gilthead seabream Sparus aurata: Indication of divergent coping styles. Appl. Anim. Behav. Sci. 2013, 143, 75–81. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Estimating regression models with unknown break-points. Stat. Med. 2003, 22, 3055–3071. [Google Scholar] [CrossRef]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: The MCMCglmm R Package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Schielzeth, H. Repeatability for gaussian and non-gaussian data: A practical guide for biologists. Biol. Rev. 2010, 85, 935–956. [Google Scholar] [CrossRef]
- Batzina, A.; Dalla, C.; Papadopoulou-Daifoti, Z.; Karakatsouli, N. Effects of environmental enrichment on growth, aggressive behaviour and brain monoamines of gilthead seabream Sparus aurata reared under different social conditions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 169, 25–32. [Google Scholar] [CrossRef]
- Batzina, A.; Dalla, C.; Tsopelakos, A.; Papadopoulou-Daifoti, Z.; Karakatsouli, N. Environmental enrichment induces changes in brain monoamine levels in gilthead seabream Sparus aurata. Physiol. Behav. 2014, 130, 85–90. [Google Scholar] [CrossRef]
- Batzina, A.; Kalogiannis, D.; Dalla, C.; Papadopoulou-Daifoti, Z.; Chadio, S.; Karakatsouli, N. Blue substrate modifies the time course of stress response in gilthead seabream Sparus aurata. Aquaculture 2014, 420, 247–253. [Google Scholar] [CrossRef]
- Batzina, A.; Karakatsouli, N. The presence of substrate as a means of environmental enrichment in intensively reared gilthead seabream Sparus aurata: Growth and behavioral effects. Aquaculture 2012, 370, 54–60. [Google Scholar] [CrossRef]
- Zhang, Z.; Fu, Y.; Zhang, Z.; Zhang, X.; Chen, S. A Comparative Study on Two Territorial Fishes: The Influence of Physical Enrichment on Aggressive Behavior. Animals 2021, 11, 1868. [Google Scholar] [CrossRef] [PubMed]
- Barreto, R.E.; Carvalho, G.G.A.; Volpato, G.L. The aggressive behavior of Nile tilapia introduced into novel environments with variation in enrichment. Zoology 2011, 114, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Torrezani, C.S.; Pinho-Neto, C.F.; Miyai, C.A.; Sanches, F.H.C.; Barreto, R.E. Structural enrichment reduces aggression in Tilapia rendalli. Mar. Freshw. Behav. Physiol. 2013, 46, 183–190. [Google Scholar] [CrossRef]
- Barley, A.J.; Coleman, R.M. Habitat structure directly affects aggression in convict cichlids Archocentrus nigrofasciatus. Curr. Zool. 2010, 56, 52–56. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Li, Z. Effects of different levels of environmental enrichment on the sheltering behaviors, brain development and cortisol levels of black rockfish Sebastes schlegelii. Appl. Anim. Behav. Sci. 2019, 218, 104825. [Google Scholar] [CrossRef]
- Roberts, L.; Taylor, J.; de Leaniz, C.G. Environmental enrichment reduces maladaptive risk-taking behavior in salmon reared for conservation. Biol. Conserv. 2011, 144, 1972–1979. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Posterior Mean | Lower 95% CI | Upper 95% CI | Effect. Samp. | pMCMC | Signif. | |
|---|---|---|---|---|---|---|
| Trial (time) | 1.233 | 0.906 | 1.543 | 622.19 | <2 × 10−4 | *** |
| Treatment (EE-NE) | 1.537 | −0.011 | 3.006 | 215.71 | 0.0477 | * |
| Trial × Treatment | −0.073 | −0.492 | 0.331 | 1597.92 | 0.7331 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arechavala-Lopez, P.; Nuñez-Velazquez, S.; Diaz-Gil, C.; Follana-Berná, G.; Saraiva, J.L. Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus labrax Juvenile Reared in Tanks. Fishes 2022, 7, 126. https://doi.org/10.3390/fishes7030126
Arechavala-Lopez P, Nuñez-Velazquez S, Diaz-Gil C, Follana-Berná G, Saraiva JL. Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus labrax Juvenile Reared in Tanks. Fishes. 2022; 7(3):126. https://doi.org/10.3390/fishes7030126
Chicago/Turabian StyleArechavala-Lopez, Pablo, Samira Nuñez-Velazquez, Carlos Diaz-Gil, Guillermo Follana-Berná, and João L. Saraiva. 2022. "Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus labrax Juvenile Reared in Tanks" Fishes 7, no. 3: 126. https://doi.org/10.3390/fishes7030126
APA StyleArechavala-Lopez, P., Nuñez-Velazquez, S., Diaz-Gil, C., Follana-Berná, G., & Saraiva, J. L. (2022). Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus labrax Juvenile Reared in Tanks. Fishes, 7(3), 126. https://doi.org/10.3390/fishes7030126