1. Introduction
Fishmeal (FM) was traditionally used as the main and most adequate protein source for carnivorous fish due to its high quality, high digestibility, and good palatability [
1,
2,
3]. Presently, its use is in a clear downward trend [
4]. This reduction is largely due to supply and price variation, coupled with the continuously increasing demand from the aquafeed industry [
4]. Hence, the use of plant feedstuffs (PF) and the inclusion of carbohydrate (CH) sources in fish feeds have been good alternatives to, respectively, decreasing dietary FM inclusion as a protein source and spare protein use for growth [
5,
6,
7,
8,
9,
10]. Gilthead seabream (
Sparus aurata), one of the species with higher production in Europe, seems able to cope with a total replacement of dietary FM by PF [
9]. This species requires about 45% of dietary protein [
11]. However, if digestible CHs are provided in a suitable quantity, dietary protein might be spared for growth instead of being used as an energy source and, therefore, reduce nitrogen wastes and dietary costs [
6,
7,
12]. Nonetheless, the maximum dietary CH inclusion that does not cause negative effects in gilthead seabream is limited to 20% [
7]. Higher dietary CH inclusion may compromise growth and the digestive and absorptive capacities [
6,
12]. Several studies with gilthead seabream were already conducted to separately evaluate the effects of dietary inclusion of PF and the protein-sparing by CHs. Overall, results showed that PF-based diets often promoted gut morphological changes, modifications on microbiota composition, decreases in gut enzymatic activity, and increases in oxidative stress of fish [
9,
13,
14,
15,
16,
17,
18,
19,
20,
21]. The inclusion of 20% or more of dietary CHs also affected fish growth performance, digestive enzyme activities, and antioxidant status [
7,
22,
23,
24]. However, the interactive effects on gut functionality and the health of gilthead seabream fed diets with lower P/CH ratios and the replacement of FM by PF as a major dietary protein source has not received much attention, and the available information is somehow dispersed. For instance, Castro et al. [
25] did not observe major changes in gut histomorphology, microbiota, α-amylase, and lipase activities of gilthead seabream fed diets with highly different P/CH ratios (50/17 and 66/0). Similarly, in the same species, Couto et al. [
26] and Fountoulaki et al. [
27] also did not find an effect of the dietary P/CH ratio on the proteolytic and amylolytic activities, nor did Castro et al. [
23] on the gut oxidative status, or antioxidant enzymes activities. All these studies evaluating different dietary P/CH ratios were made with FM as the main dietary protein source. To our knowledge, only one study is available that evaluated dietary P/CH ratios using PF as the main protein source [
12]. In this study, the authors reported that fish fed a P40/CH39 diet had higher lipase and trypsin activities and lower α-amylase activity than those fed a P46/CH19 diet.
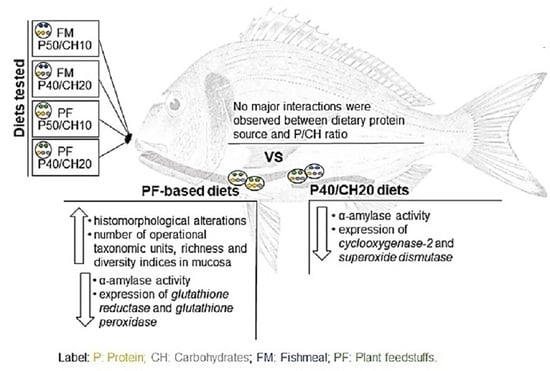
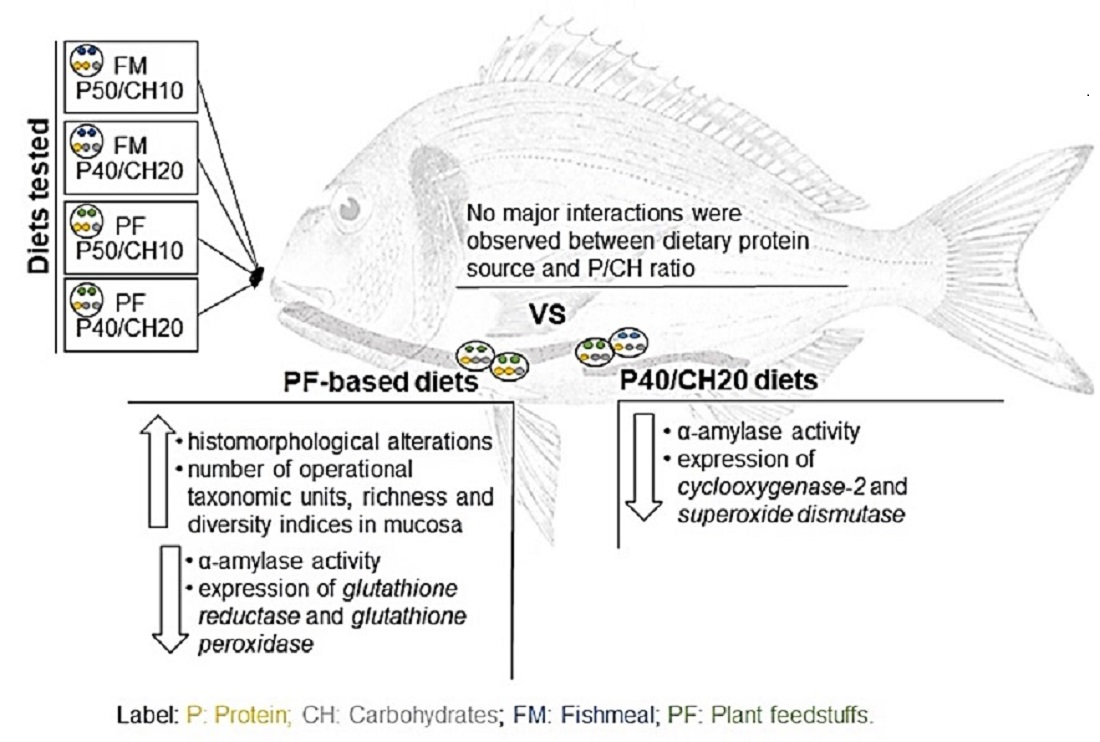
Recently, we assessed the effects of FM- or PF-based diets with different P/CH ratios (50/10 and 40/20) in gilthead seabream growth, feed utilization, appetite regulation, and intermediary metabolism [
28]. Results showed that diets only slightly modified fish appetite and metabolic parameters, although growth was higher in fish fed the FM-P50/CH10 diet than those fed the FM-P40/CH20 diet. Further, reducing the dietary P/CH ratio led to a decrease in the feed efficiency and an increase in the protein efficiency ratio.
The present study is a follow-up to our previous study [
28]. While the previous study aimed to evaluate the effect of dietary protein sources (FM vs. PF) and P/CH ratio on gilthead seabream appetite regulation and intermediary metabolism, the present study aims to evaluate, for the first time, the effects of these factors (dietary protein source and P/CH ratio) on gilthead seabream gut function and health, by assessing gut histomorphology, gut microbiota composition, digestive enzymes activity, and gut immunological and oxidative stress genes expression.
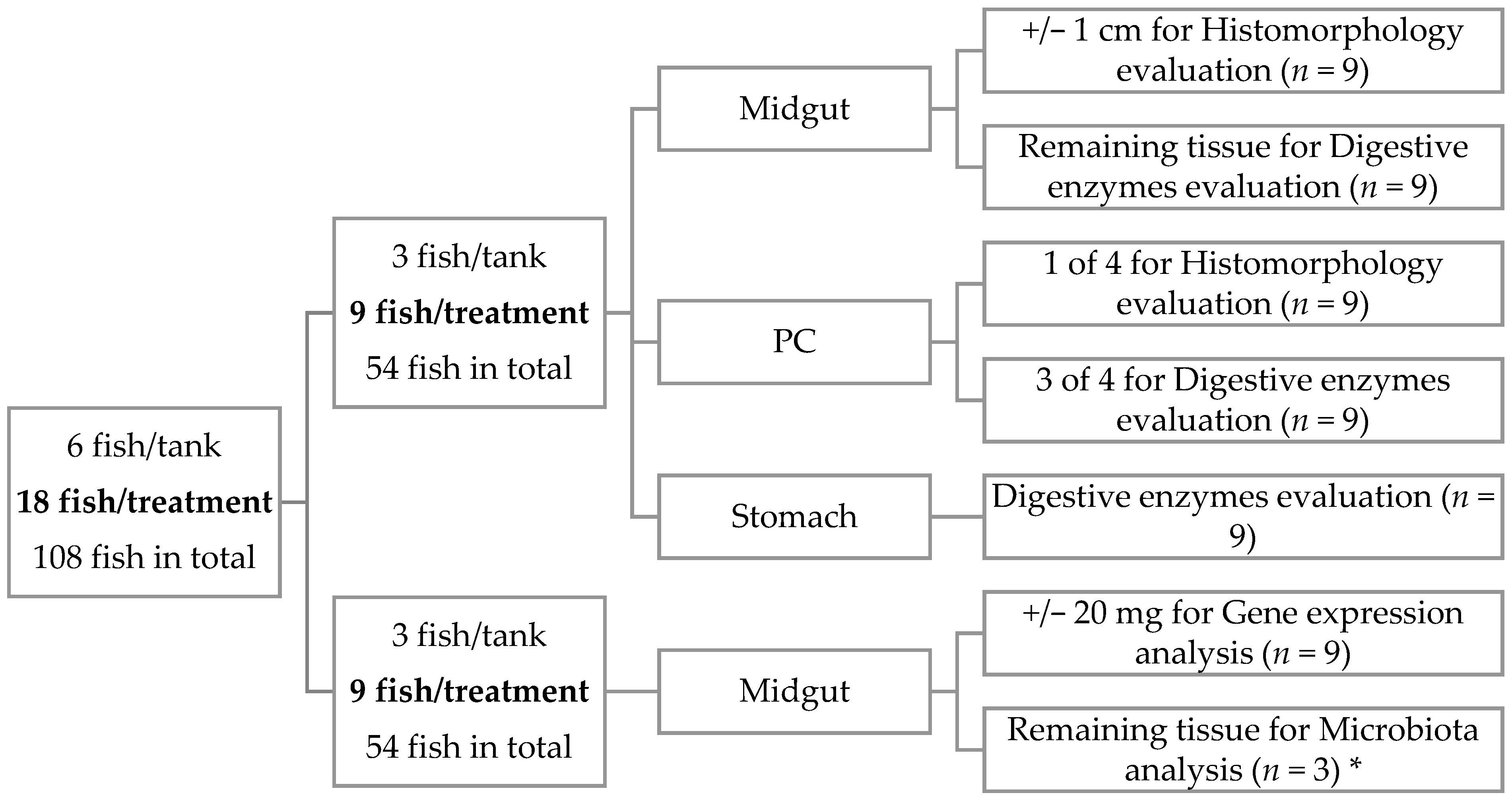
3. Results
During the trial, all experimental diets were well-accepted by the fish, and the fish survival rate was 100%. Results of the rearing trial were not the aim of this study and are presented elsewhere [
28].
Regarding the PC histomorphology, fish fed the PF-P50/CH10 diet presented a higher total mean score (2.23) than those in the remaining experimental conditions, where the total mean score ranged between 1.78 and 1.88 (
Table 2). Lamina propria width was higher in fish fed the PF-P50/CH10 diet than in those fed the FM-based diets (
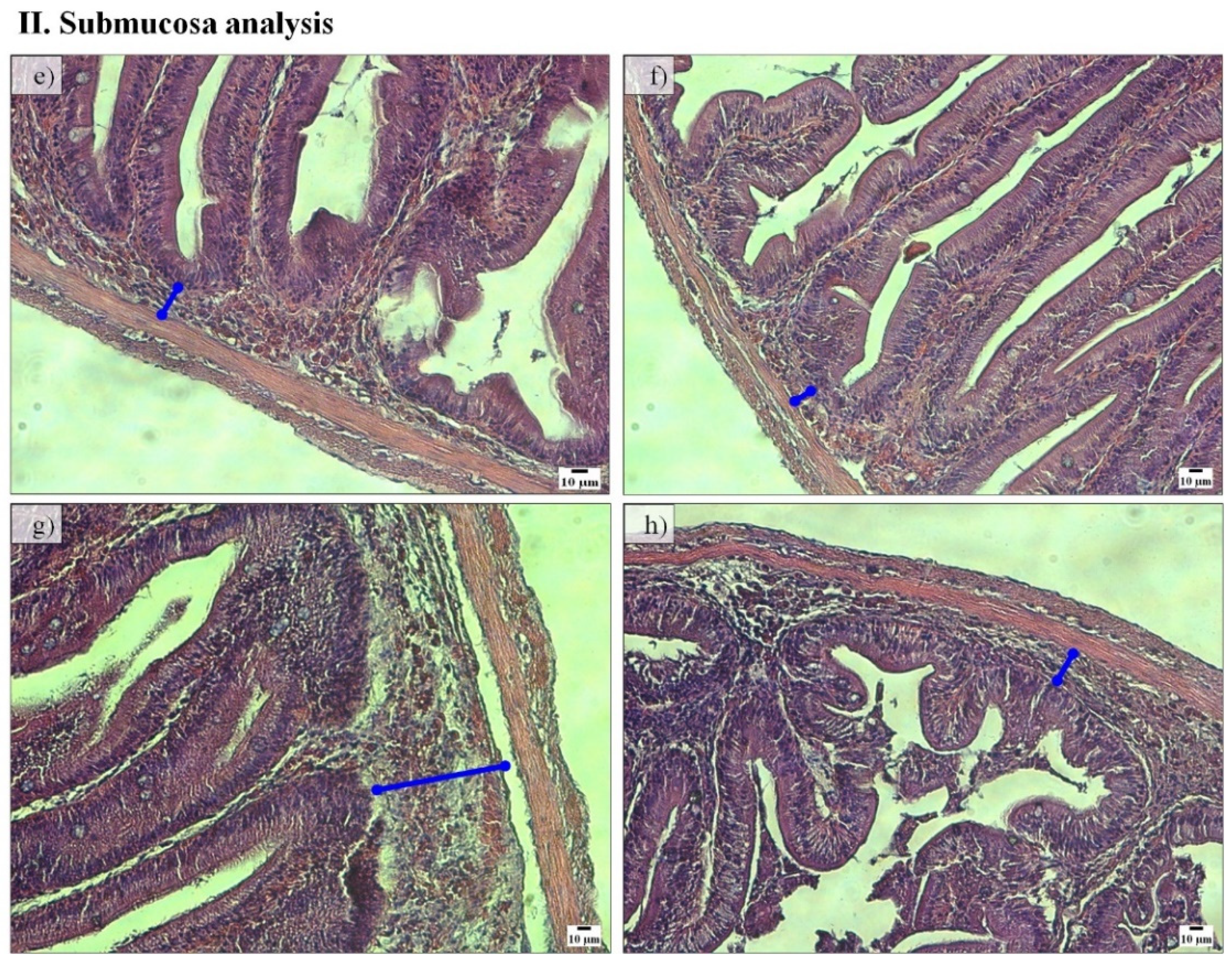
Figure 2I). Fish fed the PF-P50/CH10 diet also presented higher submucosa widths than those remaining in the experimental conditions (
Figure 2II). Lamina propria cellularity was higher in fish fed the FM-P50/CH10 and PF-P50/CH10 diets than the FM-P40/CH20 diet. The enterocytes vacuolization was higher in fish fed the PF-based diets.
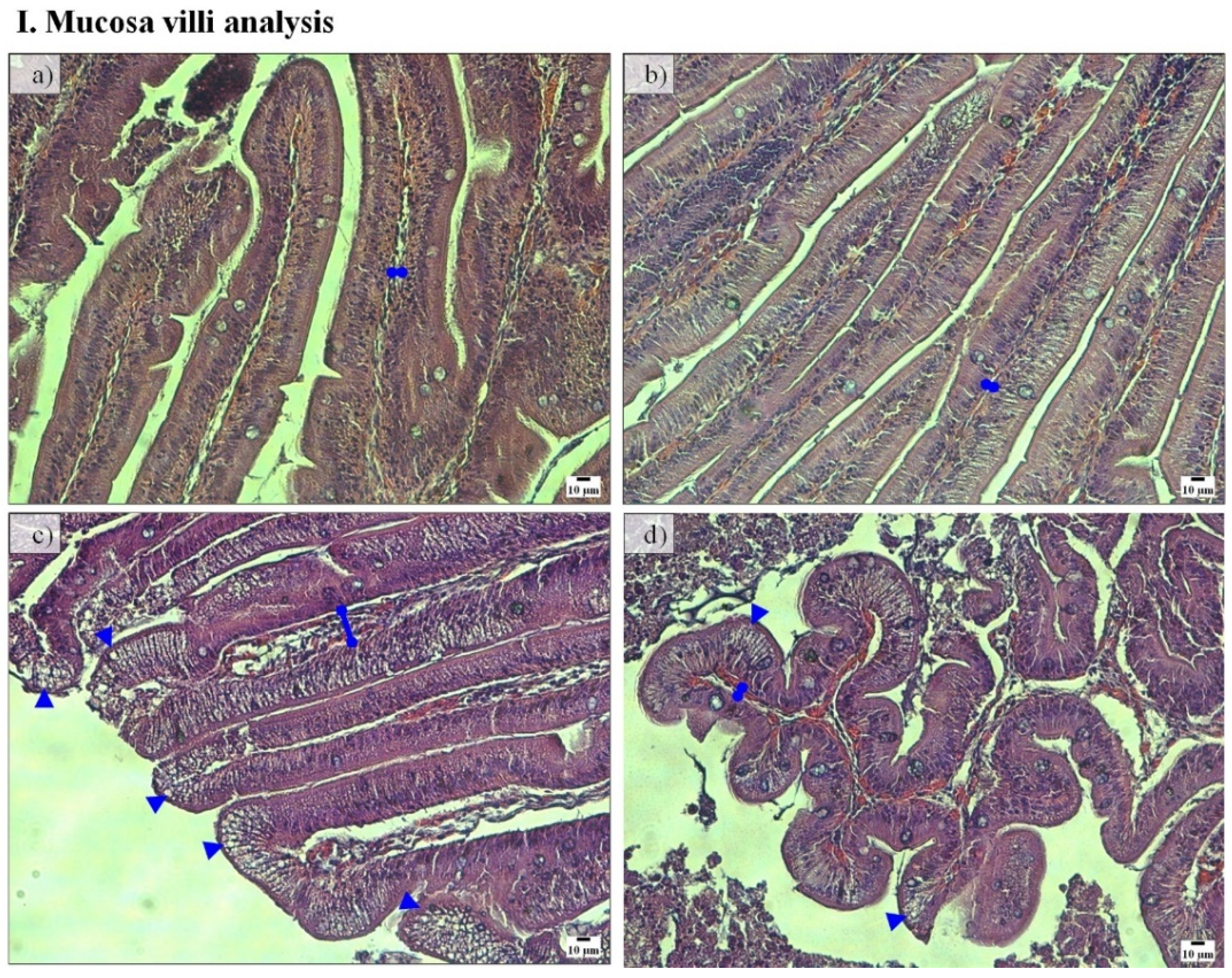
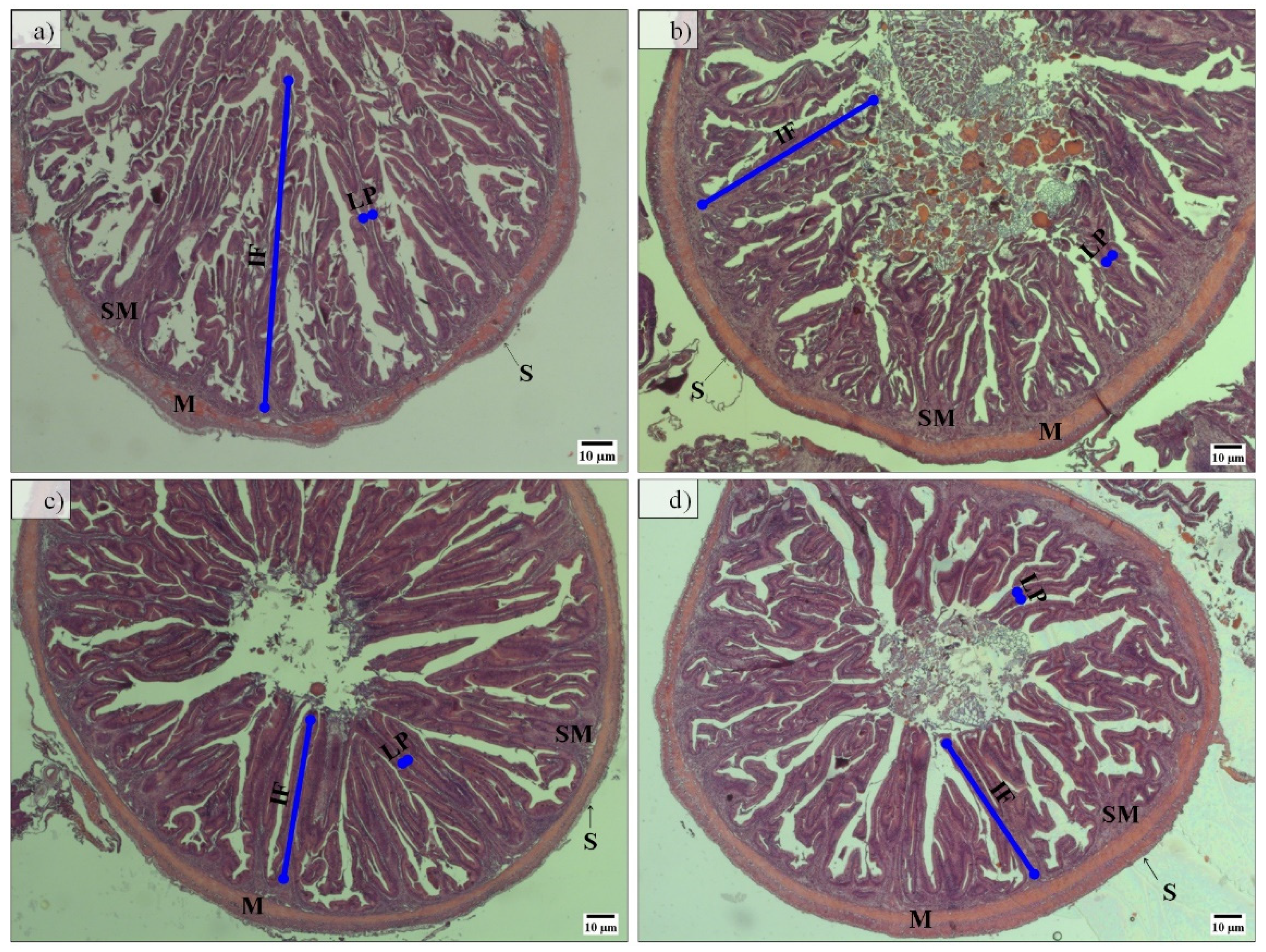
Regarding midgut histomorphology, fish fed the PF-based diets presented a higher total mean score (2.77) and gut fold height than fish fed FM-based diets, which have a total mean score of 2.28 (
Figure 3 and
Table 3). No further differences between groups were detected.
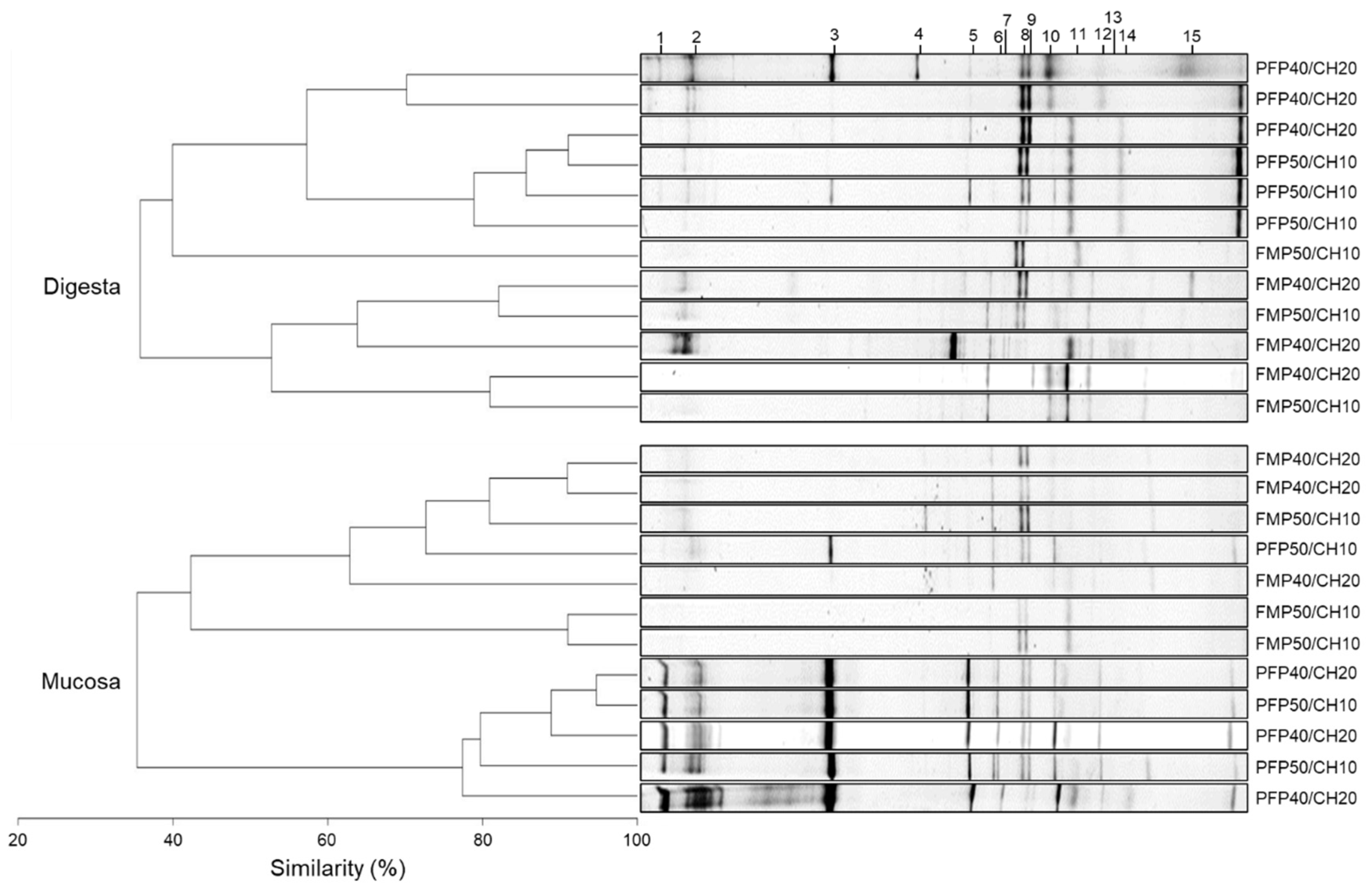
DGGE fingerprints of the hypervariable V3 region of the 16S rRNA genes present in digesta and mucosa gut samples revealed that, independently of the dietary treatment, gut bacterial communities maintained a similarity, near 40% within both gut samples (
Figure 4). Moreover, two clusters were observed in both gut microbiota regions, corresponding to samples recovered from fish fed the FM- and the PF-based diets, except for the FM-P50/CH10 diet in the digesta, which did not cluster with the remaining FM-based diets, and the PF-P50/CH10 diet in the mucosa, which did not cluster with the remaining PF-based diets. Despite this clear cluster separation, in digesta samples, the dietary composition did not affect the average number of OTUs, richness, and diversity indices (
Table 4). Only the similarity index was higher in fish fed PF-P50/CH10 than in fish fed the FM-P50/CH10 diet. In mucosa samples, PF-based diets led to a higher number of gut OTUs, richness, and diversity indices than FM-based diets, while the similarity index was not different between groups. Sequence analysis from DGGE-selected bands showed that the dominant allochthonous and autochthonous bacteria detected were either corresponding to uncultured bacteria not yet assigned to a specific taxon or were closely related to genera belonging to the phylum Firmicutes and Proteobacteria, namely, Lactobacillus, Pseudomonas, Klebsiella, and Vibrio (
Table 5 and
Figure 4). Except for band 15, which was only found in digesta, all other bands were detected in digesta and mucosa samples.
Concerning digestive enzymes, α-amylase activity was lower in fish fed the PF-based diets, for both PC and midgut, and in fish fed the P40/CH20 diet only in the PC (
Table 6). Proteolytic activity was higher in the PC of fish fed the P50/CH10 diet, but only within the PF-based diet-fed fish. Pepsin and lipase activities were not affected by dietary composition.
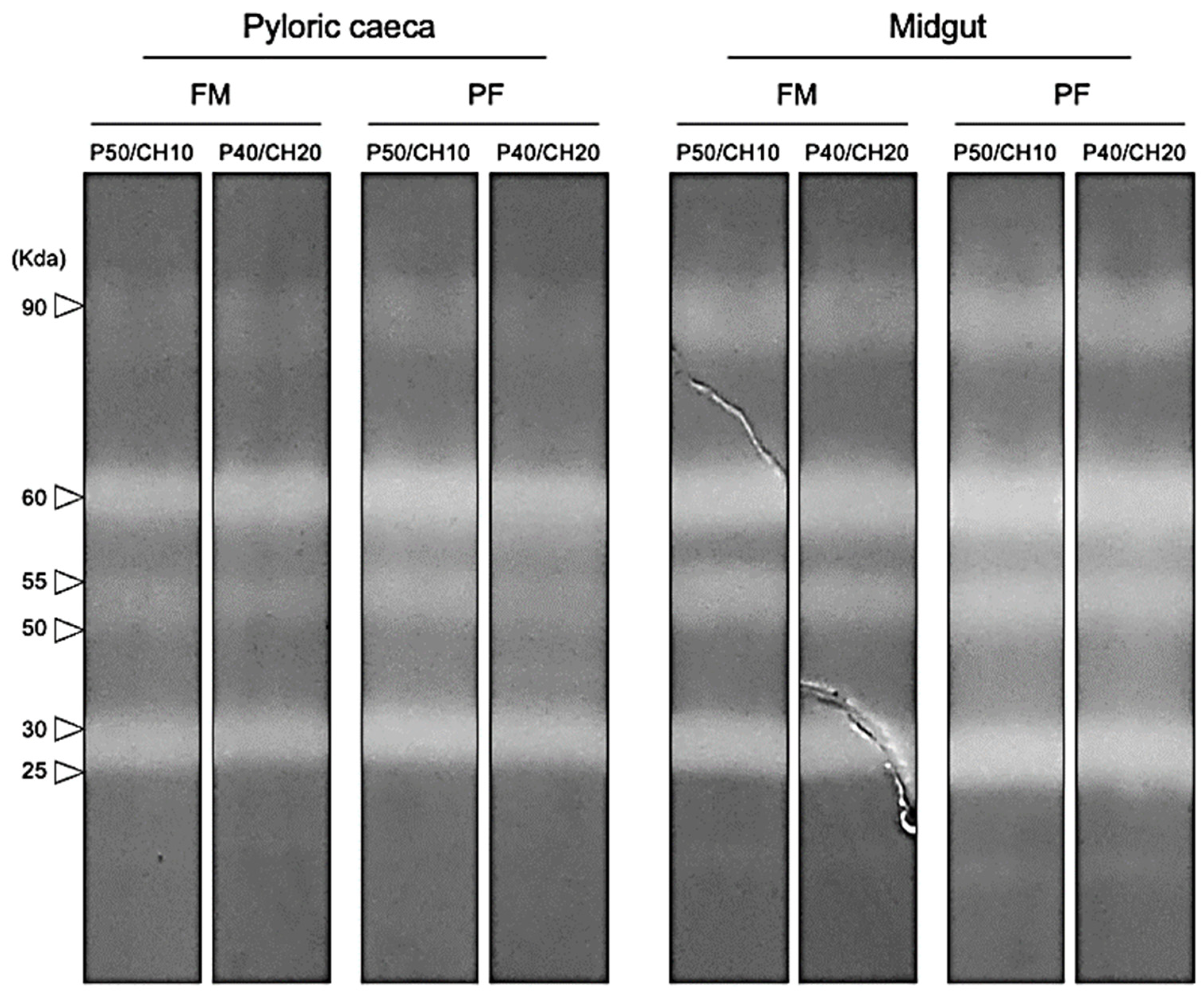
Alkaline protease zymograms, from both PC and midgut, revealed the presence of six bands with proteolytic activity against casein, three identified as trypsin-like proteases (90, 60, and 55 KDa), and the other three as chymotrypsin-like proteases (50, 30, and 25 KDa). All treatments presented the same number of proteolytic bands (
Figure 5).
Concerning immune-related gene expressions, only
cyclooxygenase-2 (
cox2) presented significant changes, being higher in fish fed the P50/CH10 diet (
Table 7). Gene expression of
immunoglobulin M heavy chain (
igM),
interleukin-1β (
il1β), and
tumor necrosis factor-α (
tnf-α) was not affected by dietary composition.
Regarding the oxidative stress-related genes, PF-based diets led to a lower expression of glutathione reductase (gr) and glutathione peroxidase (gpx1), while superoxide dismutase (sod) expression was lower in fish fed the P40/CH20 diet. The gene expression of 70 kilodalton heat shock proteins (hsp70) and catalase (cat) was not affected by dietary composition.
4. Discussion
The presence of antinutritional factors on PF, namely, in soybean products, was reported as leading to gut inflammation in gilthead seabream [
15,
18,
20,
21,
39]. Among the observed gut morphological alterations caused by soybean meal were a decrease in gut fold height, an enlargement of submucosa and lamina propria, an increased number of inflammatory cells on tissues, and modifications on enterocytes vacuolization [
15,
18,
20,
21]. Although we have assessed the midgut and PC, and previous studies analyzed the distal gut, the present results agree with the reported observations in this species, since fish fed the PF-P50/CH10 diet, which has a higher soybean meal content (25% compared with 19% for PF-P40/CH20, and no soybean meal content for FM-based diets), also presented more histological alterations when compared with fish fed the other diets. The histological modifications observed in the midgut and PC were mainly in gut fold height, width and cellularity of lamina propria, width of the submucosa, and/or in enterocytes vacuolization. Similarly, gilthead seabream juveniles fed 30% soybean meal presented a moderately and diffusely expanded distal gut lamina propria [
14], while juveniles fed soy saponins and phytosterols presented histomorphological alterations of the intestinal mucosal structure [
17]. Nonetheless, during the on-growing period (fish of similar sizes to those of the present study) gilthead seabream showed a high tolerance to soy saponins and phytosterols [
29]. This indicates that fish responses can be different, depending on the life stage, dietary ingredients/antinutrients combinations, and intestine portions.
Moreover, in the present study, PC seemed to be more sensitive to dietary composition changes than midgut, where fewer histomorphological alterations were observed. This agreed with the study of Couto et al. [
29], which observed that dietary soy saponins and phytosterols affected PC histomorphology but not the distal gut of on-growing gilthead seabream.
However, it is important to add that the observed histomorphology modifications were not enough to consistently affect gilthead seabream growth [
28]. Nonetheless, a longer experimental trial could have exposed those differences.
The composition of gut microbiota also affects gut functionality since, for instance, bacteria might have a role in nutrients’ digestion and immune functions, being affected by diet composition [
39]. In the present study, protein source was the single factor affecting gut microbiota. The only detectable effect on digesta microbiota was an increase of the similarity index in fish fed the PF-P50/CH10 diet, indicating that this diet might modulate gut bacteria populations towards a higher similarity between samples. The absence of any other major effect on digesta microbiota in fish fed different dietary compositions was previously observed in gilthead seabream [
25]. This lack of effect could be expected, since digesta microbiota comprises transient (allochthonous) microorganisms, which are often surrounded by the resident microbiota to the gut wall and, thus, do not last a long time in the gut [
40].
The higher number of OTUs, richness, and diversity indices observed in the mucosa microbiota of fish fed the PF-based diets agree with what was previously reported for this species, at the juvenile stage, fed soybean meal-based diets compared with FM-based diets [
16], and for other species also fed PF-based diets, such as Senegalese sole (
Solea senegalensis) and Atlantic salmon [
41,
42,
43]. These results could be explained by the presence of non-digestible carbohydrates on PF, which provide the required substrate for gut bacteria proliferation [
44,
45]. It should be noticed that higher richness and diversity indices, as in fish fed the PF-based diets, can be undesirable since they can be associated with the presence of pathogenic bacteria in gut microbiota [
18,
46]. On the other hand, a diverse gut microbiome, with the increase of microorganisms from the Firmicutes phylum, can stimulate a fish’s innate immunity and reduce the gut surface area for the establishment of pathogenic bacteria, improving the fish’s health [
47,
48,
49]. Although, in the present study, none of the immune-related genes measured were affected by the use of PF, the dominant allochthonous and autochthonous bacteria detected were indeed the most closely related to the Firmicutes and Proteobacteria phyla, as already described in gilthead seabream fed different dietary compositions [
18,
47]. However, in future studies, a higher-resolution method, such as next-generation sequencing and FISH, could improve the characterization of the bacterial communities under different dietary feeding regimes, providing not only the full identification of the species and/or subspecies of the bacteria, but also allowing for their quantification. This more in-depth characterization and quantification of the bacterial species and/or subspecies will possibly allow for a clearer connection between microbiota and gut functionality.
Both
Pseudomonas sp. and
Lactobacillus sp. can produce α-amylase [
50]; however, as their presence was detected in fish fed all experimental diets, no link can be made between the presence of α-amylase-producing bacteria, the dietary ratios, and α-amylase activity measured. Indeed, the lack of differences in the gut microbiota of fish fed different dietary P/CH ratios could be partially explained by the use of pregelatinized maize starch as the main carbohydrate source. Gilthead seabream presents almost 100% starch digestibility of diets including 10 to 30% of this ingredient [
26]; thus, pregelatinized maize starch does not seem to provide a substantial substrate for microbial fermentation and development. A similar lack of changes in gut microbiota was reported for gilthead seabream and other fish species fed also with highly digestible starch [
26,
51,
52].
For diets’ digestion, several enzymes are needed, with each enzyme presenting a specific role. α-amylase, proteases, and lipase are, respectively, responsible for the enzymatic hydrolysis of starch, proteins, and lipids [
51,
52,
53]. Despite that we did not observe any major effect on the feed intake of fish fed the different diets [
28], in the present study, α-amylase activity in PC and midgut and proteolytic activity in PC were affected by the dietary composition. The α-amylase activity was lower in the PC and midgut of fish fed the P40/CH20 diet, and in the PC, it was also lower in fish fed the PF-based diets than those fed the FM-based diets. The influence of dietary P/CH ratio can be related to the adsorption of α-amylase by starch, as suggested by Spannhof and Plantikow [
54], who observed that α-amylase secreted by fish during the digestive process was adsorbed by the starch present in the diets [
55,
56]. This lower α-amylase activity observed in fish fed the P40/CH20 diet can partially explain the lower feed efficiency observed in our previous study in fish fed the P40/CH20 diet, in comparison with those fed the P50/CH10 diet [
28]. The effects of dietary protein sources may be related to the ingredients used, namely, wheat gluten, which is a source of α-amylase inhibitors [
55,
56].
According to Hidalgo et al. [
57] and Fernández et al. [
58], α-amylase activity is more dependent on fish nutritional habits than the proteolytic activity, and this is further supported by the lack of effects on the proteolytic activity reported in gilthead seabream fed diets with different P/CH ratios [
25,
26,
27]. However, studies in other fish species showed that higher dietary protein levels increased proteolytic activity [
59,
60,
61,
62]. In the present study, higher proteolytic activity in fish fed the diet with a higher protein content was also observed in the PC, but only in fish fed the PF-based diets. Moreover, no differences were found regarding the alkaline protease pattern, as observed in the zymograms of the different dietary treatments, suggesting the proteases present are the same independently of the diet offered. Differently, García-Meilán et al. [
24] observed that, in gilthead seabream fed FM-based diets, PC proteolytic activity was higher in fish fed lower dietary protein-content diets (P35 and P38), while in the midgut, the proteolytic activity increased progressively as dietary protein increased, stabilizing at 41% to 47% of protein. Thus, more studies should be conducted to clarify the effects of dietary protein level and source on proteolytic activity in the gut.
In the present study, fish fed the PF-based diets presented lower
gr and
gpx1 gene expression than those fed FM-based diets, which may indicate that the former were more vulnerable to oxidative stress [
63]. This evidence seems to be in agreement with the presence of soybean meal antinutritional factors, such as the β-conglycinin, which has been identified as one of the major feed allergens [
64,
65]. This allergen has an N-glycan structure, essential for the formation of di-tyrosine bridges, which trigger the process responsible for oxidative stress, increasing the malondialdehyde content, and causing oxidative damages [
64].
Regarding dietary P/CH ratio effects on oxidative stress, the decrease of
sod gene expression in fish fed the P40/CH20 diet may indicate that those fish were also more susceptible to oxidative stress. Nevertheless, Castro et al. [
23] observed in gilthead seabream that the intestinal sod activity was not affected by the use of different dietary P/CH ratios. Indeed, a disconnection between the gene expression and enzymatic activity results was previously reported by other studies [
22,
66]. Thus, we might not disregard that the response at the biochemical level might be different of the one obtained at molecular level. Hence, future studies should also include enzymatic activities which, together with the gene expression analyses, will allow for a more complete conclusion.
Sitjà-Bobadilla et al. [
13] and Kokou et al. [
19,
20] reported, in gilthead seabream fed PF-based diets, a synchronism between the immune and stress responses and the gut histomorphological alterations. A similar relationship was observed in the present study, although no effects were observed in the immune-related genes analyzed, except for
cox2 expression, which was higher in fish fed the high-protein diets. Cox2 is linked mainly to inflammation [
67,
68], so it might be expected that an increase of
cox2 gene expression would be accompanied by higher histomorphological scores in this group, which did not happen. The absence of effects on immune-related responses seems to agree with the lack of mortality or diseases observed in our previous study [
28].


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}