
Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.2. Diets
2.3. Experimental Design and Sampling Procedure
2.4. Growth Performance and Biometric Parameters
2.5. Metabolites in Plasma and Tissues
2.6. Hormones
2.7. Statistical Analyses
3. Results
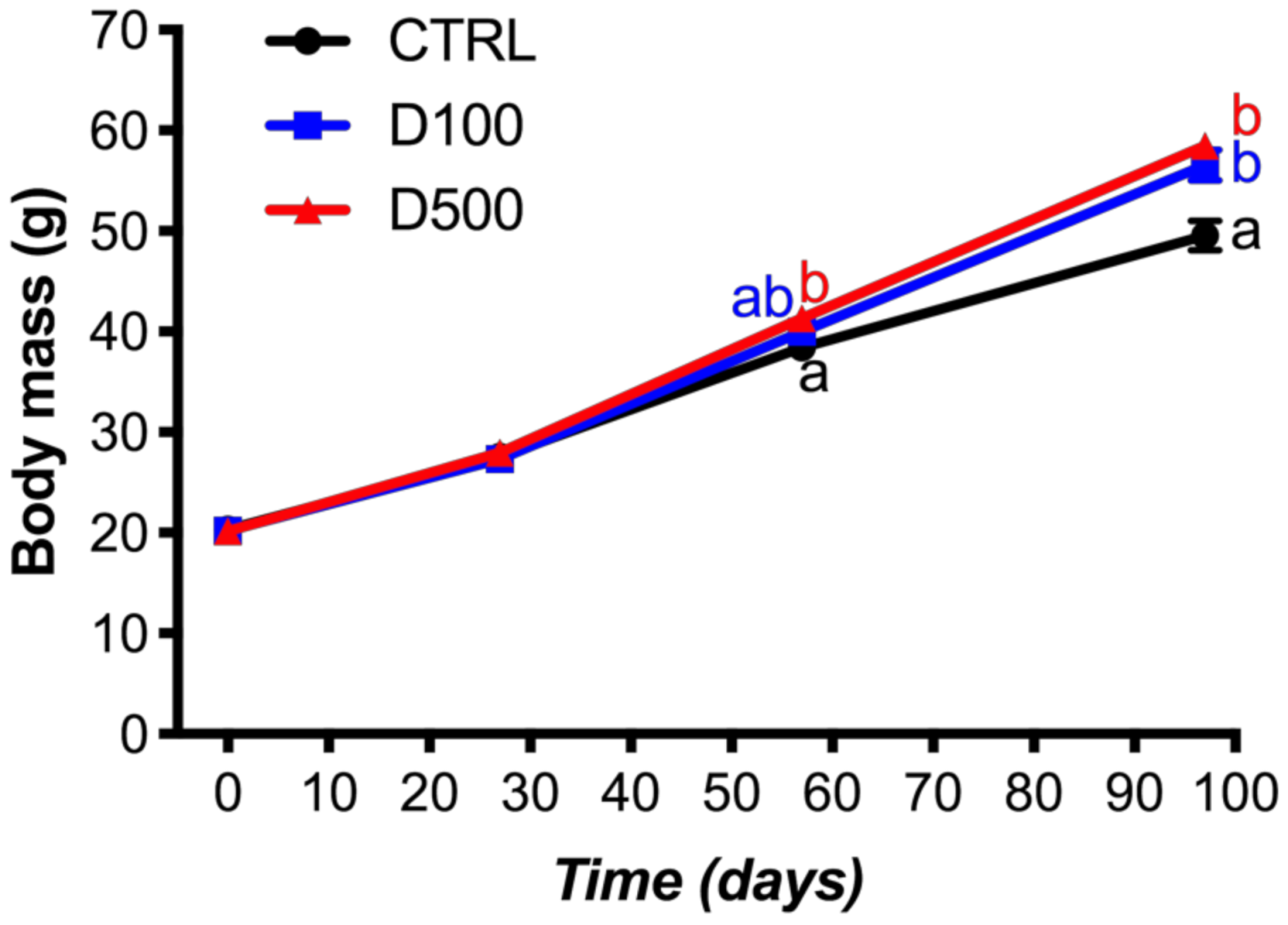
3.1. Growth Performance and Biometric Parameters

3.2. Metabolites in Plasma and Tissues
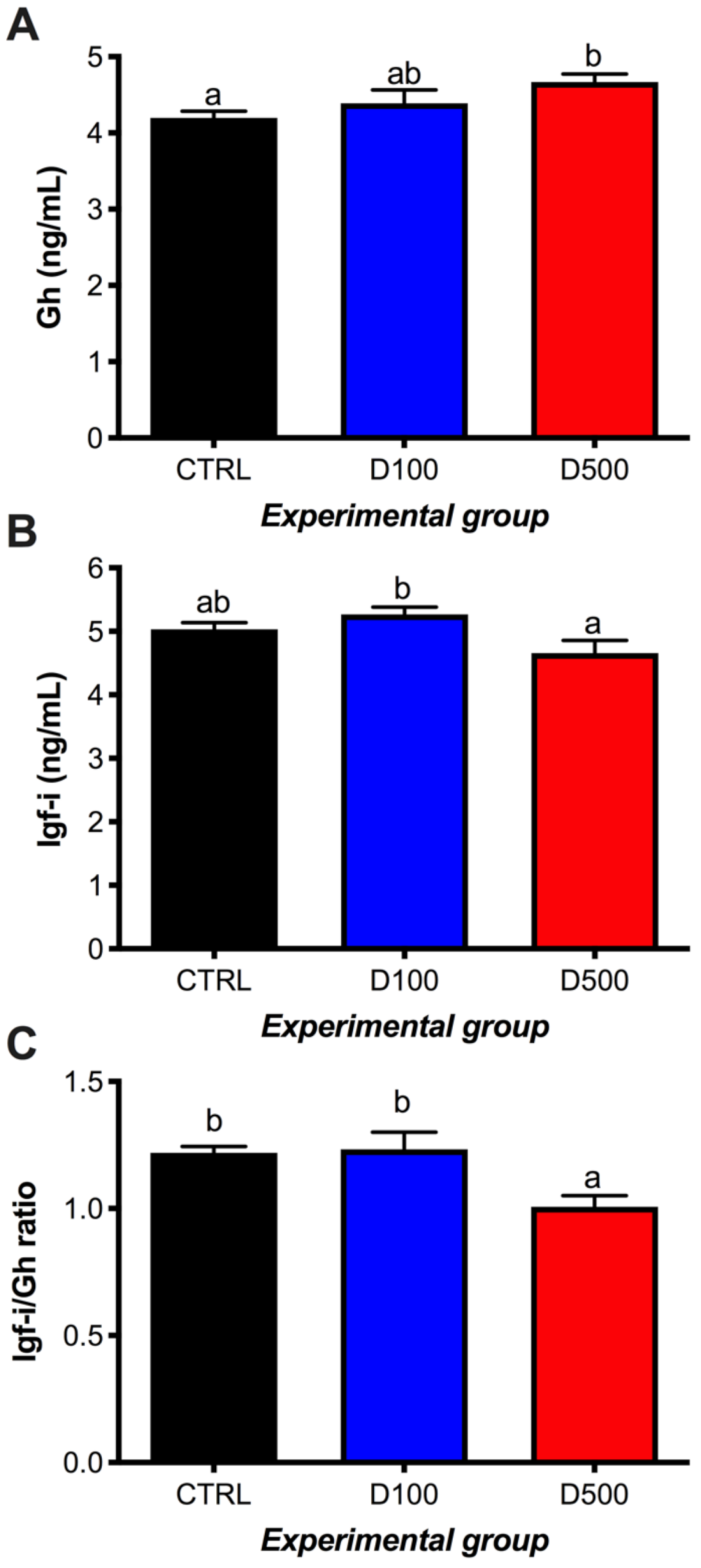
3.3. GH, IGF-I, and Cortisol in Plasma
4. Discussion
5. Conclusions and Open Issues
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Meeting the sustainable development goals. In The State of World Fisheries and Aquaculture 2018; FAO: Rome, Italy, 2018; Available online: https://www.fao.org/documents/card/es/c/I9540EN (accessed on 26 January 2022).
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Rajeswari, J.J.; Velasco, C.; Unniappan, S. Nutrient Regulation of Endocrine Factors Influencing Feeding and Growth in Fish. Front. Endocrinol. 2019, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Vélez, E.J.; Unniappan, S. A Comparative Update on the Neuroendocrine Regulation of Growth Hormone in Vertebrates. Front. Endocrinol. 2021, 11, 1174. [Google Scholar] [CrossRef] [PubMed]
- Estensoro, I.; Ballester-Lozano, G.; Benedito-Palos, L.; Grammes, F.; Martos-Sitcha, J.A.; Mydland, L.T.; Calduch-Giner, J.A.; Fuentes, J.; Karalazos, V.; Ortiz, A.; et al. Dietary butyrate helps to restore the intestinal status of a marine teleost (Sparus aurata) fed extreme diets low in fish meal and fish oil. PLoS ONE 2016, 11, e0166564. [Google Scholar] [CrossRef]
- Martos-Sitcha, J.A.; Simó-Mirabet, P.; Piazzon, M.C.; de las Heras, V.; Calduch-Giner, J.A.; Puyalto, M.; Tinsley, J.; Makol, A.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Dietary sodium heptanoate helps to improve feed efficiency, growth hormone status and swimming performance in gilthead sea bream (Sparus aurata). Aquac. Nutr. 2018, 24, 1638–1651. [Google Scholar] [CrossRef]
- Perera, E.; Sánchez-Ruiz, D.; Sáez, M.I.; Galafat, A.; Barany, A.; Fernández-Castro, M.; Antonio Jesús Vizcaíno, A.J.; Fuentes, J.; Martínez, T.F.; Mancera, J.M.; et al. Low dietary inclusion of nutraceuticals from microalgae improves feed efficiency and modifies intermediary metabolisms in gilthead sea bream (Sparus aurata). Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Khatib, N.; Gaidhane, S.; Gaidhane, A.M.; Khatib, M.; Simkhada, P.; Gode, D.; Zahiruddin, Q.S. Ghrelin: Ghrelin as a regulatory Peptide in growth hormone secretion. J. Clin. Diagn Res. 2014, 8, MC13–MC17. [Google Scholar] [CrossRef]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Kaiya, H.; Kangawa, K.; Miyazato, M. What is the general action of ghrelin for vertebrates? Comparisons of ghrelin’s effects across vertebrates. Gen. Comp. Endocrinol. 2013, 181, 187–191. [Google Scholar] [CrossRef]
- Riley, L.G.; Hirano, T.; Grau, E.G. Rat ghrelin stimulates growth hormone and prolactin release in the tilapia, O. mossambicus. Zool. Sci. 2002, 19, 797–800. [Google Scholar] [CrossRef]
- Kaiya, H.; Miyazato, M.; Kangawa, K.; Peter, R.E.; Unniappan, S. Ghrelin: Amultifunctional hormone in non-mammalian vertebrates. Comp. Biochem. Physiol. A 2008, 149, 109–128. [Google Scholar] [CrossRef]
- Kaiya, H.; Miyazato, M.; Kangawa, K. Recent advances in the phylogenetic study of ghrelin. Peptides 2011, 32, 2155–2174. [Google Scholar] [CrossRef]
- Jönsson, E. The role of ghrelin in energy balance regulation in fish. Gen. Comp. Endocrinol 2013, 187, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Perelló-Amorós, M.; Vélez, E.J.; Vela-Albesa, J.; Sánchez-Moya, A.; Riera-Heredia, N.; Hedén, I.; Fernández-Borràs, J.; Blasco, J.; Calduch-Giner, J.A.; Navarro, I.; et al. Ghrelin and Its Receptors in Gilthead Sea Bream: Nutritional Regulation. Front. Endocrinol. 2018, 9, 399. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Guillén, C.; Dias, J.; Rocha, F.; Castanheira, M.F.; Martins, C.I.; Laizé, V.; Gavaiaa, P.J.; Engrola, S. Does a ghrelin stimulus during zebrafish embryonic stage modulate its performance on the long-term? Comp. Biochem. Physiol. A 2019, 228, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Guillén, C.; Yufera, M.; Engrola, S. Ghrelin in Senegalese sole (Solea senegalensis) post-larvae: Paracrine effects on food intake. Comp. Biochem. Physiol. A 2017, 204, 85–92. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Bernardini, G.; Gornati, R.; Saroglia, M. Sea bass ghrelin: Molecular cloning and mRNA quantification during fasting and refeeding. Gen. Comp. Endocrinol. 2008, 155, 341–351. [Google Scholar] [CrossRef]
- Momany, F.A.; Bowers, C.Y.; Reynolds, G.A.; Chang, D.; Hong, A.; Newlander, K. Design, synthesis and biological activity of peptides which release growth hormone, in vitro. Endocrinology 1981, 108, 31–39. [Google Scholar] [CrossRef]
- Camanni, F.; Ghigo, E.; Arvat, E. Growth Hormone-Releasing Peptides and Their Analogs. Front. Neuroendocrinol. 1998, 19, 47–72. [Google Scholar] [CrossRef]
- Hashizume, T.; Sasaki, K.; Sakai, M.; Tauchi, S.; Masuda, H. The effect of new growth hormone-releasing peptide (KP 102) on the release of growth hormone in goats. Anim. Sci. Technol. 1997, 68, 247–256. [Google Scholar]
- Roh, S.G.; Lee, H.G.; Phung, L.T.; Hidari, H. Characterization of Growth Hormone Secretion to Growth Hormone releasing Peptide-2 in Domestic Animals-A Review. Asian-Australas J. Anim. Sci. 2002, 15, 757–766. [Google Scholar] [CrossRef]
- Chan, C.B.; Fung, C.K.; Fung, W.; Margaret, C.L.; Cheng, C.H. Stimulation of growth hormone secretion from seabream pituitary cells in primary culture by growth hormone secretagogues is independent of growth hormone transcription. Comp. Biochem. Physiol. C 2004, 139, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Bowers, C.Y.; Momany, F.A.; Reynolds, G.A.; Hong, A. On the in vitro and in vivo activity of a new synthetic hexapeptide that acts on the pituitary to specifically release growth hormone. Endocrinology 1984, 114, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Yahashia, S.; Kang, K.S.; Kaiya, H.; Matsudaa, K. GHRP-6 mimics ghrelin-induced stimulation of food intake and suppression of locomotor activity in goldfish. Peptides 2012, 34, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Lugo, J.M.; Rodríguez, A.; Helguera, Y.; Morales, R.; González, O.; Acosta, J.; Besada, V.; Sánchez, A.; Estrada, M.P. Recombinant novel pituitary adenylate cyclase activating polypeptide (PACAP) from African catfish (Clarias gariepinus) authenticates its biological function as a growth promoting factor in lower vertebrates. J. Endocrinol. 2008, 197, 583–597. [Google Scholar] [CrossRef]
- Martinez, R.; Ubieta, K.; Herrera, F.; Forellat, A.; Morales, R.; de la Nuez, A.; Rodriguez, R.; Reyes, O.; Oliva, A.; Estrada, M.P. A novel GH secretagogue, A233, exhibits enhanced growth activity and innate immune system stimulation in teleosts fish. J. Endocrinol. 2012, 214, 409–419. [Google Scholar] [CrossRef]
- Martinez, R.; Carpio, Y.; Morales, A.; Lugo, J.M.; Herrera, F.; Zaldívar, C.; Carrillo, O.; Arenal, A.; Pimentel, E.; Estrada, M.P. Oral administration of the growth hormone secretagogue-6 (GHRP-6) enhances growth and non-specific immune responses in tilapia (Oreochromis sp.). Aquaculture 2016, 452, 304–310. [Google Scholar] [CrossRef]
- Lugo, J.M.; Oliva, A.; Morales, A.; Reyes, O.; Garay, H.E.; Herrera, F.; Cabrales, R.; Perez, E.; Estrada, M.P. The biological role of pituitary adenylate cyclase-activating polypeptide (PACAP) in growth and feeding behavior in juvenile fish. J. Pept. Sci. 2010, 16, 633–643. [Google Scholar] [CrossRef]
- Shepherd, B.S.; Eckert, S.M.; Parhar, I.S.; Vijayan, M.M.; Wakabayashi, I.; Hirano, T. The hexapeptide KP-102 (D-ala-D-beta-Nal-ala-trp-D-phe-lys-NH(2) stimulates growth hormone release in a cichlid fish (Oreochromis mossambicus). J. Endocrinol. 2000, 167, 7–10. [Google Scholar] [CrossRef]
- Shepherd, B.S.; Johnson, J.K.; Silverstein, J.T.; Parhar, I.S.; Vijayan, M.M.; McGuire, A.; Weber, G.M. Endocrine and orexigenic actions of growth hormone secretagogues in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. A 2007, 146, 390–399. [Google Scholar] [CrossRef]
- Rigos, G.; Kogiannou, D.; Padrós, F.; Cristofol, C.; Florio, D.; Fioravanti, M.; Zarza, C. Best therapeutic practices for the use of antibacterial agents in finfish aquaculture: A particular view on European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata) in Mediterranean aquaculture. Rev. Aquac. 2021, 13, 1285–1323. [Google Scholar] [CrossRef]
- Adelmann, M.; Kollner, B.; Bergmann, S.M.; Fischer, U.; Lange, B.; Weitschies, W.; Enzmann, P.J.; Fichtner, D. Development of an oral vaccine for immunisation of rainbow trout (Oncorhynchus mykiss) against viral haemorrhagic septicaemia. Vaccine 2008, 26, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Keppler, D.; Decker, K. Glycogen. Determination with amyloglucosidase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; pp. 1127–1131. [Google Scholar]
- Martínez, R.; Morales, C.; Arenal, A.; Morales, A.; Herrera, F.; González, V.; Estrada, M.P. Growth Hormone Secretagogue (A233) Improves Growth and Changes the Tissue Fatty Acid Profile in Juvenile Tilapia (Oreochromis niloticus). Lipids 2018, 53, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, R.; González, J.; Montoya, G.; Jara, A.; Ortíz, N.; Piedra, P.; Habit, E. Relación longitud-peso y factor de condición de los peces nativos del río San Pedro (cuenca del río Valdivia, Chile). Gayana (Concepción) 2012, 76, 86–100. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Ortega, A. Cultivo de Dorada (Sparus aurata); Fundación Observatorio Español de Acuicultura, Consejo Superior de Investigaciones Científicas, Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2008; pp. 1–44. [Google Scholar]
- Herbinger, C.M.; Friars, G.W. Correlation between condition factor and total lipid content in Atlantic salmon, Salmo salar L., parr. Aqua Res. 1991, 22, 527–529. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J. The involvement of growth hormone in growth regulation, energy homeostasis and immune function in the gilthead sea bream (Sparus aurata): A short review. Fish Physiol Biochem. 2000, 22, 135–144. [Google Scholar] [CrossRef]
- Simó-Mirabet, P.; Felip, A.; Estensoro, I.; Martos-Sitcha, J.A.; de las Heras, V.; Calduch-Giner, J.; Puyalto, M.; Karalazos, V.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Impact of low fish meal and fish oil diets on the performance, sex steroid profile and male-female sex reversal of gilthead sea bream (Sparus aurata) over a three-year production cycle. Aquaculture 2018, 490, 64–74. [Google Scholar] [CrossRef]
- Perera, E.; Simó-Mirabet, P.; Shin, H.S.; Rosell-Moll, E.; Naya-Catalá, F.; de las Heras, V.; Martos-Sitcha, J.A.; Karalazos, V.; Armero, E.; Pérez-Sánchez, J. Selection for growth is associated in gilthead sea bream (Sparus aurata) with diet flexibility, changes in growth patterns and higher intestine plasticity. Aquaculture 2019, 507, 349–360. [Google Scholar] [CrossRef]
- Triantaphyllopoulos, K.A.; Cartas, D.; Miliou, H. Factors influencing GH and IGF-I gene expression on growth in teleost fish: How can aquaculture industry benefit? Rev. Aquac. 2020, 12, 1637–1662. [Google Scholar] [CrossRef]
- Mingarro, M.; de Celis, S.V.R.; Astola, A.; Pendón, C.; Valdivia, M.M.; Pérez-Sánchez, J. Endocrine mediators of seasonal growth in gilthead sea bream (Sparus aurata): The growth hormone and somatolactin paradigm. Gen. Comp. Endocrinol. 2002, 128, 102–111. [Google Scholar] [CrossRef]
- Unniappan, S.; Lin, X.; Cervini, L.; Rivier, J.; Kaiya, H.; Kangawa, K.; Peter, R.E. Goldfish ghrelin: Molecular characterization of the complementary deoxyribonucleic acid, partial gene structure and evidence for its stimulatory role in food intake. Endocrinology 2002, 143, 4143–4146. [Google Scholar] [CrossRef] [PubMed]
- Unniappan, S.; Peter, R.E. In vitro and in vivo effects of ghrelin on luteinizing hormone and growth hormone release in goldfish. Am. J. Physiol.—Regul. Integr. Comp. Physiol. 2004, 286, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.E.; McIntyre, P.B.; Buels, K.S.; Gilbert, D.M.; Michel, E. Diet predicts intestine length in Lake Tanganyika’s cichlid fishes. Funct. Ecol. 2009, 23, 1122–1131. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose metabolism in fish: A review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Saccol, E.M.H.; Parrado-Sanabria, Y.A.; Gagliardi, L.; Jerez-Cepa, I.; Mourão, R.H.V.; Heinzmann, B.M.; Baldisserotto, B.; Pavanato, M.A.; Mancera, J.M.; Martos-Sitcha, J.A. Myrcia sylvatica essential oil in the diet of gilthead sea bream (Sparus aurata L.) attenuates the stress response induced by high stocking density. Aquac. Nutr. 2018, 24, 1381–1392. [Google Scholar] [CrossRef]
- Azeredo, R.; Machado, M.; Martos-Sitcha, J.A.; Martínez-Rodríguez, G.; Moura, J.; Peres, H.; Oliva-Teles, A.; Afonso, A.; Mancera, J.M.; Costas, B. Dietary tryptophan induces opposite health-related responses in the Senegalese sole (Solea senegalensis) reared at low or high stocking densities with implications in disease resistance. Front. Physiol. 2019, 10, 508. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Martos-Sitcha, J.A.; Queiroz, A.; Calduch-Giner, J.A.; Gonçalves, J.F.M.; Rocha, C.M.; Abreu, H.T.; Scharma, J.W.; Ozorio, R.O.A.; Pérez-Sánchez, J. Dietary supplementation of heat-treated Gracilaria and Ulva seaweeds enhanced acute hypoxia tolerance in gilthead sea bream (Sparus aurata). Biol. Open 2017, 6, 897–908. [Google Scholar] [CrossRef]
- Naya-Català, F.; Martos-Sitcha, J.A.; de las Heras, V.; Simó-Mirabet, P.; Calduch-Giner, J.À.; Pérez-Sánchez, J. Targeting the Mild-Hypoxia Driving Force for Metabolic and Muscle Transcriptional Reprogramming of Gilthead Sea Bream (Sparus aurata) Juveniles. Biology 2021, 10, 416. [Google Scholar] [CrossRef]
- Poudel, P.; Levesque, C.L.; Samuel, R.; St-Pierre, B. Dietary inclusion of Peptiva, a peptide-based feed additive, can accelerate the maturation of the fecal bacterial microbiome in weaned pigs. BMC Vet. Res. 2020, 16, 1–13. [Google Scholar] [CrossRef]
- Herrera, F.; Velazquez, J.; Lugo, J.M.; Orellana, P.; Ruiz, J.; Vega, M.; Romero, A.; Santos, N.; Ramses, G.; Rodríguez-Ramos, T.; et al. Oral Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) formulation modified muscle fatty acid profile and cytokines transcription in head kidney in rainbow trout (Oncorhynchus mykiss) fingerlings. Aquac. Rep. 2021, 20, 100772. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| CTRL | D100 | D500 | p1 | |
|---|---|---|---|---|
| Initial body weight (g) | 20.32 ± 0.08 | 20.21 ± 0.05 | 20.13 ± 0.03 | 0.129 |
| Final body weight (g) | 49.57 ± 1.45 a | 56.53 ± 1.56 b | 58.50 ± 0.46 b | 0.006 |
| Final fork length (cm) | 14.58 ± 0.14 | 14.50 ± 0.20 | 14.85 ± 0.20 | 0.374 |
| K 2 | 1.85 ± 0.03 a | 1.99 ± 0.04 b | 2.00 ± 0.05 b | 0.015 |
| Weight gain (%) 3 | 144.0 ± 7.1 a | 179.8 ± 7.6 b | 190.6 ± 2.32 b | 0.004 |
| SGR (%) 4 | 0.92 ± 0.03 a | 1.07 ± 0.03 b | 1.10 ± 0.01 b | 0.004 |
| Feed intake (g DM/fish) | 44.67 ± 0.71 a | 47.01 ± 0.69 b | 48.36 ± 0.58 b | 0.017 |
| FE (%) 5 | 63 ± 2 a | 77 ± 2 b | 79 ± 1 b | <0.001 |
| HSI (%) 6 | 1.59 ± 0.07 | 1.62 ± 0.08 | 1.72 ± 0.10 | 0.491 |
| MSI (%) 7 | 0.69 ± 0.05 | 0.59 ± 0.12 | 0.56 ± 0.05 | 0.120 |
| ILI (%) 8 | 107.4 ± 5.8 | 105.0 ± 7.9 | 109.3 ± 7.5 | 0.914 |
| CTRL | D100 | D500 | p1 | |
|---|---|---|---|---|
| Plasma glucose (mM) | 1.67 ± 0.05 | 1.59 ± 0.02 | 1.69 ± 0.03 | 0.089 |
| Plasma lactate (mM) | 3.14 ± 0.24 a | 2.39 ± 0.22 b | 3.25 ± 0.19 a | 0.013 |
| Plasma triglycerides (mM) | 2.61 ± 0.24 | 2.41 ± 0.21 | 2.29 ± 0.14 | 0.529 |
| Plasma proteins (mg·mL−1) | 38.85 ± 2.79 | 40.25 ± 1.65 | 41.34 ± 1.58 | 0.699 |
| Plasma cholesterol (mg·dL−1) | 258.7 ± 8.9 a | 326.7 ± 12.4 b | 318.1 ± 10.9 b | <0.001 |
| Plasma cortisol (ng·mL−1) | 15.19 ± 1.14 | 16.04 ± 0.95 | 15.84 ± 0.73 | 0.808 |
| Hepatic glucose (µmol·gww−1) | 2.87 ± 0.20 | 2.56 ± 0.24 | 3.31 ± 0.34 | 0.150 |
| Hepatic glycogen (µmol·gww−1) | 19.77 ± 0.50 a | 20.52 ± 0.73 a | 24.33 ± 1.49 b | 0.007 |
| Hepatic triglycerides (µmol·gww−1) | 137.6 ± 13.1 | 148.6 ± 11.0 | 147.0 ± 14.0 | 0.807 |
| Muscular glucose (µmol·gww−1) | 1.94 ± 0.19 | 1.97 ± 0.22 | 2.05 ± 0.20 | 0.939 |
| Muscular glycogen (µmol·gww−1) | 1.02 ± 0.19 | 1.13 ± 0.26 | 1.38 ± 0.27 | 0.556 |
| Muscular triglycerides (µmol·gww−1) | 80.7 ± 7.9 | 78.1 ± 5.7 | 81.4 ± 7.2 | 0.941 |
| Muscular lactate (µmol·gww−1) | 51.66 ± 3.54 | 56.36 ± 2.71 | 62.80 ± 3.12 | 0.057 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Viera, L.; Martí, I.; Martínez, R.; Perera, E.; Estrada, M.P.; Mancera, J.M.; Martos-Sitcha, J.A. Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata. Fishes 2022, 7, 31. https://doi.org/10.3390/fishes7010031
Rodríguez-Viera L, Martí I, Martínez R, Perera E, Estrada MP, Mancera JM, Martos-Sitcha JA. Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata. Fishes. 2022; 7(1):31. https://doi.org/10.3390/fishes7010031
Chicago/Turabian StyleRodríguez-Viera, Leandro, Ignacio Martí, Rebeca Martínez, Erick Perera, Mario Pablo Estrada, Juan Miguel Mancera, and Juan Antonio Martos-Sitcha. 2022. "Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata" Fishes 7, no. 1: 31. https://doi.org/10.3390/fishes7010031
APA StyleRodríguez-Viera, L., Martí, I., Martínez, R., Perera, E., Estrada, M. P., Mancera, J. M., & Martos-Sitcha, J. A. (2022). Feed Supplementation with the GHRP-6 Peptide, a Ghrelin Analog, Improves Feed Intake, Growth Performance and Aerobic Metabolism in the Gilthead Sea Bream Sparus aurata. Fishes, 7(1), 31. https://doi.org/10.3390/fishes7010031