1. Introduction
Growth is one of the key metrics studied in fish population dynamics because it affects present and future population status [
1]. This metric can also provide important information regarding prey availability and habitat quality (i.e., fast growth indicates sufficient prey resources and habitat quality, whereas slow growth may suggest the opposite) [
2]. Therefore, having a basic understanding of growth rate for individuals in a study population, and how this metric compares to other populations (e.g., those known to exhibit fast growth), can help guide conservation and management efforts. This information, however, can be challenging to gather for rare species, especially during early life history. For instance, the pallid sturgeon (
Scaphirhynchus albus) is a federally endangered species inhabiting the Mississippi and Missouri rivers in the United States [
3], and published literature only documents seven wild-produced exogenously feeding age-0 pallid sturgeon (20–48 mm in length) that have been genetically confirmed [
4,
5].
Given the limited information available for pallid sturgeon during early life history, the stocking of age-0 individuals is one potential tool to address this knowledge gap [
6]. In the Missouri River, pallid sturgeon propagation began in the early 1990s, resulting in an established, long-term stocking program capable of marking and releasing a wide range of ages [
7]. During July of 2018, pallid sturgeon ranging from either 53–56 or 41–42 days old [
6] were released into the lower Missouri River (LMOR; Gavins Point Dam, South Dakota downstream to the Mississippi River confluence at St. Louis, Missouri) as part of two stockings conducted during consecutive weeks. The subsequent capture of some of these individuals provided the opportunity to learn about a potentially important and difficult-to-study life stage for an endangered species in this large river system. Gemeinhardt et al. (2021) [
6] documented dispersal distance following these stocking events, but there is also an opportunity to assess growth in this river and to make length-at-age comparisons with individuals raised entirely in Gavins Point National Fish Hatchery (GPNFH; Yankton, South Dakota, USA). Given the energetically favorable rearing conditions coupled with abundant food resources (i.e., all pallid sturgeon were fed until satiation), hatcheries can provide a benchmark for assessing growth. For example, Coho Salmon
Oncorhynchus kisutch reared from fry to smolt in a hatchery were significantly longer and heavier than siblings reared in a natural river side-channel [
8].
Food limitation is a potential limiting factor for pallid sturgeon recruitment to age 1 in the LMOR (e.g., [
9]), but information on prey consumption in this system is limited given the small number of wild pallid sturgeon captures, which have consisted of individuals <50 mm long [
5]. As such, laboratory work has also been conducted to better understand prey use and growth. For example, Porreca et al. (2017) [
10] modeled the growth rate potential of age-0 pallid sturgeon (average size = 161 mm) as well as the closely related shovelnose sturgeon (
Scaphirhynchus platorynchus, average size = 166 mm) to compare the ability of these two congeners to assimilate prey. They reported that pallid sturgeon yielded a lower modeled growth rate potential in most, but not all, of the artificial habitat treatments; however, these treatments may not reflect river conditions and relative differences do not necessarily indicate that pallid sturgeon are unable to grow and survive in the wild. Thus, the purpose of our study is to use field and hatchery data to provide insight regarding growth and potential food limitation for age-0 pallid sturgeon (stocked at fork lengths ranging from approximately 70 to 135 mm) that have spent over 80% of their life in the LMOR.
2. Materials and Methods
Pallid sturgeon propagation involved spawning six wild females and eight wild males (using a total of eight crosses) in May of 2018 at GPNFH [
6]. Following hatch, offspring rearing occurred for either 53–56 or 41–42 days in water temperatures of 21.1–22.3 °C [
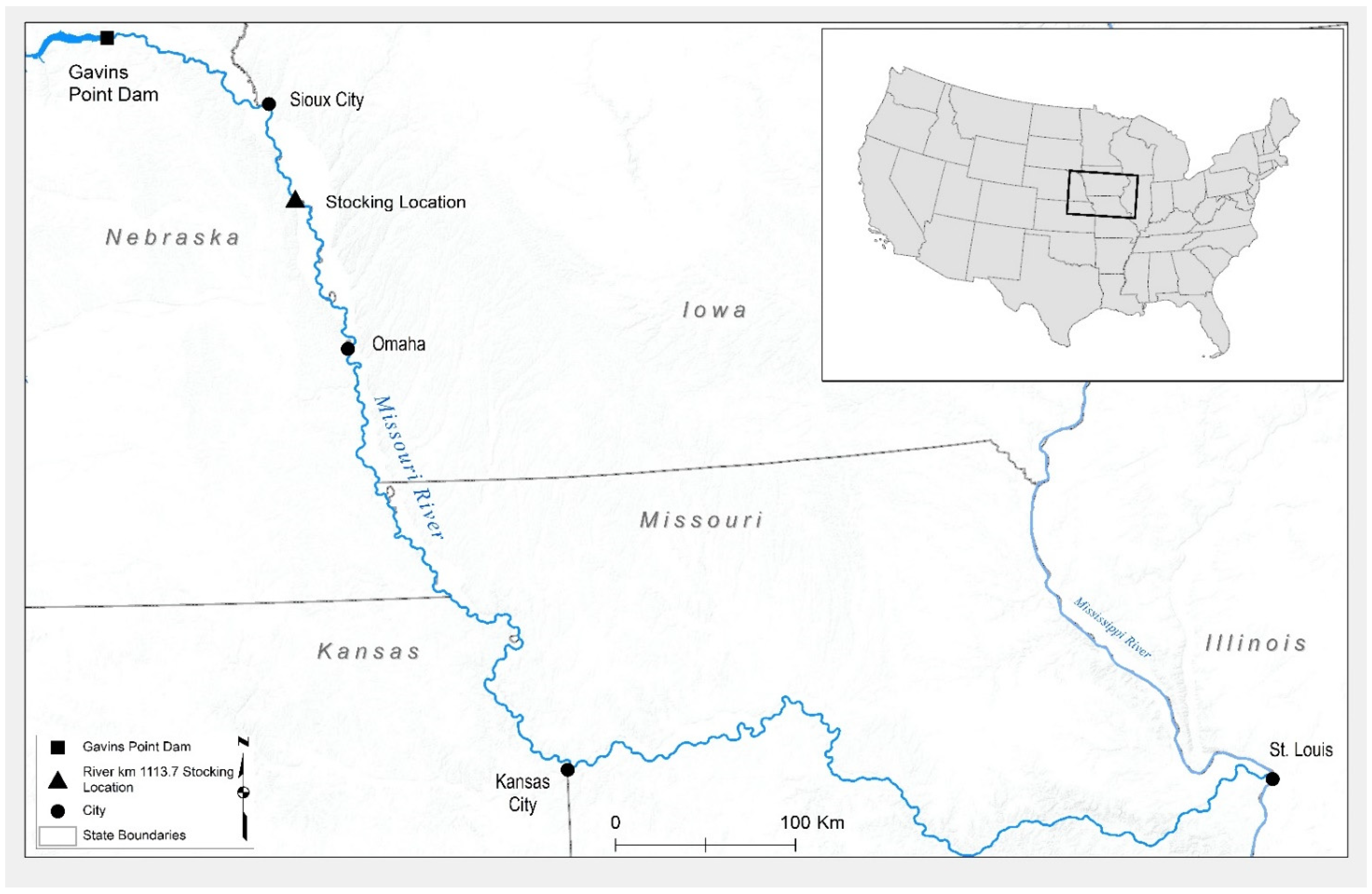
6]. Stocking occurred during consecutive weeks in July 2018 at river km 1113.7 near Decatur, Nebraska (
Figure 1). During the first stocking event 9–12 July, 4861 fish were released at a mean fork length of 118 mm. During the second stocking event on 18-19 July, 3634 individuals were released at a mean fork length of 80 mm. Prior to stocking, hatchery staff marked all individuals with Visual Implant Elastomer (VIE) tags to allow for the quick and reliable detection of post-stocking capture (
Figure 2). Additionally, colored VIE tags were used to distinguish between stockings.
Pallid sturgeon were sampled from July 2018 through June 2021 with baited trotlines (set overnight) or otter trawls (either 4-mm or 16-mm mesh) during routine annual sampling efforts. We compared the growth of captured pallid sturgeon to fish reared in GPNFH (length recorded prior to release into the LMOR) during 2002–2018 by constructing length-at-age scatterplots for individuals captured from both stocking events as well as fish raised at GPNFH. Linear regression with the associated 95% prediction interval was conducted for GPNFH fish to provide context to the growth of captured individuals. We also performed linear regression analysis specific to each stocking event and used analysis of covariance (ANCOVA), with age as the covariate, to assess potential differences in slope and y-intercept values among stocking 1, stocking 2, and GPNFH. If slope values were significantly different (i.e., not parallel), then y-intercept comparisons were not conducted [
11]. Three individuals from the first stocking were removed from the growth analysis, as they had spent little time in the LMOR (13–41 days) prior to capture.
3. Results
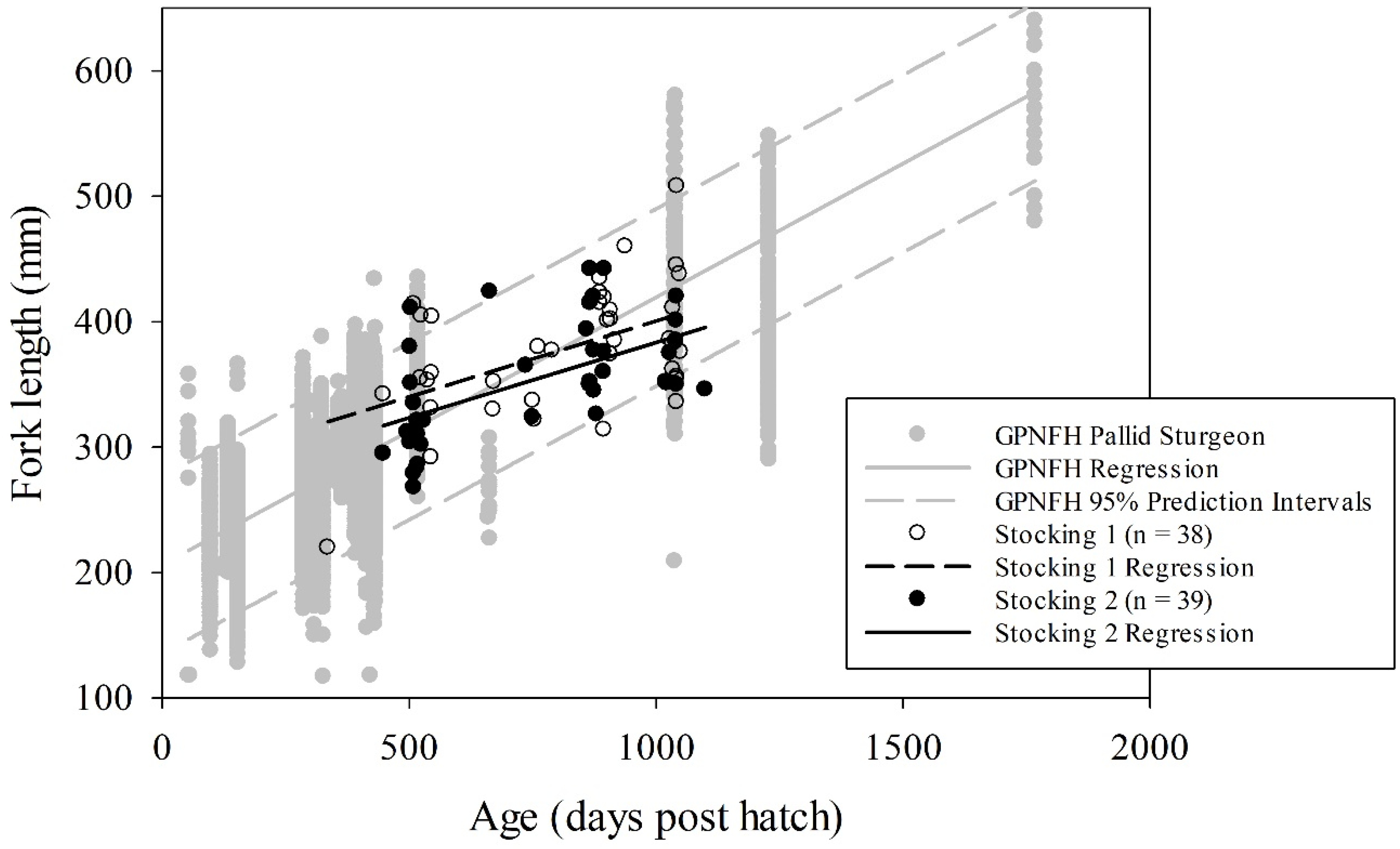
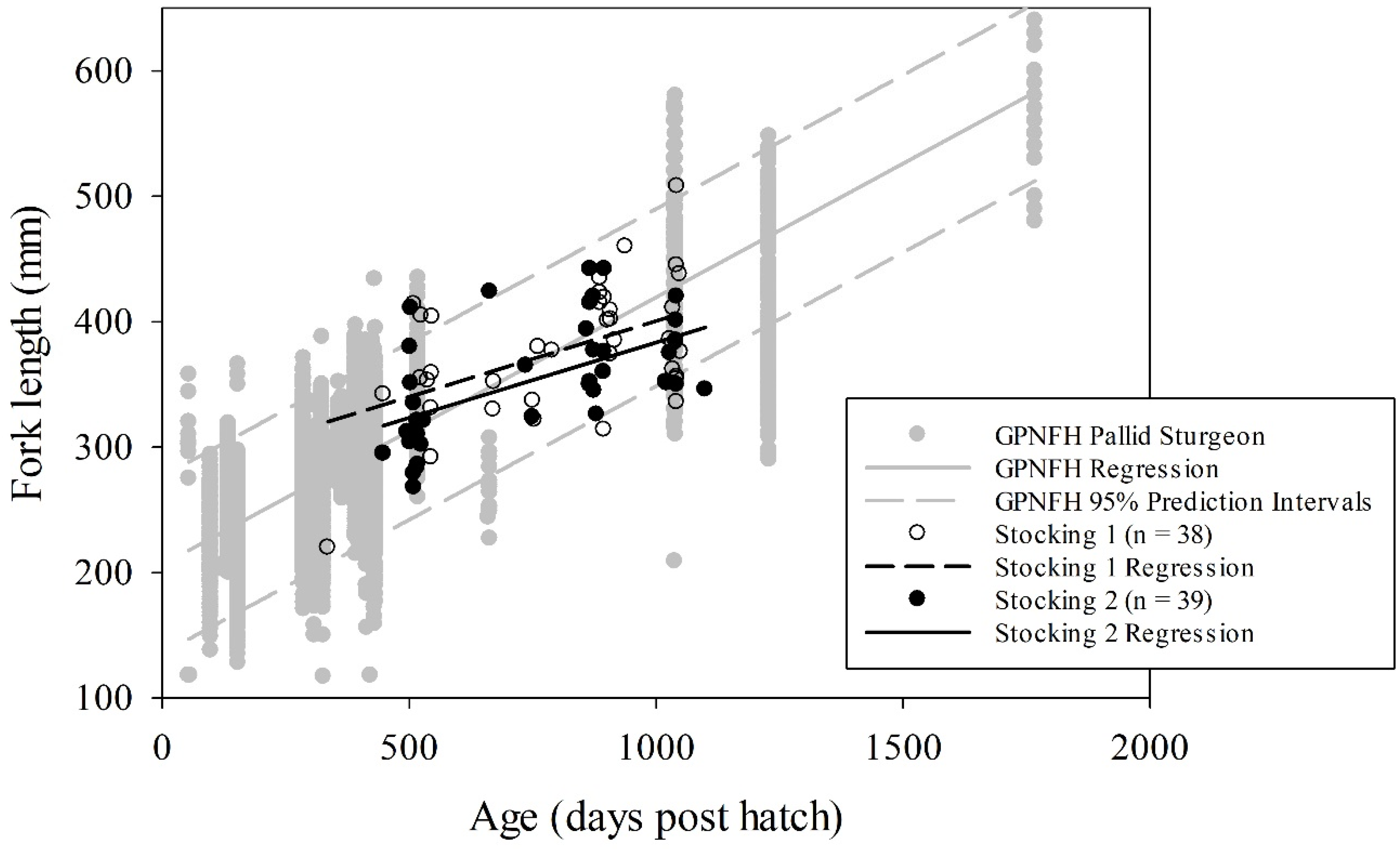
Trotlines accounted for 88% of captures during this study, while otter trawls accounted for the remaining 12%. We documented 77 stocked pallid sturgeon that recruited to age-1 and older (38 from stocking 1 and 39 from stocking 2). These individuals spent a minimum of 83% of their post-hatch life in the LMOR. The length-at-age linear regressions (
Figure 3) were significant for stocking 1 (r
2 = 0.25,
p = 0.001,
df = 1), stocking 2 (r
2 = 0.32,
p < 0.001,
df = 1), and GPNFH (r
2 = 0.58,
p < 0.001,
df = 1). Over 78% and 87% of stocking 1 and stocking 2 captures, respectively, fell within the 95% prediction interval for fish reared at GPNFH (
Figure 3). Only 11% and 8% of stocking 1 and 2 captures, respectively, fell below the 95% prediction interval, while 11% and 5% of stocking 1 and 2 captures, respectively, fell above the 95% prediction interval (
Figure 3). Regarding the ANCOVA, stocking 1 and 2 did not have different slope (
F = 0.0,
p = 0.98) or y-intercept (
F = 3.15,
p = 0.08) values, whereas stocking 1 (
F = 11.2,
p < 0.001) and stocking 2 (
F = 12.6,
p < 0.001) both had different slope values compared to GPNFH.
4. Discussion
This study provided a rare opportunity to document the survival of relatively small age-0 pallid sturgeon stocked into the LMOR and to assess subsequent growth. Many of the hypothesized factors limiting pallid sturgeon population growth involve the age-0 life stage, and one of the main recovery objectives is to increase recruitment to age 1 [
9]. However, relatively little information is available for much of this life stage because wild age-0 pallid sturgeon captures are rare [
5], and most age-0 pallid sturgeon are stocked late in the year at larger sizes and older ages relative to this study. For example, Steffensen et al. (2019) [
7] noted that from 1999 to 2015, the average age-0 pallid sturgeon stocked was 231 mm long and 153 days old. During the present study, we captured 0.94% of the fish stocked during July of 2018, and our captures confirmed that at least 0.90% recruited to age 1 or older. Despite the relatively small body size for both stockings during July of 2018, the rate of capture was high relative to most other age-0 stockings, as Steffensen et al. (2019) [
7] reported an average age-0 capture rate of 0.34%. These findings may have implications for future decisions regarding age-0 stocking size, but other factors (e.g., parental crosses, river conditions at the time of release) may have also influenced subsequent survival and capture rates [
7].
This study also provided an opportunity to better understand if insufficient prey resources, which is a hypothesized limiting factor for age-0 pallid sturgeon [
9], may be affecting growth and survival of these stocked individuals. Porreca et al. (2017) [
10] observed subtle physiological differences (e.g., alimentary canal length) between pallid sturgeon (average size = 161 mm) and shovelnose sturgeon (average size = 166 mm) during a laboratory study. They also modeled growth rate potential for both species hypothesizing that interspecific differences in the ability to metabolize food may account for the difference in survivorship between these species in the wild. However, Gosch et al. (2019) [
5] discussed how these laboratory comparisons by Porreca et al. (2017) [
10] may not be applicable to sturgeon in the LMOR. For example, the pallid sturgeon growth rate potential was higher than shovelnose sturgeon in the gravel-dominant (mean diameter = 1 cm) substrate treatments with approximately 80% or more gravel [
10], and widely available sand dune habitat in the LMOR may also provide important energetic refugia that could benefit pallid sturgeon growth [
12]. The individuals captured during the present study allowed us to directly assess pallid sturgeon growth in the wild because fish were stocked at smaller sizes, and subsequently captured at larger sizes, than the individuals studied by Porreca et al. (2017) [
10]. Given that individuals were fed until satiation in an energetically favorable environment, we were not surprised that the overall length-at-age trend line for GPNFH was steeper indicating faster growth relative to stocking 1 and 2. However, we did not expect the high degree of overlap among the length-at-age scatterplots with values for stocking 1 and 2 always falling within the range of values observed for GPNFH. We were also surprised that only 9% of captured fish fell below the 95% prediction intervals for GPNFH, and that the length-at-age trend lines indicated higher growth for individuals <800 days old and <600 days old from stocking 1 and stocking 2, respectively, compared to GPNFH (
Figure 3). These unexpected observations may suggest that the ANCOVA results were not biologically significant. Overall, this study demonstrates that the LMOR is capable of yielding hatchery-level growth for young pallid sturgeon, which is inconsistent with the Porreca et al. (2017) [
10] hypothesis that pallid sturgeon are endangered due to a relatively limited ability to assimilate prey in the contemporary LMOR.
One potential explanation for the observed growth is that extensive flooding during 2019 provided increased prey resources in the LMOR. However, Gosch et al. (2021) [
13] documented that age-0
Scaphirhynchus sturgeon (≤120 mm) captured from floodplain and mainstem habitats in 2019 did not have increased condition (based on length-weight relationships) compared to other years. Additionally, condition declined for many juvenile pallid sturgeon (400 to 699 mm) following the 2011 LMOR flood [
14]. Previous research has also suggested that wild age-0 pallid sturgeon consume similar prey in similar amounts as the abundant shovelnose sturgeon [
4,
5], and that natural reproduction was responsible for an estimated 8% of pallid sturgeon captures in the LMOR from 2003 to 2015 [
15]. Our results, coupled with this prior research, suggest a lack of evidence for the hypothesis that LMOR pallid sturgeon are limited by insufficient prey resources, and we recommend focusing research efforts on other hypothesized limiting factors (e.g., altered drift dynamics) [
16]. If future research suggests otherwise, an adaptive management framework is in place to reprioritize prey-related research [
5]. Gemeinhardt et al. (2021) [
6] demonstrated that artificial stockings can be a valuable tool for understanding the dispersal of young pallid sturgeon, and this study further highlights the potential benefits of endangered species stocking efforts.


 ,
, {kind=link}
{kind=link}
{kind=link}