Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus)



Abstract
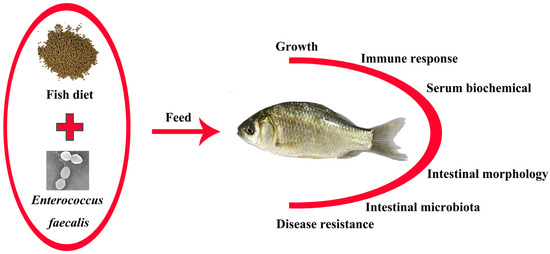
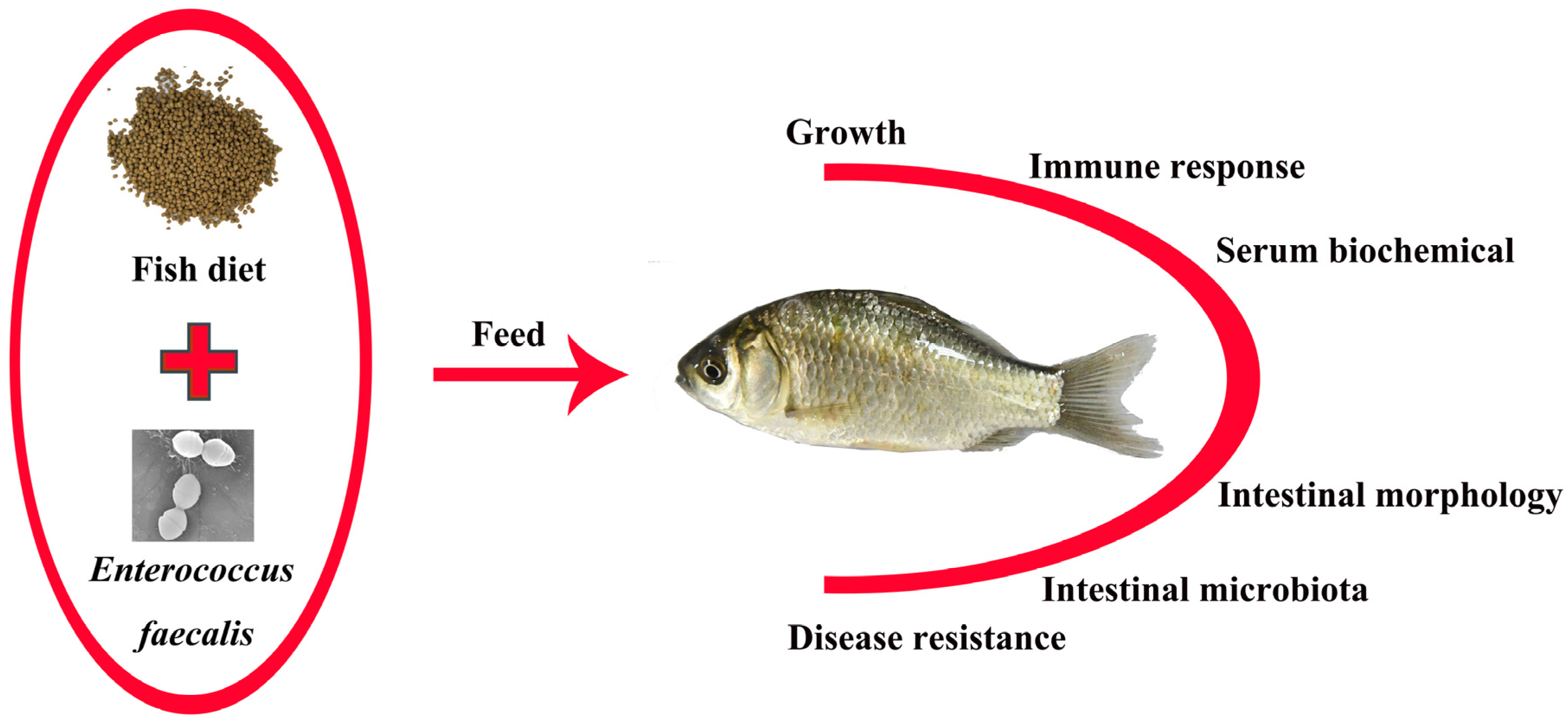
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Laboratory Fish and Rearing
2.3. Growth Performance
2.4. Sample Collection
2.5. Serum Biochemical Analysis
2.6. Real-Time PCR Analysis of Immune-Related Genes
2.7. Histopathology Analysis
2.8. Genomic DNA Extraction and 16S rRNA Gene Sequencing
2.9. Challenge Test
2.10. Statistical Analyses
3. Results
3.1. Growth Performance
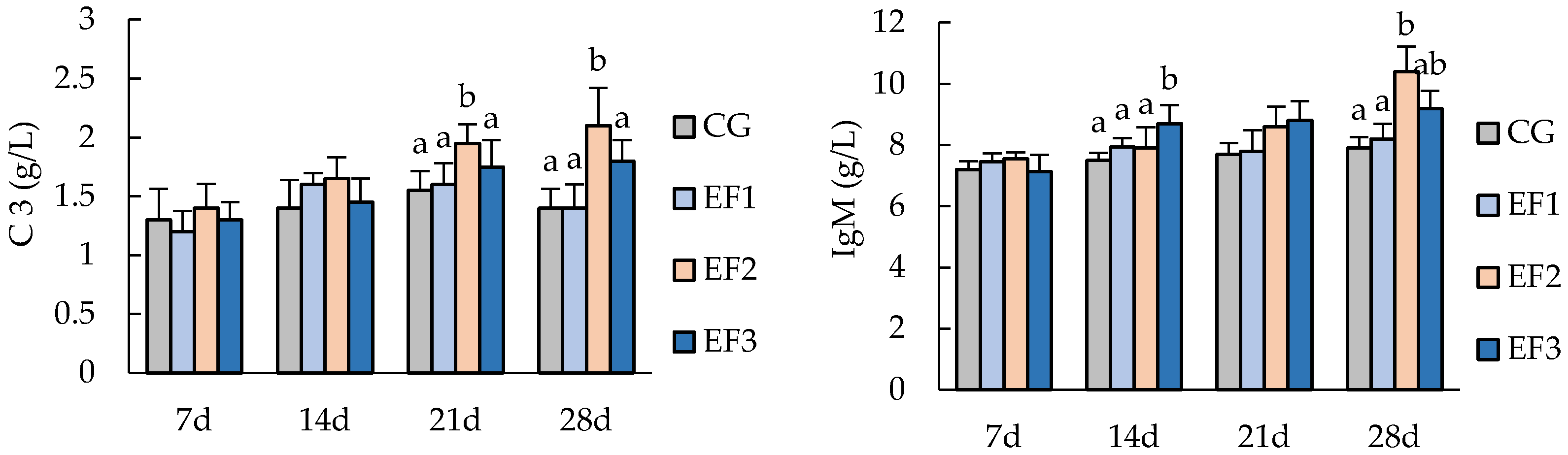
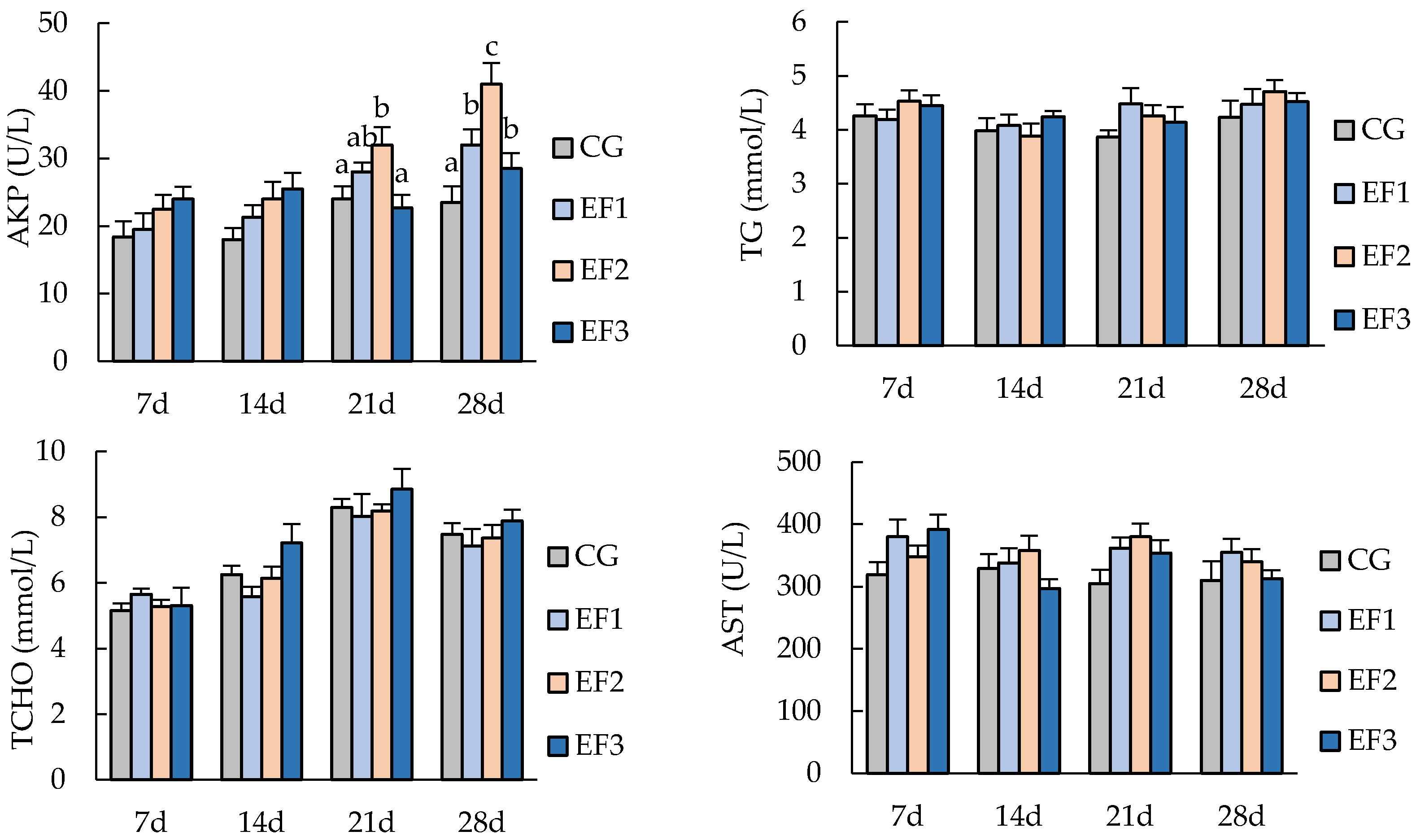
3.2. Serum Biochemical Parameters
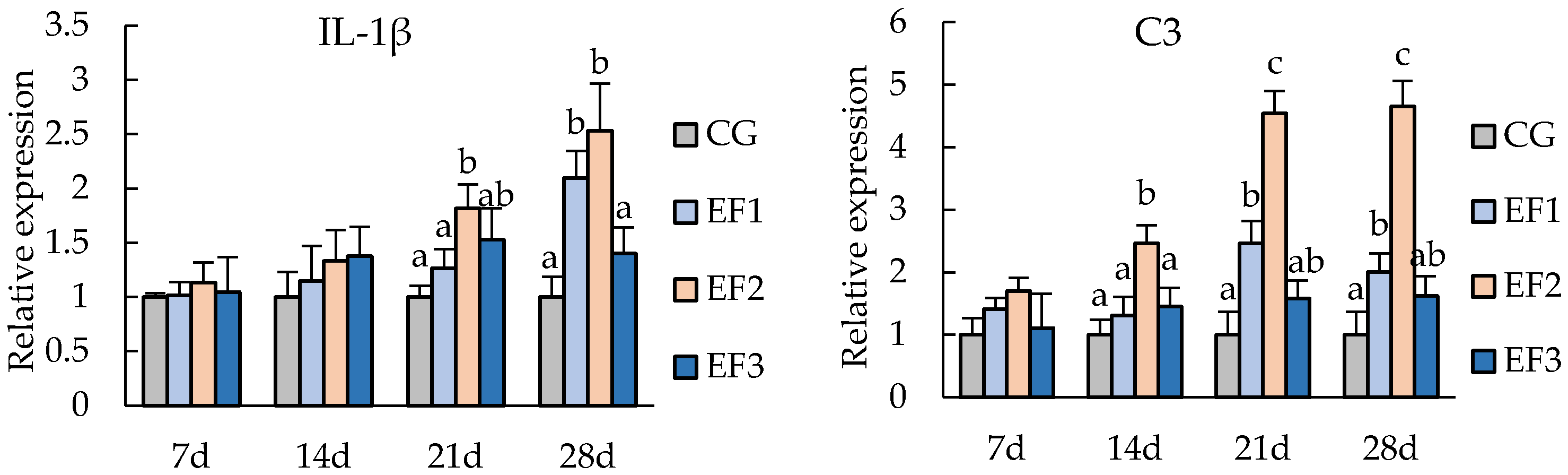
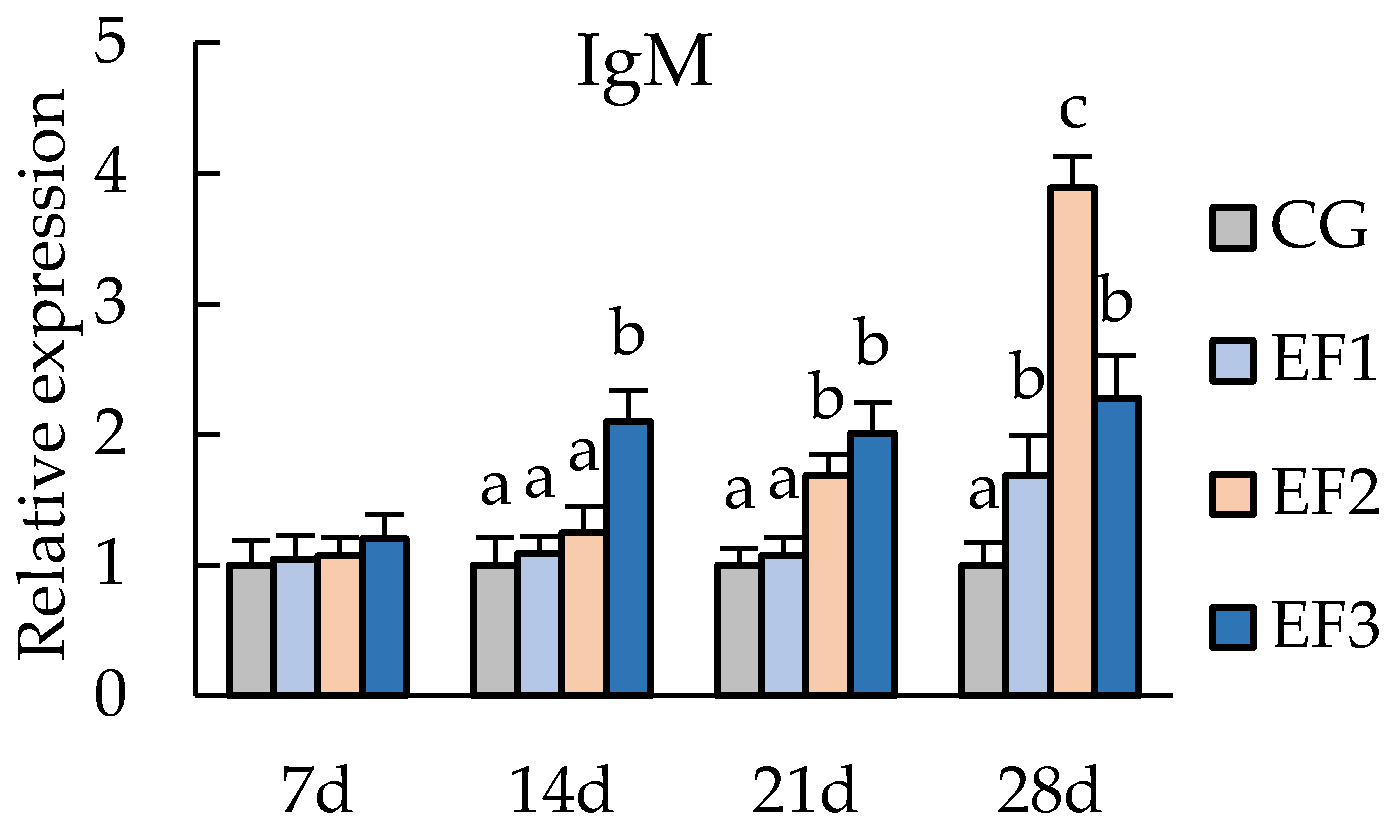
3.3. Changes in Immunity-Related Genes Expressions
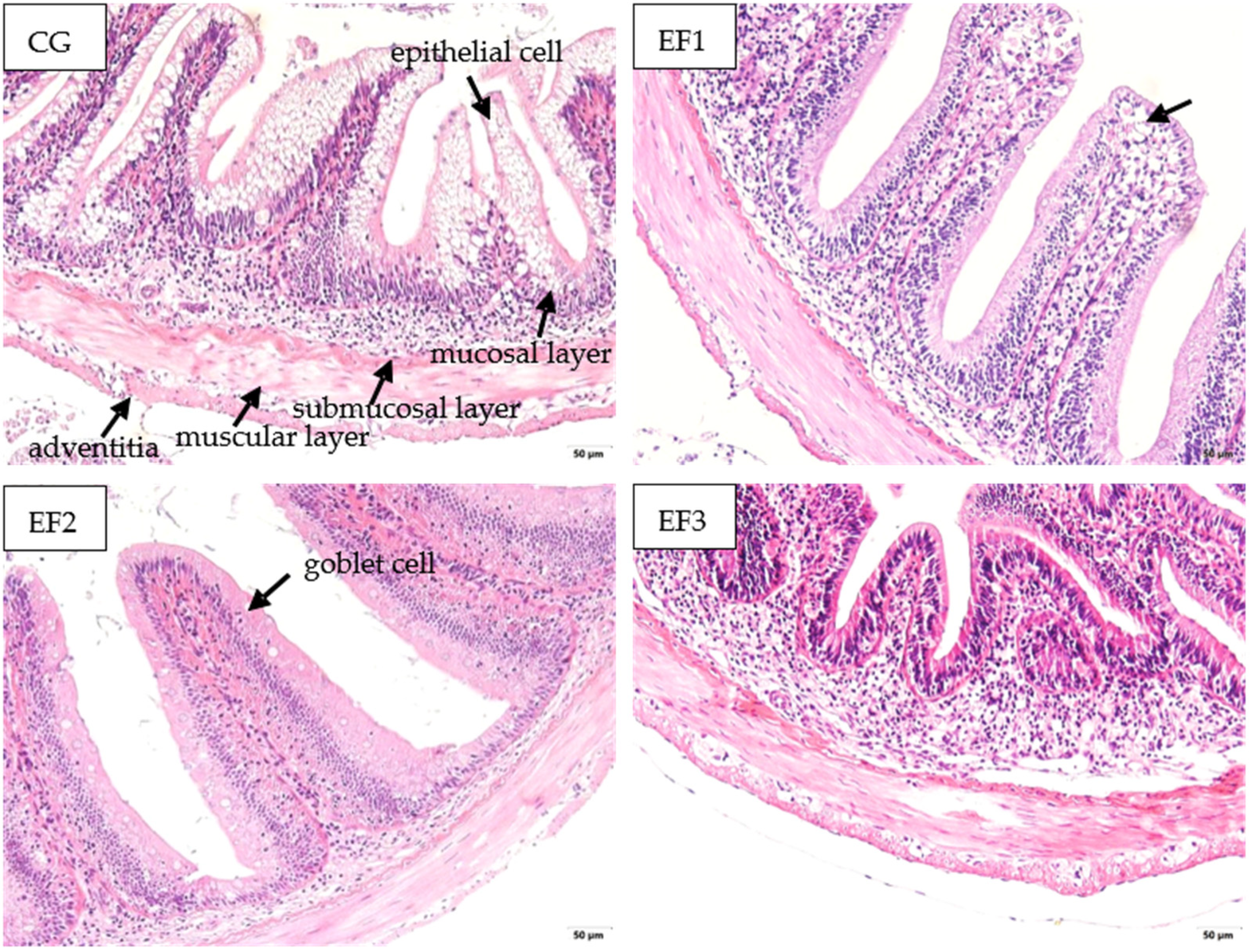
3.4. Changes in Intestinal Morphology
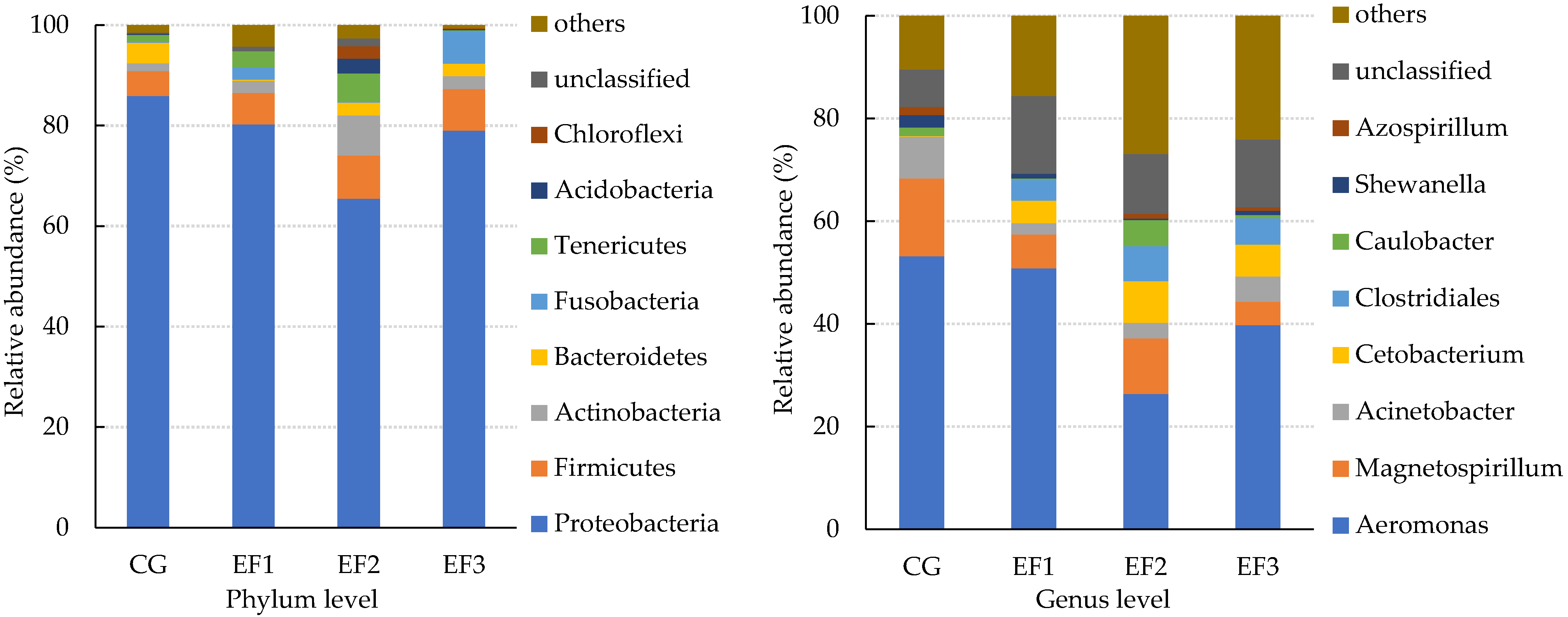
3.5. Changes in the Intestinal Microbiota
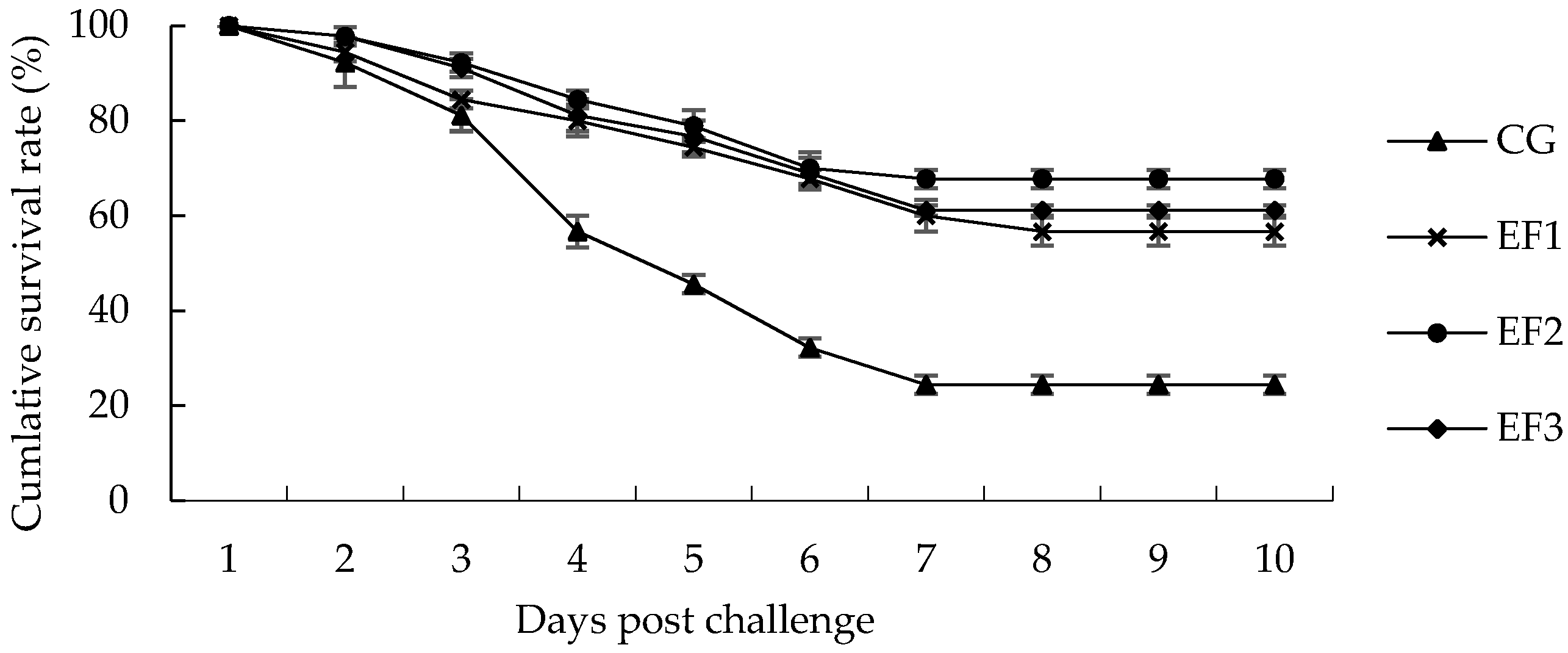
3.6. Challenge Test
4. Discussion
4.1. Growth Performance
4.2. Serum Biochemical Parameters
4.3. Immune Related Gene Expressions
4.4. Intestinal Morphology
4.5. Changes in the Intestinal Microbiota
4.6. Disease Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, M.; Xu, X.; Li, S.; Wu, T.; Yan, Y.; Gu, W. The metabolic responses of crucian carp blood to Cyprinid herpesvirus 2 infection. Aquaculture 2019, 498, 72–82. [Google Scholar] [CrossRef]
- Yang, F.; Yang, F.; Wang, G.; Kong, T.; Liu, B. Pharmacokinetics of florfenicol and its metabolite florfenicol amine in crucian carp (Carassius auratus) at three temperatures after single oral administration. Aquaculture 2019, 503, 446–451. [Google Scholar] [CrossRef]
- Fisheries and Fisheries Administration Bureau of Ministry of Agriculture and Industry. China Fisheries Yearbook; China Agriculture Press: Beijing, China, 2021; p. 25.
- Wu, S.J. Dietary Astragalus membranaceus polysaccharide ameliorates the growth performance and innate immunity of juvenile crucian carp (Carassius auratus). Int. J. Biol. Macromol. 2020, 149, 877–881. [Google Scholar] [CrossRef]
- Chen, F.; Sun, J.F.; Han, Z.R.; Yang, X.J.; Xian, J.A.; Lv, A.J.; Hu, X.C.; Shi, H.Y. Isolation, Identification and Characteristics of Aeromonas veronii From Diseased Crucian Carp (Carassius auratus gibelio). Front. Microbiol. 2019, 10, 2742. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yang, Y.; Liu, J.; Awan, F.; Lu, C.; Liu, Y. Inhibition of Aeromonas hydrophilainduced intestinal inflammation and mucosal barrier function damage in crucian carp by oral administration of Lactococcus lactis. Fish Shellfish Immunol. 2018, 83, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zeng, L.B.; Zhang, H.; Zhou, Z.; Ma, J.; Fan, Y.D. Cyprinid herpesvirus 2 infection emerged in cultured gibel carp, Carassius auratus gibelio in China. Vet. Microbiol. 2013, 166, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Rawls, J.F. Intestinal microbiota composition in fishes is influenced by host ecology and environment. Mol. Ecol. 2012, 21, 3100–3102. [Google Scholar] [CrossRef]
- She, R.; Li, T.T.; Luo, D.; Li, J.B.; Yin, L.Y.; Li, H.; Liu, Y.M.; Li, X.Z.; Yan, Q.G. Changes in the Intestinal Microbiota of Gibel Carp (Carassius gibelio) Associated with Cyprinid herpesvirus 2 (CyHV-2) Infection. Curr. Microbiol. 2017, 74, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Rawls, J.F.; Samuel, B.S.; Gordon, J.I. Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. USA 2004, 101, 4596–4601. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Peera, H.; James, V. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenter 2013, 6, 39–51. [Google Scholar]
- Seyed, H.H.; Einar, R.; Alireza, S.M.; Maria, Á.E. Probiotic, prebiotic and synbiotic supplements in sturgeon aquaculture: A review. Rev. Aquacult. 2016, 8, 89–102. [Google Scholar]
- Matsuura, Y.; Takasaki, M.; Miyazawa, R.; Nakanishi, T. Stimulatory effects of heat-killed Enterococcus faecalis on cell-mediated immunity in fish. Dev. Comp. Immunol. 2017, 74, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nueno-Palop, C.; Narbad, A. Probiotic assessment of Enterococcus faecalis CP58 isolated from human gut. Int. J. Food Microbiol. 2011, 145, 390–394. [Google Scholar] [CrossRef]
- Sayyed, K.A.; Einar, R.; Fatimah, M.Y.; Hassan, M.D.; Aini, I. Properties of Enterococcus faecalis, a new probiotic bacterium isolated from the intestine of snakehead fish (Channa striatus Bloch). Afr. J. Microbiol. Res. 2014, 8, 2215–2222. [Google Scholar] [CrossRef]
- Castro, M.S.; Molina, M.A.; Azpiroz, M.B.; Díaz, A.M.; Ponzio, R.; Sparo, M.D.; Manghi, M.A.; Canellada, A.M. Probiotic activity of Enterococcus faecalis CECT7121: Effects on mucosal immunity and intestinal epithelial cells. J. Appl. Microbiol 2016, 121, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Q.; Luo, L.; Zhou, Y.; Ling, H.Y.; Yang, Q.H.; Qi, D.S. Dietary administration of Enterococcus faecalis affects the growth, disease resistance and immune function of tilapia (Oreochromis niloticus). Aquacult. Rep. 2020, 18, 100440. [Google Scholar] [CrossRef]
- Uriel, R.E.; Shuichi, S.; Yutaka, H.; Hiroshi, F.; John, S. Effects of Inactivated Enterococcus faecalis and Mannan Oligosaccharide and Their Combination on Growth, Immunity, and Disease Protection in Rainbow Trout. N. Am. J. Aquacult. 2013, 75, 416–428. [Google Scholar]
- Sayyed, K.A.; Einar, R.; Fatimah, M.Y.; Hassan, H.D. Dietary supplement of Enterococcus faecalis on digestive enzyme activities, short-chain fatty acid production, immune system response and disease resistance of Javanese carp (Puntius gonionotus, Bleeker 1850). Aquacult. Nutr. 2017, 23, 331–338. [Google Scholar]
- Yang, Q.H.; Lü, Y.L.; Zhang, G.; Gong, Y.; Li, Z.Z.; Tran, N.T.; He, Y.Y.; Zhu, C.H.; Lu, Y.S.; Zhang, Y.L.; et al. Lactic acid bacteria, Enterococcus faecalis Y17 and Pediococcus pentosaceus G11, improved growth performance, and immunity of mud crab (Scylla paramamosain). Fish Shellfish Immunol. 2019, 93, 135–143. [Google Scholar] [CrossRef]
- Yang, G.; Cao, H.Z.; Jiang, W.H.; Hu, B.Q.; Jian, S.Q.; Wen, C.G.; Kajbaf, K.; Kumar, V.; Tao, Z.Y.; Peng, M. Dietary supplementation of Bacillus cereus as probiotics in Pengze crucian carp (Carassius auratus var. Pengze): Effects on growth performance, fillet quality, serum biochemical parameters and intestinal histology. Aquac. Res. 2019, 50, 2207–2217. [Google Scholar] [CrossRef]
- Kong, Y.D.; Li, M.; Tian, J.X.; Zhao, L.H.; Kang, Y.H.; Zhang, L.; Wang, G.Q.; Shan, X.F. Effects of recombinant Lactobacillus casei on growth performance, immune response and disease resistance in crucian carp, Carassius auratus. Fish Shellfish Immunol. 2020, 99, 73–85. [Google Scholar] [CrossRef]
- He, M.s.; Liu, G.y.; Liu, Y.h.; Yang, K.C.; Qi, X.Z.; Huang, A.G.; Liu, T.Q.; Wang, G.X.; Wang, E.L. Effects of geniposide as immunostimulant on the innate immune response and disease resistance in crucian carp. Aquaculture 2020, 529, 735713. [Google Scholar] [CrossRef]
- Fan, Y.D.; Zhang, X.P.; Zhou, Y.; Jiang, N.; Liu, W.Z.; Zeng, L.B. Molecular cloning of Gibel carp (Carassius auratus gibelio) complement component C3 and its expression profile after Cyprinid herpesvirus 2 infection. J. Vet. Med. Sci. 2020, 82, 47–55. [Google Scholar] [CrossRef]
- Tang, J.; Wang, W.; Jiang, Y.; Chu, W.H. Diazinon exposure produces histological damage, oxidative stress, immune disorders and gut microbiota dysbiosis in crucian carp (Carassius auratus gibelio). Environ. Pollut. 2021, 269, 116–129. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Grice, A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Nakhro, K.; Chowdhury, S.; Kamilya, D. Effects of potential probiotic Bacillus amyloliquefaciens FPTB16 on systemic and cutaneous mucosal immune responses and disease resistance of catla (Catla catla). Fish Shellfish Immunol. 2013, 35, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Arun, C.; Rahul, S. Probiotics in aquaculture: A promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar]
- Wang, Y.B.; Tian, Z.Q.; Yao, J.T.; Li, W.F. Effect of probiotics, Enteroccus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture 2008, 277, 203–207. [Google Scholar] [CrossRef]
- Li, C.; Ren, Y.C.; Jiang, S.H.; Zhou, S.; Zhao, J.S.; Wang, R.J.; Li, Y.M. Effects of dietary supplementation of four strains of lactic acid bacteria on growth, immune-related response and genes expression of the juvenile sea cucumber Apostichopus japonicus Selenka. Fish Shellfish Immunol. 2018, 74, 69–75. [Google Scholar] [CrossRef]
- Safari, O.; Paolucci, M. Effect of in vitro selected synbiotics (galactooligosaccharide and mannanoligosaccharide with or without Enterococcus faecalis) on growth performance, immune responses and intestinal microbiota of juvenile narrow clawed crayfish, Astacus leptodactylus leptodactylus Eschscholtz, 1823. Aquacult. Nutr. 2018, 24, 247–259. [Google Scholar]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Ono, S.; Fujieda, T. Comparison of the effects of inosine and inosine monophosphate on growth, immune response, stress resistance and gut morphology of juvenile red sea bream, Pagrus major. Aquaculture 2016, 458, 64–74. [Google Scholar] [CrossRef]
- Cao, H.Z.; Yu, R.H.; Zhang, Y.Y.; Hu, B.Q.; Jian, S.Q.; Wen, C.G.; Kimia, K.; Vikas, K.; Gang, Y. Effects of dietary supplementation with β-glucan and Bacillus subtilis on growth, fillet quality, immune capacity, and antioxidant status of Pengze crucian carp (Carassius auratus var. Pengze). Aquaculture 2019, 508, 106–112. [Google Scholar] [CrossRef]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef] [PubMed]
- María, Á.E. An overview of the immunological defenses in fish skin. Int. Sch. Res. Not. 2012, 2012, 853470. [Google Scholar] [CrossRef]
- Zhong, X.; Liu, M.; Xu, C.; Liu, W.; Abasubong, K.; Li, X. Dietary supplementation of Streptococcus faecalis benefits the feed utilization, antioxidant capability, innate immunity, and disease resistance of blunt snout bream (Megalobrama amblycephala). Fish Physiol. Biochem. 2019, 45, 643–656. [Google Scholar] [CrossRef]
- Jinhwan, P.; Wooju, K.; Wi-Sik, K.; Hyun-Do, J.; Suhee, H. Cloning and expressional analysis of secretory and membrane-bound IgM in rock bream (Oplegnathus fasciatus) under megalocytivirus infection and vaccination. Fish Shellfish Immunol. 2019, 87, 275–285. [Google Scholar]
- Giri, S.S.; Sukumaran, V.; Oviya, M. Potential probiotic Lactobacillus plantarum VSG3 improves the growth, immunity, and disease resistance of tropical freshwater fish, Labeo rohita. Fish Shellfish Immunol. 2013, 34, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Austin, B. Innate immune responses in rainbow trout (Oncorhynchus mykiss, Walbaum) induced by probiotics. Fish Shellfish Immunol 2006, 21, 513–524. [Google Scholar] [CrossRef]
- Low, C.; Wadsworth, S.; Burrells, C.; Secombes, C. Expression of immune genes in turbot (Scophthalmus maximus) fed a nucleotide-supplemented diet. Aquaculture 2003, 221, 23–40. [Google Scholar] [CrossRef]
- Meng, F.X.; Sun, Y.N.; Liu, X.Z.; Wang, J.X.; Xu, T.J.; Wang, R.X. Analysis of C3 suggests three periods of positive selection events and different evolutionary patterns between fish and mammals. PLoS ONE 2012, 7, e37489. [Google Scholar] [CrossRef] [PubMed]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory effects triggered by lactic acid bacteria exopolysaccharides: New insights into molecular interactions with host cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.J.; Li, Z.H.; Ou, M.F.; Wu, X.J.; Qiao, X.L.; Wei, W.; Liu, Y.; Ye, J.M.; Wang, W.N. Hypoimmunity and intestinal bacterial imbalance are closely associated with blue body syndrome in cultured Penaeus vannamei. Aquaculture 2020, 522, 735118. [Google Scholar] [CrossRef]
- Lee, S.; Katya, K.; Park, Y.; Won, S.; Seong, M.; Hamidoghli, A.; Bai, S.C. Comparative evaluation of dietary probiotics Bacillus subtilis WB60 and Lactobacillus plantarum KCTC3928 on the growth performance, immunological parameters, gut morphology and disease resistance in Japanese eel, Anguilla japonica. Fish Shellfish Immunol. 2017, 61, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, J.G.; Tattoli, I.; Girardin, S.E. The intestinal epithelial barrier: How to distinguish between the microbial flora and pathogens. Semin. Immunol. 2007, 19, 106–115. [Google Scholar] [CrossRef]
- Galeano, J.A.C.; Herrera, A.L.; Suescún, J.P. The probiotic Enterococcus faecium modifies the intestinal morphometric parameters in weaning piglets. Rev. Fac. Nac. Agron. Medellín 2016, 69, 7803–7811. [Google Scholar] [CrossRef]
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, J.S.; Baker, T.M.R.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Blander, J.M.; Longman, R.S.; Iliev, I.D.; Sonnenberg, G.F.; Artis, D. Regulation of inflammation by microbiota interactions with the host. Nat. Immunol. 2017, 18, 851–860. [Google Scholar] [CrossRef]
- Emilie, C.; Yannick, G.; Kevin, M.; Bénédicte, L.; David, P.; Fabien, P.; Florian, N.; Denis, S. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.B.; Gatesoupe, F.J.; Li, T.T.; Wang, X.H.; Zhang, Q.Q.; Feng, D.Y.; Feng, Y.Q.; Chen, H.; Li, A.H. Significant improvement of intestinal microbiota of gibel carp (Carassius auratus gibelio) after traditional Chinese medicine feeding. J. Appl. Microbiol. 2018, 124, 829–841. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Miao, S.Y.; Zhao, C.Z.; Zhu, J.Y.; Hu, J.T.; Dong, X.J.; Sun, L.S. Dietary soybean meal affects intestinal homoeostasis by altering the microbiota, morphology and inflammatory cytokine gene expression in northern snakehead. Sci. Rep. 2018, 8, 113–122. [Google Scholar] [CrossRef]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; Sinderen, D. Genomics of actinobacteria: Tracing the evolutionary history of an ancient phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef]
- Lu, W.H.; Chen, H.; Zhou, Y.; Li, Y.J.; Wang, X.D.; Huag, C.G. Identification and drug sensitive test of the pathogen in Acinetobacter disease from hybrid crucian carp (Carassius auratus gibelio ♀ × Cyprinus carpio ♂). Fish. Sci. 2010, 29, 156–161. [Google Scholar]
- Tsuchiya, C.; Sakata, T.; Sugita, H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 2008, 46, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Baños, A.; Ariza, J.J.; Nuñez, C.; Gil-Martínez, L.; García-López, J.D.; Martínez-Bueno, M.; Valdivia, E. Effects of Enterococcus faecalis UGRA10 and the enterocin AS-48 against the fish pathogen Lactococcus garvieae. Studies in vitro and in vivo. Food Microbiol. 2019, 77, 69–77. [Google Scholar] [CrossRef]
- Hao, K.; Wu, Z.Q.; Li, D.L.; Yu, X.B.; Wang, G.X.; Ling, F. Effects of Dietary Administration of Shewanella xiamenensis A-1, Aeromonas veronii A-7, and Bacillus subtilis, Single or Combined, on the Grass Carp (Ctenopharyngodon idella) Intestinal Microbiota. Probiotics Antimicro. 2017, 9, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X.; Kang, Y.H.; Zhan, S.Z.; Ze, L.; Jin, S.N.; Chen, C.; Zhang, L.; Shen, J.Y.; Wang, C.F.; Wang, G.Q.; et al. Effect of Bacillus velezensis on Aeromonas veronii -Induced Intestinal Mucosal Barrier Function Damage and Inflammation in Crucian Carp (Carassius auratus). Front. Microbiol. 2019, 10, 2663. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′–3′) | GenBank Number |
|---|---|---|
| β-action-F | GATGATGAAATTGCCGCACTG | AB039726 [24] |
| β-action-R | ACCGACCATGACGCCCTGATGT | |
| IL-1β-F | GCGCTGCTCAACTTCATCTTG | AJ249137 [24] |
| IL-1β-R | GTGACACATTAAGCGGCTTCA | |
| C3-F | AGTGAAATGGTGGAAGCAGAAAG | KF110786 [25] |
| C3-R | TACGTATACCGAGACATCGAAGG | |
| IgM-F | GTGGAACTTGATGCCCCAAT | MK272741.1 |
| IgM-R | CATCAGCAAGCCAAGACACAA |
| Parameters | CG | EF1 | EF2 | EF3 |
|---|---|---|---|---|
| Initial weight (g) | 47.67 ± 0.62 | 48.60 ± 0.15 | 48.37 ± 0.52 | 48.01 ± 0.51 |
| Weight at sampling (g) | 64.57 ± 0.93 a | 68.44 ± 1.05 b | 69.45 ± 0.96 b | 67.38 ± 2.74 ab |
| WGR (%) | 35.47 ± 1.11 a | 40.83 ± 1.78 b | 43.59 ± 0.58 b | 40.31 ± 4.26 ab |
| SGR (%/d) | 1.08 ± 0.03 a | 1.22 ± 0.04 b | 1.29 ± 0.02 b | 1.21 ± 0.11 b |
| Survival rate (%) | 96.00 ± 2.00 | 96.67 ± 2.31 | 98.00 ± 2.00 | 96.67 ± 3.01 |
| (CF) (g/cm3) | 2.29 ± 0.16 | 2.45 ± 0.49 | 2.41 ± 0.13 | 2.21 ± 0.18 |
| Sample | OTUs | Chao 1 | Shannon |
|---|---|---|---|
| CG | 824.67 ± 56.52 a | 857.24 ± 46.64 a | 5.40 ± 0.31 |
| EF1 | 842.33 ± 23.51 a | 911.79 ± 40.69 a | 5.12 ± 0.83 |
| EF2 | 1025.67 ± 81.24 b | 1190.46 ± 102.67 b | 5.75 ± 0.09 |
| EF3 | 878.67 ± 79.74 a | 882.85 ± 41.28 a | 5.61 ± 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Li, Y.; Xue, M.; Xiao, Z.; Fan, Y.; Zeng, L.; Zhou, Y. Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus). Fishes 2022, 7, 18. https://doi.org/10.3390/fishes7010018
Xu Y, Li Y, Xue M, Xiao Z, Fan Y, Zeng L, Zhou Y. Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus). Fishes. 2022; 7(1):18. https://doi.org/10.3390/fishes7010018
Chicago/Turabian StyleXu, Yan, Yiqun Li, Mingyang Xue, Zidong Xiao, Yuding Fan, Lingbing Zeng, and Yong Zhou. 2022. "Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus)" Fishes 7, no. 1: 18. https://doi.org/10.3390/fishes7010018
APA StyleXu, Y., Li, Y., Xue, M., Xiao, Z., Fan, Y., Zeng, L., & Zhou, Y. (2022). Effects of Dietary Enterococcus faecalis YFI-G720 on the Growth, Immunity, Serum Biochemical, Intestinal Morphology, Intestinal Microbiota, and Disease Resistance of Crucian Carp (Carassius auratus). Fishes, 7(1), 18. https://doi.org/10.3390/fishes7010018