
The Innate Immune Response to Infection by Polyascus gregaria in the Male Chinese Mitten Crab (Eriocheir sinensis), Revealed by Proteomic Analysis

Abstract
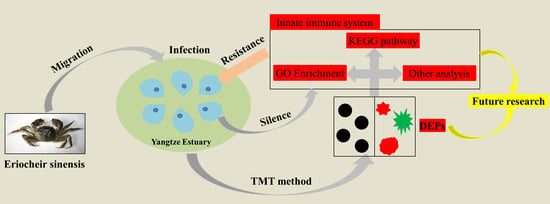
1. Introduction
2. Materials and Methods
2.1. Sample Site and Crabs
2.2. Experimental Protein Preparation
2.3. TMT Protein Labelling and HPLC Fractionation
2.4. LC-MS/MS Analysis
2.5. Data Analysis
2.6. Enrichment of Pathways Analysis
2.7. Enrichment of Protein Domain Analysis
3. Results
3.1. Identification and Quantitative Protein Profiling
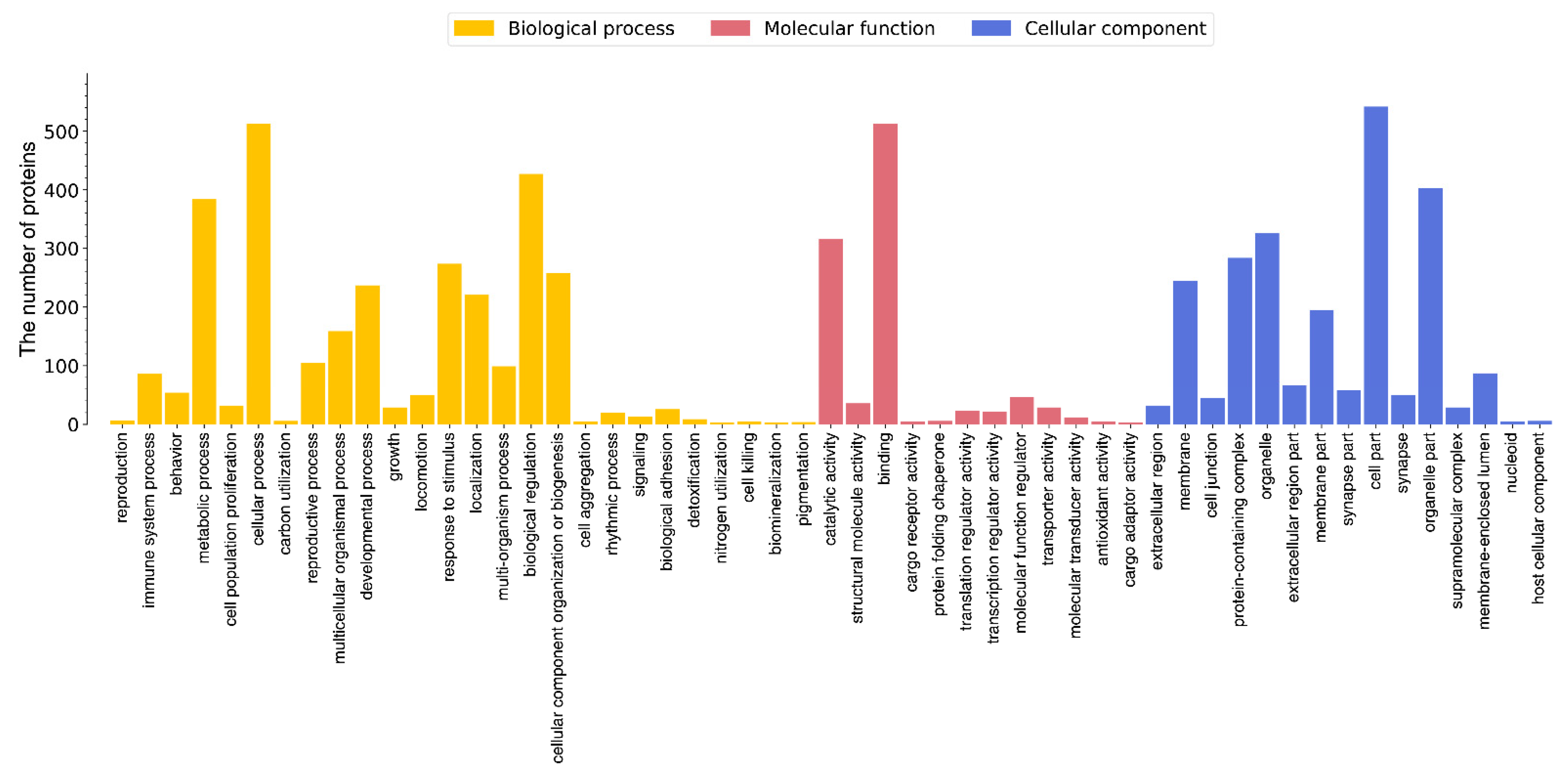
3.2. GO Enrichment
3.3. KEGG Analysis with DEPs
3.4. Enrichment of Protein Domain
3.5. Subcellular Location of DEPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Wang, M.; Ge, J.; Yu, J.; Su, S.; Li, J.; Tang, Y. Molecular insights into information processing and developmental and immune regulation of Eriocheir sinensis megalopa under hyposaline stress. Genomics 2020, 112, 4647–4656. [Google Scholar] [CrossRef] [PubMed]
- Gillard, M.; Thiébaut, G.; Deleu, C.; Leroy, B. Present and future distribution of three aquatic plants taxa across the world: Decrease in native and increase in invasive ranges. Biol. Invasions 2017, 19, 2159–2170. [Google Scholar] [CrossRef]
- Qiu, G.; Xiong, L.; Liu, Z.; Yan, Y.; Shen, H. A first generation microsatellite-based linkage map of the Chinese mitten crab Eriocheir sinensis and its application in quantitative trait loci (QTL) detection. Aquaculture 2016, 451, 223–231. [Google Scholar] [CrossRef]
- Bonami, J.; Zhang, S. Viral diseases in commercially exploited crabs: A review. J. Invertebr. Pathol. 2011, 106, 6–17. [Google Scholar] [CrossRef]
- Miroliubov, A.; Borisenko, I.; Nesterenko, M.; Korn, O.; Lianguzova, A.; Ilyutkin, S.; Lapshin, N.; Dobrovolskij, A.A. Muscular system in the interna of Polyascus polygenea and Sacculina pilosella (Cirripedia: Rhizocephala: Sacculinidae). Invertebr. Zool. 2019, 16, 48–56. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Q.; Zhang, T.; Li, Z.; Liu, J. Effects of water temperature on growth, feeding and molting of juvenile Chinese mitten crab Eriocheir sinensis. Aquaculture 2017, 468, 169–174. [Google Scholar] [CrossRef]
- Veillet, A. Recherches Sur le Parasitisme des Crabes et des Galathées par les Khizocéphales et les Épicarides; Annales de l’Institut Oceanographique Monaco: Monte-Carlo, Monaco, 1945; Volume 22, pp. 193–341. [Google Scholar]
- Wang, J.; Yang, B.; Wang, W.; Song, X.; Jiang, Q.; Qiu, L.; Wang, L.; Song, L. The enhanced immune protection in Chinese mitten crab Eriocheir sinensis against the second exposure to bacteria Aeromonas hydrophila. Front. Immunol. 2019, 10, 2041. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Bao, J.; Xing, Y.; Feng, C.; Li, X.; Chen, Q. Proteomic analysis of the hemolymph after Metschnikowia bicuspidata infection in the Chinese mitten crab Eriocheir sinensis. Front. Immunol. 2021, 12, 659–723. [Google Scholar]
- Wang, W.; Chen, J. Ultrastructural study on a novel microsporidian, Endoreticulatus eriocheir sp. nov. (Microsporidia, Encephalitozoonidae), parasite of Chinese mitten crab, Eriocheir sinensis (Crustacea, Decapoda). J. Invertebr. Pathol. 2007, 94, 77–83. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, L.; Zhou, J.; Zou, J.; Fan, L. New insights into the immune regulation and tissue repair of Litopenaeus vannamei during temperature fluctuation using TMT-based proteomics. Fish Shellfish Immunol. 2020, 106, 975–981. [Google Scholar] [CrossRef]
- Sousa, L.G.; Cuartas, E.I.; Petriella, A.M. Fine structural analysis of the epithelial cells in the hepatopancreas of Palaemonetes argentinus (Crustacea, Decapoda, Caridea) in intermoult. Biocell 2005, 29, 25. [Google Scholar] [CrossRef]
- Gao, T.; Xu, Y.; Wang, K.; Deng, Y.; Yang, Y.; Lu, Q.; Pan, J.; Xu, Z. Comparative LC-MS based non-targeted metabolite profiling of the Chinese mitten crab Eriocheir sinensis suffering from hepatopancreatic necrosis disease (HPND). Aquaculture 2018, 491, 338–345. [Google Scholar] [CrossRef]
- Li, X.; Cui, Z.; Liu, Y.; Song, C.; Shi, G. Transcriptome analysis and discovery of genes involved in immune pathways from hepatopancreas of microbial challenged mitten crab Eriocheir sinensis. PLoS ONE 2013, 8, e68233. [Google Scholar] [CrossRef]
- Li, S.; Jia, Z.; Li, X.; Geng, X.; Sun, J. Calmodulin is a stress and immune response gene in Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2014, 40, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Roux, M.; Pain, A.; Klimpel, K.; Dhar, A. The lipopolysaccharide and β-1, 3-glucan binding protein gene is upregulated in white spot virus-infected shrimp (Penaeus stylirostris). J. Virol. 2002, 76, 7140–7149. [Google Scholar] [CrossRef]
- Ried, C.; Wahl, C.; Miethke, T.; Wellnhofer, G.; Landgraf, C.; Schneider-Mergener, J.; Adolf, H. High affinity endotoxin-binding and neutralizing peptides based on the crystal structure of recombinant Limulus anti-lipopolysaccharide factor. J. Biol. Chem. 1996, 271, 28120–28127. [Google Scholar] [CrossRef] [PubMed]
- Gross, P.; Bartlett, T.; Browdy, C.; Chapman, R.; Warr, G. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Glenner, H.; Lützen, J.; Takahashi, T. Molecular and morphological evidence for a monophyletic clade of asexually reproducing Rhizocephala: Polyascus, new genus (Cirripedia). J. Crustacean Biol. 2003, 23, 548–557. [Google Scholar] [CrossRef]
- Jorgen, L.; Tohru, T. Sacculina polygenea, a new species of rhizocephalan (Cirripedia: Rhizocephala) from Japan, parasitic on the intertidal crab Hemigrapsus sanguineus (De Haan, 1835) (Decapoda: Brachyura: Grapsidae). Crustacean Res. 1997, 26, 103–108. [Google Scholar]
- Høeg, J.; Lützen, J. Crustacea Rhizocephala; Christiansen, M.E., Ed.; Norwegian University Press: Oslo, Norway, 1985; Volume 6, pp. 1–92. [Google Scholar]
- Rowley, A.; Davies, C.; Malkin, S.; Bryan, C.; Thomas, J.; Batista, F.; Coates, C. Prevalence and histopathology of the parasitic barnacle, Sacculina carcini in shore crabs, Carcinus maenas. J. Invertebr. Pathol. 2020, 171, 107338. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.; Wu, Y.; Tung, T.; Wang, G.; Toullec, J.; Liu, S.; Huang, W.; Lee, C. Metabolic effects of parasitisation by the barnacle Polyascus plana (Cirripedia: Rhizocephala: Sacculinidae) on a grapsid host, Metopograpsus thukuhar. Dis. Aquat. Org. 2016, 119, 199–206. [Google Scholar] [CrossRef]
- Waiho, K.; Fazhan, H.; Glenner, H.; Ikhwanuddin, M. Infestation of parasitic rhizocephalan barnacles Sacculina beauforti (Cirripedia, Rhizocephala) in edible mud crab Scylla Olivacea. PeerJ 2017, 5, e3419. [Google Scholar] [CrossRef]
- Tsuchida, K.; Lützen, J.; Nishida, M. Sympatric three-species infection by Sacculina parasites (Cirripedia: Rhizocephala: Sacculinidae) of an intertidal grapsoid crab. J. Crustacean Biol. 2006, 26, 474–479. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Jensen, T. The effect of Sacculina carcini infections on the fouling, burying behaviour and condition of the shore crab, Carcinus maenas. Ophelia 2006, 2, 270–275. [Google Scholar]
- Heg, J.T. The biology and life cycle of the Rhizocephala (Cirripedia). J. Mar. Biol. Assoc. 1995, 75, 517–550. [Google Scholar] [CrossRef]
- Kristensen, T.; Nielsen, A.; Jørgensen, A.; Mouritsen, K.; Glenner, H.; Christensen, J.; Lützen, J.; Høeg, J. The selective advantage of host feminization: A case study of the green crab Carcinus maenas and the parasitic barnacle Sacculina carcini. Mar. Biol. 2012, 159, 2015–2023. [Google Scholar] [CrossRef]
- Adam, P.; Rowley, A.F. Tissue changes in the shore crab Carcinus maenas as a result of infection by the parasitic barnacle Sacculina carcini. Dis. Aquat. Org. 2008, 80, 75–79. [Google Scholar]
- Lützen, J. Growth, reproduction, and life span in Sacculina carcini Thompson (Cirripedia: Rhizocephala) in the Isefjord, Denmark. Sarsia 1984, 69, 91–105. [Google Scholar] [CrossRef]
- Trédez, F.; Rabet, N.; Bellec, L.; Audebert, F. Synchronism of naupliar development of Sacculina carcini Thompson, 1836 (Pancrustacea, Rhizocephala) revealed by precise monitoring. Helgol. Mar. Res. 2016, 70, 26. [Google Scholar] [CrossRef]
- Waiho, K.; Fazhan, H.; Zhang, Y.; Afiqah-Aleng, N.; Moh, J.; Ikhwanuddin, M.; Hassan, M.; Norainy, M.; Ma, H. Gonadal transcriptomic analysis of the mud crab Scylla olivacea infected with rhizocephalan parasite Sacculina beauforti. Genomics 2020, 112, 2959–2969. [Google Scholar] [CrossRef]
- Tang, B.; Wang, Q.; Chen, L.; Yang, S. Features of an intersex Chinese mitten crab, Eriocheir Japonica sinensis (decapoda, brachyura). Freshw. Fish 2005, 78, 371–377. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Aratake, H. Morphological modifications caused by Sacculina polygenea in Hemigrapsus sanguineus (De Haan) (Brachyura: Grapsidae). Crustacean Res. 1997, 26, 125–145. [Google Scholar] [CrossRef]
- Iwanaga, S.; Lee, B.-L. Recent advances in the innate immunity of invertebrate animals. BMB Rep. 2005, 38, 128–150. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Lee, B.L.; Söderhäll, K. Cell-mediated immunity in arthropods: Hematopoiesis, coagulation, melanization and opsonization. Immunobiology 2006, 211, 213–236. [Google Scholar] [CrossRef]
- Kong, P.; Wang, L.; Zhang, H.; Zhou, Z.; Qiu, L.; Gai, Y.; Song, L. Two novel secreted ferritins involved in immune defense of Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2010, 28, 604–612. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, A.; Ao, W.; Wang, Z.; Yuan, J.; Song, Q.; Wei, D.; Ye, H. Proteomic analysis of serum proteins from HIV/AIDS patients with Talaromyces marneffei infection by TMT labeling-based quantitative proteomics. Clin. Proteom. 2018, 15, 40. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Jiang, Z. Immune induction identified by TMT proteomics analysis in Fusobacterium nucleatum autoinducer-2 treated macrophages. Expert Rev. Proteom. 2020, 17, 175–185. [Google Scholar] [CrossRef]
- Du, N.S. Crustaceology; Scence Press: Beijing, China, 1987; Volume 1. [Google Scholar]
- Yan, B.; Liu, X.; Zhou, Y.; Zhang, M.; Fang, P.; Jiang, M.; Yuan, R.; Hu, X.; Gao, G.; Xue, R.; et al. Transcriptomic analysis reveals that hepatopancreatic necrosis disease in Eriocheir sinensis (Chinese mitten crabs) may be the result of autophagy and apoptosis. Aquaculture 2020, 515, 734579. [Google Scholar] [CrossRef]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef]
- Sun, M.X.; Huang, L.; Wang, R.; Yu, Y.L.; Li, C.; Li, P.P.; Hu, X.C.; Hao, H.P.; Ishag, H.A.; Mao, X. Porcine reproductive and respiratory syndrome virus induces autophagy to promote virus replication. Autophagy 2012, 8, 1434–1447. [Google Scholar] [CrossRef]
- Galluzzi, L.; Pedro, J.; Levine, B.; Green, D.; Kroemer, G. Pharmacological modulation of autophagy: Therapeutic potential and persisting obstacles. Nat. Rev. Drug Discov. 2017, 16, 487–511. [Google Scholar] [CrossRef]
- Maruyama, T.; Noda, N.N. Autophagy-regulating protease Atg4: Structure, function, regulation and inhibition. J. Antibiot. 2018, 71, 72–78. [Google Scholar] [CrossRef]
- Suzuki, K.; Kirisako, T.; Kamada, Y.; Mizushima, N.; Noda, T.; Ohsumi, Y. The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J. 2001, 20, 5971–5981. [Google Scholar] [CrossRef]
- Suzuki, K.; Kubota, Y.; Sekito, T.; Ohsumi, Y. Hierarchy of Atg proteins in pre-autophagosomal structure organization. Clarendon Press 2007, 12, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, J.; Zhang, X.; Wei, S.; Huang, X.; Huang, Y.; Wei, J.; Qin, Q. Fish Autophagy Protein 5 Exerts Negative Regulation on Antiviral Immune Response against Iridovirus and Nodavirus. Front. Immunol. 2019, 10, 517. [Google Scholar] [CrossRef]
- Jin, M.; Klionsky, D.J. Transcriptional regulation of ATG9 by the Pho23-Rpd3 complex modulates the frequency of autophagosome formation. Autophagy 2014, 10, 1681–1682. [Google Scholar] [CrossRef] [PubMed]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef]
- Luzio, J.P.; Pryor, P.R.; Bright, N.A. Lysosomes: Fusion and function. Nat. Rev. Mol. Cell Biol. 2007, 8, 622–632. [Google Scholar] [CrossRef]
- Hou, L.; Zhou, H.; Wan, H.; Liu, Z.; Wang, L.; Cheng, Y.; Wu, X.; Gu, W.; Wang, W.; Meng, Q. TMT-based quantitative proteomic analysis of Eriocheir sinensis hemocytes and thoracic ganglion during Spiroplasma eriocheiris infection. Fish Shellfish Immunol. 2020, 96, 126–137. [Google Scholar] [CrossRef]
- Hipolito, V.E.B.; Ospina-Escobar, E.; Botelho, R.J. Lysosome remodelling and adaptation during phagocyte activation. Cell Microbiol. 2018, 20, e12824. [Google Scholar] [CrossRef]
- Raposo, G.; Fevrier, B.; Stoorvogel, W.; Marks, M. Lysosomes related organelles: A view from immunity and pigmentation. Cell Struct. Funct. 2002, 27, 443–456. [Google Scholar] [CrossRef][Green Version]
- Labrousse, A.M.; Meunier, E.; Record, J.; Labernadie, A.; Beduer, A.; Vieu, C.; Safta, T.; Maridonneau-Parini, I. Frustrated phagocytosis on micro-patterned immune complexes to characterize lysosome movements in live macrophages. Front. Immunol. 2011, 2, 51. [Google Scholar] [CrossRef]
- Colbert, J.; Matthews, S.; Miller, G.; Watts, C. Diverse regulatory roles for lysosomal proteases in the immune response. Eur. J. Immunol. 2009, 39, 2955–2965. [Google Scholar] [CrossRef]
- Turk, B.; Turk, D.; Turk, V. Lysosomal cysteine proteases: More than scavengers. Biochim. Biophys. Acta 2000, 1477, 98–111. [Google Scholar] [CrossRef]
- Turk, B.; Stoka, V.; Rozman-Pungercar, J.; Cirman, T.; Droga-Mazovec, G.; Oreic, K.; Turk, V. Apoptotic Pathways: Involvement of lysosomal proteases. Biol. Chem. 2002, 383, 1035–1044. [Google Scholar] [CrossRef]
- Keegan, P.M.; Wilder, C.L.; Platt, M.O. Tumor necrosis factor alpha stimulates cathepsin K and V activity via juxtacrine monocyte-endothelial cell signaling and JNK activation. Mol. Cell Biochem. 2012, 367, 65–72. [Google Scholar] [CrossRef]
- Tsukuba, T.; Yanagawa, M.; Kadowaki, T.; Takii, R.; Okamoto, Y.; Sakai, E.; Okamoto, K.; Yamaoto, K. Cathepsin E deficiency impairs autophagic proteolysis in macrophages. PLoS ONE 2013, 8, e82415. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, Y.X.; Liu, J.; Yang, C.; Feng, Q.; Xu, W. A regulatory pathway, Ecdysone-transcription factor Relish-Cathepsin L, is involved in insect fat body dissociation. PLoS Genet. 2013, 9, e1003273. [Google Scholar] [CrossRef]
- Ning, M.; Yuan, M.; Liu, M.; Gao, Q.; Wei, P.; Gu, W.; Wang, W.; Meng, Q. Characterization of cathepsin D from Eriocheir sinensis involved in Spiroplasma eriocheiris infection. Dev. Comp. Immunol. 2018, 86, 1–8. [Google Scholar] [CrossRef]
- Tulone, C.; Uchiyama, Y.; Novelli, M.; Grosvenor, N.; Softig, P.; Chain, B. Haematopoietic development and immunological function in the absence of cathepsin D. BMC Immunol. 2007, 8, 22. [Google Scholar] [CrossRef]
- Li, W.; Jin, X.; He, L.; Jiang, H.; Gong, Y.; Xie, Y.; Wang, J. Molecular cloning, characterization, expression and activity analysis of cathepsin L in Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2010, 29, 1010–1018. [Google Scholar] [CrossRef]
- Qiu, L.; Jiang, S.; Huang, J.; Wang, W.; Zhang, D.; Wu, Q.; Yang, K. Molecular cloning and mRNA expression of cathepsin C gene in black tiger shrimp (Penaeus monodon). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 150, 320–325. [Google Scholar] [CrossRef]
- Zhao, Z.; Yin, Z.; Weng, S.; Guan, H.; Li, S.; Xing, K.; Chan, S.; He, J. Profiling of differentially expressed genes in hepatopancreas of white spot syndrome virus-resistant shrimp (Litopenaeus vannamei) by suppression subtractive hybridisation. Fish Shellfish Immunol. 2007, 22, 520–534. [Google Scholar] [CrossRef]
- Shi, G.P.; Bryant, R.A.R.; Riese, R.; Verhelst, S.; Driessen, C.; Li, Z.; Bromme, D.; Ploegh, H.; Chapman, H. Role for Cathepsin F in invariant chain processing and Major Histocompatibility Complex Class II peptide loading by Macrophages. J. Exp. Med. 2000, 191, 1177–1186. [Google Scholar] [CrossRef]
- Turk, B.; Turk, D.; Salvesen, G. Regulating cysteine protease activity: Essential role of protease inhibitors as guardians and regulators. Curr. Pharm. Des. 2002, 8, 1623–1637. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, Y.; Jiang, H.; Xu, S.; Zhao, X.; Wang, J. Antibacterial activity of serine protease inhibitor 1 from kuruma shrimp Marsupenaeus japonicus. Dev. Comp. Immunol. 2014, 44, 261–269. [Google Scholar] [CrossRef]
- Rimphanitchayakit, V.; Tassanakajon, A. Structure and function of invertebrate Kazal-type serine proteinase inhibitors. Dev. Comp. Immunol. 2009, 34, 377–386. [Google Scholar] [CrossRef]
- Tang, H.; Kambris, Z.; Lemaitre, B.; Hashimoto, C. A serpin that regulates immune melanization in the respiratory system of Drosophila. Dev. Cell 2008, 15, 617–626. [Google Scholar] [CrossRef]
- Wetsaphan, N.; Rimphanitchayakit, V.; Tassanakajon, A.; Somboonwiwat, K. PmSERPIN3 from black tiger shrimp Penaeus monodon is capable of controlling the proPO system. Dev. Comp. Immunol. 2013, 41, 110–119. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Z.; Yang, J.; Gai, Y.; Zhou, Z.; Wang, L.; Yue, F.; Song, L. Identification and characterization of a serine protease inhibitor Esserpin from the Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2013, 34, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Söderhäll, K.; Cerenius, L. Role of the prophenoloxidase-activating system in invertebrate immunity. Curr. Opin. Immunol. 1998, 10, 23–28. [Google Scholar] [CrossRef]
- Park, D.; Shin, S.; Hong, S.; Park, H. Immunological detection of Serpin in the Fall webworm, Hyphantria cunea and its inhibitory activity on the prophenoloxidase system. Mol. Cells 2000, 10, 186–192. [Google Scholar] [CrossRef]
- Koskella, B. Resistance gained, resistance lost: An explanation for host-parasite coexistence. PLoS Biol. 2018, 16, e3000013. [Google Scholar] [CrossRef]
- Jensen, A.; Schneider, M.; Høeg, J.; Glenner, H.; Lützen, J. Variation in juvenile stages and success of male acquisition in Danish and French populations of the parasitic barnacle Sacculina carcini (Cirripedia: Rhizocephala) parasitizing the shore crab Carcinus maenas. Mar. Biol. Res. 2019, 15, 191–203. [Google Scholar] [CrossRef]
- Han, F.; Zhang, X. Characterization of a ras-related nuclear protein (Ran protein) up-regulated in shrimp antiviral immunity. Fish Shellfish Immunol. 2007, 23, 937–944. [Google Scholar] [CrossRef]
- Scheele, J.S.; Marks, R.E.; Boss, G.R. Signaling by small GTPases in the immune system. Immunol. Rev. 2007, 218, 92–101. [Google Scholar] [CrossRef]
- Patin, E.C.; Orr, S.J.; Schaible, U.E. Macrophage Inducible C-Type Lectin as a Multifunctional Player in Immunity. Front. Immunol. 2017, 8, 861. [Google Scholar] [CrossRef]
- Liyanage, D.; Omeka, W.; Yang, H.; Lim, C.; Kwon, H.; Choi, C.; Lee, J. Expression profiling, immune functions, and molecular characteristics of the tetraspanin molecule CD63 from Amphiprion clarkii. Dev. Comp. Immunol. 2021, 123, 104168. [Google Scholar] [CrossRef]
- Wang, Y.; Su, J.; Yuan, B.; Fu, D.; Niu, Y.; Yue, D. The role of C1QBP in CSF-1-dependent PKCζ activation and macrophage migration. Exp. Cell Res. 2018, 362, 11–16. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Sample ID | CL (mm) | CW (mm) | CT (mm) | Weight (g) | PN |
|---|---|---|---|---|---|---|
| non-parasitized | ESC1 | 54.92 | 51.57 | 26.43 | 76.06 | 0 |
| ESC2 | 57.22 | 53.34 | 26.08 | 85.34 | 0 | |
| ESC3 | 56.19 | 56.19 | 29.34 | 80.12 | 0 | |
| ESC4 | 56.87 | 51.51 | 37.97 | 89.42 | 0 | |
| ESC5 | 57.57 | 55.45 | 31.76 | 98.87 | 0 | |
| ESC6 | 57.24 | 56.32 | 27.76 | 105.42 | 0 | |
| Parasitized | ESI1 | 61.63 | 58.25 | 28.84 | 102.19 | 21 |
| ESI2 | 49.83 | 48.06 | 23.41 | 60.00 | 17 | |
| ESI3 | 54.30 | 53.49 | 26.58 | 80.94 | 27 | |
| ESI4 | 46.52 | 47.07 | 22.20 | 46.96 | 21 | |
| ESI5 | 45.74 | 43.73 | 20.94 | 46.46 | 17 | |
| ESI6 | 61.11 | 59.53 | 28.02 | 106.66 | 24 |
| Protein ID | Name | Coverage | 1.2-Fold | p Value |
|---|---|---|---|---|
| Unigene003240.p1 | Cell division control protein 42 | 17 | 2.63 | 0.0000 |
| Unigene000553.p1 | E3 ubiquitin-protein ligase HERC4 | 4 | 1.96 | 0.0000 |
| Unigene003237.p1 | Ras-related protein Rab-7A | 26 | 1.30 | 0.0001 |
| Unigene005035.p1 | UTP--glucose-1-phosphate uridylyltransferase | 22 | 1.30 | 0.0002 |
| Unigene000566.p1 | Ras-related GTP-binding protein A/B | 17 | 1.34 | 0.0005 |
| Unigene004773.p1 | Chaperonin GroEL | 38 | 1.73 | 0.0007 |
| Unigene002294.p1 | Ras-related protein Rab-30 | 17 | 2.04 | 0.0010 |
| Unigene000261.p1 | Dynactin 1 | 2 | 1.71 | 0.0032 |
| Unigene003260.p1 | Ras-related protein Rab-6A | 39 | 2.02 | 0.0033 |
| Unigene004729.p1 | High mobility group protein B1 | 7 | 1.41 | 0.0034 |
| Unigene002219.p1 | Ras homolog gene family, member A | 29 | 1.56 | 0.0039 |
| Unigene004782.p1 | Calreticulin | 33 | 2.81 | 0.0041 |
| Unigene006723.p1 | Ras-related protein Rab-1A | 43 | 1.85 | 0.0044 |
| Unigene000585.p1 | Cathepsin F | 4 | 1.40 | 0.0045 |
| Unigene000091.p1 | Kindlin 2 | 8 | 1.39 | 0.0055 |
| Unigene007453.p1 | Small subunit ribosomal protein s6e | 28 | 2.07 | 0.0059 |
| Unigene000740.p1 | Ras-related protein Rab-2A | 50 | 1.69 | 0.0073 |
| Unigene004596.p1 | Protein disulfide-isomerase A1 | 49 | 1.71 | 0.0115 |
| Unigene004078.p2 | Tubulin-specific chaperone A | 16 | 2.51 | 0.0124 |
| Unigene001138.p1 | Matrix metalloproteinase-14 | 3 | 1.44 | 0.0150 |
| Unigene000336.p1 | GTPase KRas | 12 | 2.06 | 0.0167 |
| Unigene007460.p1 | Small subunit ribosomal protein s3e | 53 | 1.38 | 0.0250 |
| Unigene000055.p1 | Laminin, alpha 3/5 | 16 | 1.26 | 0.0270 |
| Unigene006004.p1 | Vacuolar protein-sorting-associated protein 4 | 8 | 1.37 | 0.0294 |
| Unigene005905.p1 | Complement component 1 Q subcomponent-binding protein, mitochondrial | 15 | 2.33 | 0.0297 |
| Unigene003829.p1 | Protein scribble | 13 | 1.31 | 0.0304 |
| Unigene001410.p1 | Fascin 1 | 4 | 1.83 | 0.0342 |
| Unigene006225.p1 | Glutathione S-transferase | 36 | 2.06 | 0.0006 |
| Unigene000029.p1 | P-type Ca2+ transporter type 2B | 9 | 1.48 | 0.0178 |
| Unigene000668.p1 | Filamin | 7 | 1.36 | 0.0038 |
| Unigene001686.p1 | 1,4-alpha-glucan branching enzyme | 27 | 1.35 | 0.0004 |
| Unigene001793.p1 | Neural cell adhesion molecule | 4 | 2.30 | 0.0408 |
| Unigene005645.p1 | AN1-type zinc finger protein 1 | 4 | 4.77 | 0.0005 |
| Unigene003973.p1 | KRAB domain-containing zinc finger protein | 2 | 4.82 | 0.0082 |
| Unigene000005.p1 | Integrin alpha 7 | 8 | 0.72 | 0.0003 |
| Unigene000042.p1 | Lysosomal alpha-glucosidase | 9 | 0.32 | 0.0021 |
| Unigene000381.p1 | Integrin beta 1 | 11 | 0.51 | 0.0069 |
| Unigene000396.p1 | Cysteine protease ATG4 | 7 | 0.78 | 0.0108 |
| Unigene000417.p1 | Serpin B | 67 | 0.49 | 0.0147 |
| Unigene000427.p1 | Glycerol-3-phosphate O-acyltransferase 1/2 | 4 | 0.60 | 0.0044 |
| Unigene000486.p1 | Hexosaminidase | 34 | 0.49 | 0.0211 |
| Unigene000506.p1 | Vacuolar protein sorting-associated protein 35 | 21 | 0.79 | 0.0108 |
| Unigene000515.p1 | Lectin, mannose-binding 2 | 13 | 0.73 | 0.0267 |
| Unigene000557.p1 | CD63 antigen | 11 | 0.53 | 0.0214 |
| Unigene000614.p1 | Dipeptidyl-peptidase 4 | 18 | 0.50 | 0.0150 |
| Unigene000677.p1 | Integrin alpha 8 | 17 | 0.71 | 0.0099 |
| Unigene000679.p1 | Protein kinase D | 6 | 0.66 | 0.0186 |
| Unigene000969.p1 | Spondin-1 | 2 | 0.70 | 0.0418 |
| Unigene001066.p1 | Guanine nucleotide exchange factor VAV | 4 | 0.71 | 0.0200 |
| Unigene001121.p1 | Ubiquitin-associated and SH3 domain-containing protein | 5 | 0.55 | 0.0354 |
| Unigene001159.p1 | Protein spaetzle | 10 | 0.55 | 0.0003 |
| Unigene001277.p1 | Beta-mannosidase | 11 | 0.55 | 0.0295 |
| Unigene001291.p1 | Vacuolar protein sorting-associated protein 18 | 2 | 0.63 | 0.0310 |
| Unigene001385.p1 | Beta-mannosidase | 12 | 0.58 | 0.0441 |
| Unigene001395.p1 | Serine/threonine-protein phosphatase 4 catalytic subunit | 5 | 0.55 | 0.0015 |
| Unigene001452.p1 | Neurobeachin | 9 | 0.73 | 0.0024 |
| Unigene001494.p1 | Integrin beta 1 | 3 | 0.47 | 0.0449 |
| Unigene001509.p1 | Serine incorporator 1/3 | 1 | 0.48 | 0.0116 |
| Unigene001818.p1 | Presenilin 1 | 2 | 0.65 | 0.0055 |
| Unigene001922.p1 | Autophagy-related protein 9 | 2 | 0.66 | 0.0129 |
| Unigene002296.p1 | Glutamyl aminopeptidase | 18 | 0.74 | 0.0103 |
| Unigene003160.p1 | Cathepsin D | 26 | 0.68 | 0.0046 |
| Unigene003249.p1 | Scavenger receptor class B, member 1 | 11 | 0.57 | 0.0020 |
| Unigene003625.p1 | Importin subunit alpha-6/7 | 7 | 0.78 | 0.0287 |
| Unigene004822.p1 | Ras-related protein Rab-27A | 6 | 0.63 | 0.0277 |
| Unigene004984.p1 | Lysosomal-associated membrane protein 1/2 | 10 | 0.60 | 0.0080 |
| Unigene005731.p1 | Glyceraldehyde 3-phosphate dehydrogenase | 77 | 0.51 | 0.0329 |
| Unigene005921.p1 | Autophagy-related protein 5 | 7 | 0.54 | 0.0144 |
| Unigene006505.p1 | Cathepsin L | 23 | 0.65 | 0.0149 |
| Unigene002835.p1 | Actin related protein 2/3 complex, subunit 5 | 31 | 0.64 | 0.0135 |
| Unigene005574.p1 | Ferritin heavy chain | 59 | 0.53 | 0.0196 |
| Unigene000294.p1 | Titin | 13 | 0.77 | 0.0067 |
| Unigene006063.p1 | Niemann-Pick C2 protein | 34 | 0.53 | 0.0168 |
| Unigene003415.p1 | Beta-glucuronidase | 24 | 0.39 | 0.0020 |
| Unigene001334.p1 | Mannose-6-phosphate isomerase | 17 | 0.78 | 0.0336 |
| Unigene006110.p1 | Mannose receptor, C type | 5 | 0.63 | 0.0190 |
| Unigene006396.p1 | Cystatin-A/B | 23 | 0.52 | 0.0050 |
| Unigene000181.p1 | Vacuole morphology and inheritance protein 14 | 5 | 0.60 | 0.0084 |
| Unigene000974.p1 | Abhydrolase domain-containing protein 5 | 9 | 0.66 | 0.0317 |
| Unigene000708.p1 | AP-1 complex subunit beta-1 | 15 | 1.64 | 0.0001 |
| GO-ID | GO-Term | Category | p-Value | Diff |
|---|---|---|---|---|
| GO:0043202 | Lysosomal lumen | C | 0.00 | 10 |
| GO:0008305 | Integrin complex | C | 0.00 | 10 |
| GO:0005764 | Lysosome | C | 0.01 | 19 |
| GO:0045298 | Tubulin complex | C | 0.01 | 7 |
| GO:0000421 | Autophagosome membrane | C | 0.14 | 3 |
| GO:0001772 | Immunological synapse | C | 1.00 | 1 |
| GO:0019882 | Antigen processing and presentation | P | 0.00 | 5 |
| GO:0035099 | Hemocyte migration | P | 0.00 | 6 |
| GO:0000045 | Autophagosome assembly | P | 0.01 | 6 |
| GO:0032482 | Rab protein signal transduction | P | 0.01 | 7 |
| GO:0002252 | Immune effector process | P | 0.08 | 2 |
| GO:0030683 | Evasion or tolerance by virus of host immune response | P | 0.08 | 2 |
| GO:0002322 | B cell proliferation involved in immune response | P | 0.28 | 1 |
| GO:0002309 | T cell proliferation involved in immune response | P | 0.28 | 1 |
| GO:0002265 | Astrocyte activation involved in immune response | P | 0.28 | 1 |
| GO:0002842 | Positive regulation of T cell mediated immune response to tumor cell | P | 0.28 | 1 |
| GO:0002739 | Regulation of cytokine secretion involved in immune response | P | 0.28 | 1 |
| GO:0061844 | Antimicrobial humoral immune response mediated by antimicrobial peptide | P | 0.28 | 1 |
| GO:0002684 | Positive regulation of immune system process | P | 0.31 | 2 |
| GO:0045087 | Innate immune response | P | 0.37 | 5 |
| GO:0002682 | Regulation of immune system process | P | 0.41 | 3 |
| GO:0002840 | Regulation of T cell mediated immune response to tumor cell | P | 0.48 | 1 |
| GO:0002429 | Immune response-activating cell surface receptor signaling pathway | P | 0.48 | 1 |
| GO:0002286 | T cell activation involved in immune response | P | 0.48 | 1 |
| GO:0006955 | Immune response | P | 0.62 | 2 |
| GO:0002376 | Immune system process | P | 0.75 | 4 |
| GO:0002250 | Adaptive immune response | P | 1.00 | 1 |
| Map ID | Map Name | p-Value | Diff |
|---|---|---|---|
| ko04142 | Lysosome | 0.00 | 23 |
| ko04141 | Protein processing in endoplasmic reticulum | 0.00 | 26 |
| ko04512 | ECM-receptor interaction | 0.01 | 8 |
| ko00511 | Other glycan degradation | 0.01 | 7 |
| ko05205 | Proteoglycans in cancer | 0.01 | 16 |
| ko00531 | Glycosaminoglycan degradation | 0.01 | 5 |
| ko04510 | Focal adhesion | 0.01 | 16 |
| ko04612 | Antigen processing and presentation | 0.01 | 7 |
| ko04140 | Autophagy animal | 0.02 | 13 |
| ko04144 | Endocytosis | 0.04 | 16 |
| ko03420 | Nucleotide excision repair | 0.04 | 5 |
| ko05134 | Legionellosis | 0.05 | 7 |
| ko03010 | Ribosome | 0.05 | 17 |
| ID | Source | Domain | p-Value | Diff |
|---|---|---|---|---|
| IPR005225 | InterPro | Small GTP-binding protein domain | 0.00 | 17 |
| IPR017853 | InterPro | Glycoside hydrolase superfamily | 0.00 | 14 |
| IPR013780 | InterPro | Glycosyl hydrolase, all-beta | 0.00 | 8 |
| IPR012675 | InterPro | Beta-grasp domain superfamily | 0.00 | 5 |
| PR00449 | PRINTS | Transforming protein P21 Ras signature | 0.00 | 11 |
| IPR002939 | InterPro | Chaperone DnaJ, C-terminal | 0.01 | 4 |
| IPR008971 | InterPro | HSP40/DnaJ peptide-binding | 0.01 | 4 |
| cd10747 | CDD | DnaJ_C | 0.01 | 4 |
| IPR012676 | InterPro | TGS-like | 0.01 | 4 |
| PS51419 | ProSiteProfiles | Small GTPase Rab1 family profile. | 0.01 | 7 |
| IPR020568 | InterPro | Ribosomal protein S5 domain 2-type fold | 0.01 | 8 |
| IPR013320 | InterPro | Concanavalin A-like lectin/glucanase domain superfamily | 0.01 | 8 |
| IPR032695 | InterPro | Integrin domain superfamily | 0.02 | 5 |
| IPR029044 | InterPro | Nucleotide-diphospho-sugar transferases | 0.02 | 5 |
| IPR001806 | InterPro | Small GTPase superfamily | 0.02 | 10 |
| IPR018526 | InterPro | Glycoside hydrolase, family 29, conserved site | 0.02 | 3 |
| IPR012347 | InterPro | Ferritin-like | 0.02 | 3 |
| cd14752 | CDD | GH31_N | 0.02 | 3 |
| IPR031919 | InterPro | Alpha-L-fucosidase, C-terminal | 0.02 | 3 |
| IPR009078 | InterPro | Ferritin-like superfamily | 0.02 | 3 |
| IPR036156 | InterPro | Beta-Galactosidase/glucuronidase domain superfamily | 0.02 | 3 |
| IPR000933 | InterPro | Glycoside hydrolase, family 29 | 0.02 | 3 |
| IPR016286 | InterPro | Alpha-L-fucosidase, metazoa-type | 0.02 | 3 |
| IPR004095 | InterPro | TGS | 0.02 | 3 |
| IPR001519 | InterPro | Ferritin | 0.02 | 3 |
| IPR000980 | InterPro | SH2 domain | 0.02 | 4 |
| IPR036860 | InterPro | SH2 domain superfamily | 0.02 | 4 |
| IPR015940 | InterPro | Ubiquitin-associated domain | 0.02 | 4 |
| IPR029048 | InterPro | Heat shock protein 70 kD, C-terminal domain superfamily | 0.02 | 4 |
| IPR006689 | InterPro | Small GTPase superfamily, ARF/SAR type | 0.02 | 4 |
| IPR001841 | InterPro | Zinc finger, RING-type | 0.04 | 1 |
| IPR009060 | InterPro | UBA-like superfamily | 0.04 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Ma, F.; Dong, J.; Li, L.; Ren, P.; Zhang, Y.; Wu, Y.; Wang, Y.; Liu, K.; Zhang, F. The Innate Immune Response to Infection by Polyascus gregaria in the Male Chinese Mitten Crab (Eriocheir sinensis), Revealed by Proteomic Analysis. Fishes 2021, 6, 57. https://doi.org/10.3390/fishes6040057
Yang Y, Ma F, Dong J, Li L, Ren P, Zhang Y, Wu Y, Wang Y, Liu K, Zhang F. The Innate Immune Response to Infection by Polyascus gregaria in the Male Chinese Mitten Crab (Eriocheir sinensis), Revealed by Proteomic Analysis. Fishes. 2021; 6(4):57. https://doi.org/10.3390/fishes6040057
Chicago/Turabian StyleYang, Yanping, Fengjiao Ma, Juanjuan Dong, Lianxing Li, Ping Ren, Yuning Zhang, Yatao Wu, Yinping Wang, Kai Liu, and Fang Zhang. 2021. "The Innate Immune Response to Infection by Polyascus gregaria in the Male Chinese Mitten Crab (Eriocheir sinensis), Revealed by Proteomic Analysis" Fishes 6, no. 4: 57. https://doi.org/10.3390/fishes6040057
APA StyleYang, Y., Ma, F., Dong, J., Li, L., Ren, P., Zhang, Y., Wu, Y., Wang, Y., Liu, K., & Zhang, F. (2021). The Innate Immune Response to Infection by Polyascus gregaria in the Male Chinese Mitten Crab (Eriocheir sinensis), Revealed by Proteomic Analysis. Fishes, 6(4), 57. https://doi.org/10.3390/fishes6040057