
Diets and Stable Isotope Signatures of Native and Nonnative Leucisid Fishes Advances Our Understanding of the Yellowstone Lake Food Web


Abstract
1. Introduction
2. Materials and Methods
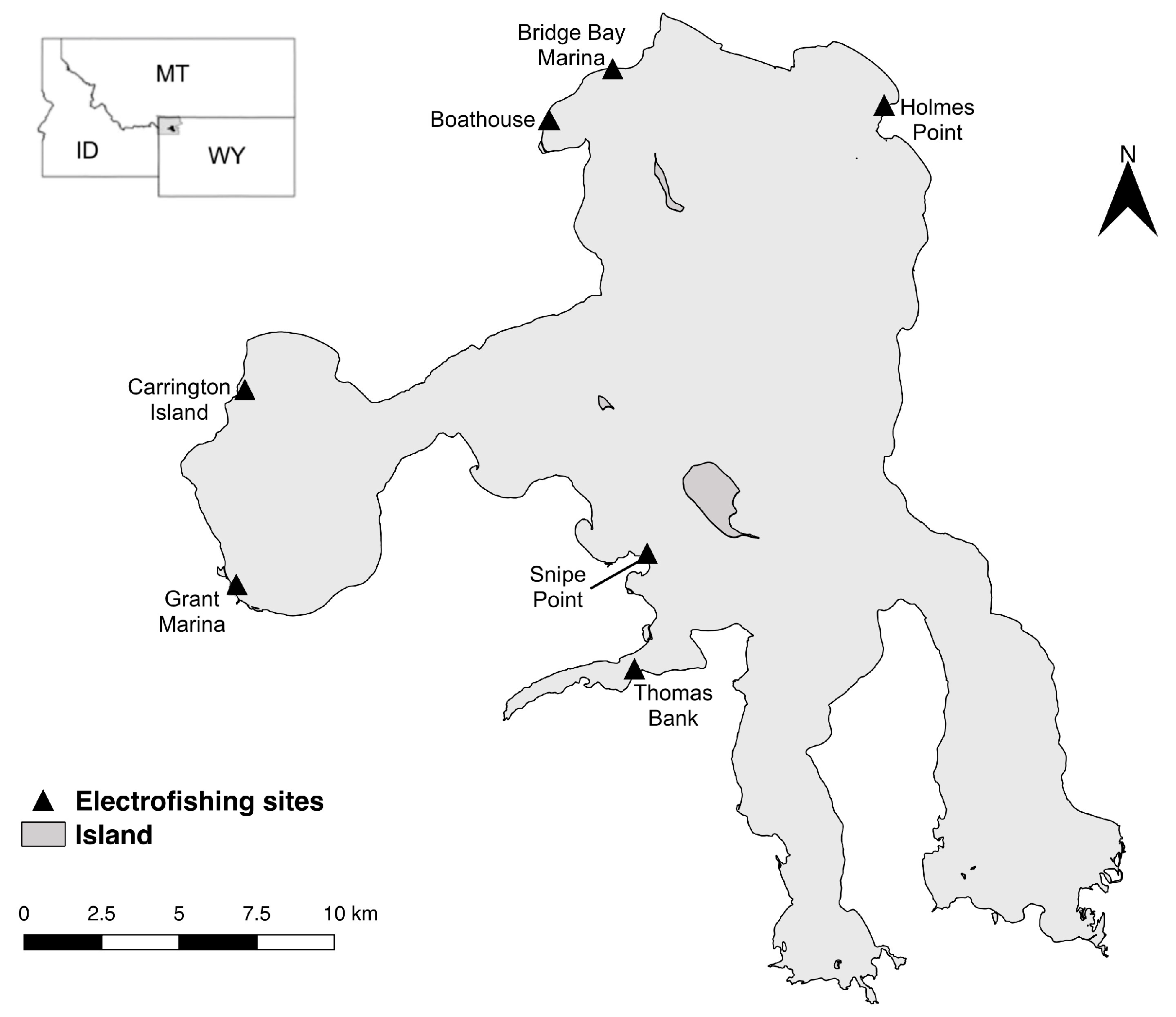
2.1. Study Area
2.2. Field Sampling
2.3. Laboratory Processing
2.4. Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chipps, S.; Garvey, J.E. Assessment of Diets and Feeding Patterns. In Analysis and Interpretation of Freshwater Fisheries Data; Guy, C.S., Brown, M.L., Eds.; American Fisheries Society: Bethesda, MA, USA, 2007; pp. 473–514. [Google Scholar]
- Kelly, E.N.; Schindler, D.W.; St Louis, V.L.; Donald, D.B.; Vladicka, K.E. Forest Fire Increases Mercury Accumulation by Fishes via Food Web Restructuring and Increased Mercury Inputs. Proc. Natl. Acad. Sci. USA 2006, 103, 19380–19385. [Google Scholar] [CrossRef]
- Albrecht, M.P.; Caramaschi, É.P.; Horn, M.H. Population Responses of Two Omnivorous Fish Species to Impoundment of a Brazilian Tropical River. Hydrobiologia 2009, 627, 181–193. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Xu, J.; Gu, B. Initial Response of Fish Trophic Niche to Hydrological Alteration in the Upstream of Three Gorges Dam. Ecol. Process. 2016, 5, 1–9. [Google Scholar] [CrossRef]
- Jo, H.; Jeppesen, E.; Ventura, M.; Buchaca, T.; Gim, J.S.; Yoon, J.D.; Kim, D.H.; Joo, G.J. Responses of Fish Assemblage Structure to Large-Scale Weir Construction in Riverine Ecosystems. Sci. Total Environ. 2019, 657, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Syslo, J.M.; Guy, C.S.; Koel, T.M. Feeding Ecology of Native and Nonnative Salmonids during the Expansion of a Nonnative Apex Predator in Yellowstone Lake, Yellowstone National Park. Trans. Am. Fish. Soc. 2016, 145, 476–492. [Google Scholar] [CrossRef]
- Ruzycki, J.R.; Beauchamp, D.A.; Yule, D.L. Effects of Introduced Lake Trout on Native Cutthroat Trout in Yellowstone Lake. Ecol. Appl. 2003, 13, 23–37. [Google Scholar] [CrossRef]
- Guy, C.S.; McMahon, T.E.; Fredenberg, W.A.; Smith, C.J.; Garfield, D.W.; Cox, B.S. Diet Overlap of Top-Level Predators in Recent Sympatry: Bull Trout and Nonnative Lake Trout. J. Fish Wildl. Manag. 2011, 2, 183–189. [Google Scholar] [CrossRef][Green Version]
- Zeug, S.C.; Brodsky, A.; Kogut, N.; Stewart, A.R.; Merz, J.E. Ancient Fish and Recent Invaders: White Sturgeon Acipenser Transmontanus Diet Response to Invasive-Species-Mediated Changes in a Benthic Prey Assemblage. Mar. Ecol. Prog. Ser. 2014, 514, 163–174. [Google Scholar] [CrossRef]
- Noble, R.L. Management of Forage Fishes in Impoundments of the Southern United States. Trans. Am. Fish. Soc. 1981, 110, 738–750. [Google Scholar] [CrossRef]
- Devries, D.R.; Stein, R.A. Manipulating Shad to Enhance Sport Fisheries in North America: An Assessment. N. Am. J. Fish. Manag. 1990, 10, 209–223. [Google Scholar] [CrossRef]
- Prabu, E.; Felix, S.; Felix, N.; Ahilan, B.; Ruby, P. An Overview on Significance of Fish Nutrition in Aquaculture Industry NADP-Development of Cost Efficient Feed for Enhancing Freshwater Fish Production View Project An Overview on Significance of Fish Nutrition in Aquaculture Industry. Int. J. Fish. Aquat. Stud. 2017, 5, 349–355. [Google Scholar]
- Cho, C.Y. Fish Nutrition, Feeds, and Feeding: With Special Emphasis on Salmonid Aquaculture. Food Rev. Int. 1990, 6, 333–357. [Google Scholar] [CrossRef]
- Garvey, J.E.; Dingledine, N.A.; Donovan, N.S.; Stein, R.A. Exploring Spatial and Temporal Variation within Reservoir Food Webs: Predictions for Fish Assemblages. Ecol. Appl. 1998, 8, 104–120. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Shuter, B.J.; Lester, N.P.; Rasmussen, J.B. Within- and among-Population Variation in the Trophic Position of a Pelagic Predator, Lake Trout (Salvelinus Namaycush). Can. J. Fish. Aquat. Sci. 2000, 57, 725–731. [Google Scholar] [CrossRef]
- Tronstad, L.M.; Hall, R.O.; Koel, T.M.; Gerow, K.G.; Hall, R.O., Jr.; Koel, T.M.; Gerow, K.G. Introduced Lake Trout Produced a Four-Level Trophic Cascade in Yellowstone Lake. Trans. Am. Fish. Soc. 2010, 139, 1536–1550. [Google Scholar] [CrossRef]
- Hobson, K.A. Tracing Origins and Migration of Wildlife Using Stable Isotopes: A Review. Oecologia 1999, 120, 314–326. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Primary Consumer Δ13C and Δ15N and the Trophic Position of Aquatic Consumers. Ecology 1999, 80, 1395–1404. [Google Scholar] [CrossRef]
- Bowen, S.H. Quantitative Description of the Diet. In Fisheries Techniques, 2nd ed.; Murphy, B.R., Willis, D., Eds.; American Fisheries Society: Bethesda, MA, USA, 1996; pp. 513–532. [Google Scholar]
- Jones, R.D.; Carty, D.G.; Gresswell, R.E.; Hudson, C.J.; Mahony, D.L. Fishery and Aquatic Management Program in Yellowstone National Park; U.S. Fish and Wildlife Service: Yellowstone National Park, WY, USA, 1993.
- Scott, J.C. A Study of the Cladocera and Copepoda of Yellowstone Lake, and Adjacent Lagoons, Yellowstone National Park, Wyoming. Ph.D. Thesis, University of Nebraska Omaha, Omaha, NE, USA, 1977. [Google Scholar]
- Dean, J.L. Fishery Management Program in Yellowstone National Park; Annual Project Report for 1970; U.S. Fish and Wildlife Service: Yellowstone National Park, WY, USA, 1971. [Google Scholar]
- Brown, C.J.D.; Graham, R.J. Observations on the Longnose Sucker in Yellowstone Lake. Trans. Am. Fish. Soc. 1954, 83, 38–46. [Google Scholar] [CrossRef]
- Furey, K.M.; Glassic, H.C.; Guy, C.S.; Koel, T.M.; Arnold, J.L.; Doepke, P.D.; Bigelow, P.E. Diets of Longnose Sucker in Yellowstone Lake, Yellowstone National Park, USA. J. Freshw. Ecol. 2020, 35, 291–303. [Google Scholar] [CrossRef]
- Syslo, J.M.; Brenden, T.O.; Guy, C.S.; Koel, T.M.; Bigelow, P.E.; Doepke, P.D.; Arnold, J.L.; Ertel, B.D. Could Ecological Release Buffer Suppression Efforts for Non-Native Lake Trout (Salvelinus Namaycush) in Yellowstone Lake, Yellowstone National Park? Can. J. Fish. Aquat. Sci. 2020, 77, 1010–1025. [Google Scholar] [CrossRef]
- Koel, T.M.; Tronstad, L.M.; Arnold, J.L.; Gunther, K.A.; Smith, D.W.; Syslo, J.M.; White, P.J. Predatory Fish Invasion Induces within and across Ecosystem Effects in Yellowstone National Park. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef]
- Hostetler, S.W.; Giorgi, F. Effects of a 2 × CO2 Climate on Two Large Lake Systems: Pyramid Lake, Nevada, and Yellowstone Lake, Wyoming. Glob. Planet. Change 1995, 10, 43–54. [Google Scholar] [CrossRef]
- Kaplinski, M.A. Geomorphology and Geology of Yellowstone Lake, Yellowstone National Park, Wyoming. Ph.D. Thesis, Northern Arizona University, Flagstaff, AZ, USA, 1991. [Google Scholar]
- Morgan, L.A.; Shanks, W.C., III; Lovalvo, D.A.; Johnson, S.Y.; Stephenson, W.J.; Pierce, K.L.; Harlan, S.S.; Finn, C.A.; Lee, G.; Webring, M. Exploration and Discovery in Yellowstone Lake: Results from High-Resolution Sonar Imaging, Seismic Reflection Profiling, and Submersible Studies. J. Volcanol. Geotherm. Res. 2003, 122, 221–242. [Google Scholar] [CrossRef]
- Varley, J.D.; Schullery, P. Yellowstone Fishes: Ecology, History, and Angling in the Park; Stackpole Books: Mechanicsburg, PA, USA, 1998. [Google Scholar]
- Koel, T.; Arnold, J.; Bigelow, P.; Doepke, P.; Ertel, B.; Ruhl, M. Yellowstone Fisheries and Aquatic Sciences: Annual Report, 2006; National Park Service: Yellowstone National Park, WY, USA, 2007.
- Benson, N.G. Limnology of Yellowstone Lake in Relation to the Cutthroat Trout; US Fish and Wildlife Service: Washington, DC, USA, 1961.
- Benke, A.C.; Huryn, A.D.; Smock, L.A.; Wallace, J.B. Length-Mass Relationships for Freshwater Macroinvertebrates in North America with Particular Reference to the Southeastern United States. J. N. Am. Benthol. Soc. 1999, 18, 308–343. [Google Scholar] [CrossRef]
- Dumont, H.J.; Van De Velde, I.; Dumont, S. The Dry Weight Estimate of Biomass in a Selection of Cladocera, Copepoda and Rotifera from the Plankton, Periphyton and Benthos of Continental Waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Hershey, A.E.; Northington, R.M.; Finlay, J.C.; Peterson, B.J. Stable Isotopes in Stream Food Webs. In Methods in Stream Ecology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 3–20. [Google Scholar]
- Hyslop, E.J. Stomach Contents Analysis—A Review of Methods and Their Application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Cortés, E. A Critical Review of Methods of Studying Fish Feeding Based on Analysis of Stomach Contents: Application to Elasmobranch Fishes. Can. J. Fish. Aquat. Sci. 1997, 54, 726–738. [Google Scholar] [CrossRef]
- Schoener, T.W. The Anolis Lizards of Bimini: Resource Partitioning in a Complex Fauna. Ecology 1968, 49, 704–726. [Google Scholar] [CrossRef]
- Wallace, R.K. An Assessment of Diet-Overlap Indexes. Trans. Am. Fish. Soc. 1981, 110, 72–76. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing Isotopic Niche Widths among and within Communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Batschelet, E. Circular Statistics in Biology; Academic Press: London, UK, 1981. [Google Scholar]
- Wilmot, O.; Tronstad, L.; Hall, R.O.; Koel, T.; Arnold, J. Lake Trout–Induced Spatial Variation in the Benthic Invertebrates of Yellowstone Lake. Park Sci. 2016, 32, 25–35. [Google Scholar]
- Johannes, R.E.; Larkin, P.A. Competition for Food between Redside Shiners (Richardsonius Balteatus) and Rainbow Trout (Salmo Gairdneri) in Two British Columbia Lakes. J. Fish. Res. Board Can. 1961, 18, 203–220. [Google Scholar] [CrossRef]
- Biesinger, K.E. Studies of the Relationship of the Redside Shiner (Richardsonius Balteatus) and the Longnose Sucker (Catostomus Catostomus) to the Cutthroat Trout (Salmo Clarki) Population in Yellowstone Lake. In All Graduate Theses and Dissertations; Utah State University: Logan, UT, USA, 1961; p. 331. [Google Scholar]
- Welch, C.A. Seasonal and Age-Based Aspects of Diet of the Introduced Redside Shiner (Richardsonius Balteatus) in Ross Lake, Washington; Western Washington University: Bellingham, WA, USA, 2011. [Google Scholar]
- Simpson, J.C. Food Analysis of Some Important Species of Wyoming Forage Fishes; University of Wyoming: Laramie, WY, USA, 1941. [Google Scholar]
- McPhail, J.D.; Lindsey, C.C. Freshwater Fishes of Northwestern Canada and Alaska. Fish. Res. Board Can. Bull. 1970, 173, 1–381. [Google Scholar]
- Isaak, D.J.; Hubert, W.A.; Berry, C.R. Conservation Assessment for Lake Chub (Couesius Plumbeus), Mountain Sucker (Catostomus Platyrhynchus), and Finescale Dace (Phoxinus Neogaeus) in the Black Hills National Forest of South Dakota and Wyoming; USDA Forest Service: Washington, DC, USA, 2003.
- Pappantoniou, A.; Dale, G. Comparative Food Habits of Two Minnow Species: Blacknose Dace, Rhinichthys Atratulus and Longnose Dace, Rhinichthys Cataractae. J. Freshw. Ecol. 1982, 1, 361–364. [Google Scholar] [CrossRef]
- Mullen, D.M.; Burton, T.M. Size-Related Habitat Use by Longnose Dace (Rhinichthys Cataractae). Am. Midl. Nat. 1995, 133, 177–183. [Google Scholar] [CrossRef]
- Gerald, J.W. Food Habits of the Longnose Dace, Rhinichthys Cataractae. Copeia 1966, 1966, 478. [Google Scholar] [CrossRef]
- Gibbons, J.R.H.; Gee, J.H. Ecological Segregation between Longnose and Blacknose Dace (Genus Rhinichthys) in the Mink River, Manitoba. J. Fish. Res. Board Can. 1972, 29, 1245–1252. [Google Scholar] [CrossRef]
- Mullen, D.M. Size Related Habitat Use in a Riffle Dwelling Fish. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 1991. [Google Scholar]
- Clarke, L.R.; Vidergar, D.T.; Bennett, D.H. Stable Isotopes and Gut Content Show Diet Overlap among Native and Introduced Piscivores in a Large Oligotrophic Lake. Ecol. Freshw. Fish 2005, 14, 267–277. [Google Scholar] [CrossRef]
- Rubenson, E.S.; Lawrence, D.J.; Olden, J.D. Threats to Rearing Juvenile Chinook Salmon from Nonnative Smallmouth Bass Inferred from Stable Isotope and Fatty Acid Biomarkers. Trans. Am. Fish. Soc. 2020, 149, 350–363. [Google Scholar] [CrossRef]
- Power, M.; Power, G.; Reist, J.D.; Bajno, R. Ecological and Genetic Differentiation among the Arctic Charr of Lake Aigneau, Northern Québec. Ecol. Freshw. Fish 2009, 18, 445–460. [Google Scholar] [CrossRef]
- Michaud, W.K.; Perry, R.C.; Dempson, J.B.; Shears, M.; Power, M. Occurrence of Lake Chub, Couesius Plumbeus, in Northern Labrador. Can. Field-Nat. 2010, 124, 113–117. [Google Scholar] [CrossRef]
- Murdoch, A.; Klein, G.; Doidge, D.W.; Power, M. Assessing the Food Web Impacts of an Anadromous Arctic Charr Introduction to a Sub-Arctic Watershed Using Stable Isotopes. Fish. Manag. Ecol. 2013, 20, 302–314. [Google Scholar] [CrossRef]
- Schmidt, S.N.; Olden, J.D.; Solomon, C.T.; Vander Zanden, M.J. Quantitative Approaches to the Analysis of Stable Isotope Food Web Data. Ecology 2007, 88, 2793–2802. [Google Scholar] [CrossRef]
- Koel, T.M.; Arnold, J.L.; Bigelow, P.E.; Brenden, T.O.; Davis, J.D.; Detjens, C.R.; Doepke, P.D.; Ertel, B.D.; Glassic, H.C.; Gresswell, R.E.; et al. Yellowstone Lake Ecosystem Restoration: A Case Study for Invasive Fish Management. Fishes 2020, 5, 18. [Google Scholar] [CrossRef]
- Gauzens, B.; Barnes, A.; Giling, D.P.; Hines, J.; Jochum, M.; Lefcheck, J.S.; Rosenbaum, B.; Wang, S.; Brose, U. Fluxweb: An R Package to Easily Estimate Energy Fluxes in Food Webs. Methods Ecol. Evol. 2018, 10, 270–279. [Google Scholar] [CrossRef]
- Christensen, V.; Walters, C.J. No Title. Ecol. Modell. 2004, 172, 109–139. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
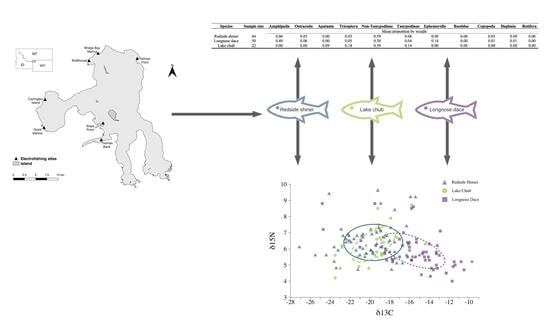
| Frequency of Occurrence | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Sample Size | Amphipoda | Ostracoda | Apatania | Tricoptera | Non-Tanypodinae | Tanypodinae | Ephemerella | Baetidae | Copepoda | Daphnia | Rotifera |
| Redside shiner | 44 | 0.07 | 0.02 | 0.00 | 0.02 | 0.66 | 0.09 | 0.14 | 0.02 | 0.11 | 0.36 | 0.02 |
| Longnose dace | 50 | 0.50 | 0.04 | 0.00 | 0.06 | 0.58 | 0.10 | 0.16 | 0.00 | 0.04 | 0.02 | 0.08 |
| Lake chub | 22 | 0.14 | 0.00 | 0.05 | 0.09 | 0.45 | 0.09 | 0.05 | 0.00 | 0.14 | 0.59 | 0.00 |
| Proportion by number | ||||||||||||
| Redside shiner | 44 | 0.01 | 0.01 | 0.00 | 0.00 | 0.75 | 0.02 | 0.01 | 0.00 | 0.04 | 0.16 | 0.00 |
| Longnose dace | 50 | 0.10 | 0.01 | 0.00 | 0.00 | 0.57 | 0.25 | 0.01 | 0.00 | 0.02 | 0.00 | 0.03 |
| Lake chub | 22 | 0.01 | 0.00 | 0.00 | 0.03 | 0.47 | 0.01 | 0.00 | 0.00 | 0.02 | 0.45 | 0.00 |
| Mean proportion by weight | ||||||||||||
| Redside shiner | 44 | 0.06 | 0.03 | 0.00 | 0.03 | 0.59 | 0.08 | 0.09 | 0.00 | 0.03 | 0.09 | 0.00 |
| Longnose dace | 50 | 0.49 | 0.06 | 0.00 | 0.05 | 0.20 | 0.04 | 0.14 | 0.00 | 0.01 | 0.01 | 0.00 |
| Lake chub | 22 | 0.00 | 0.00 | 0.09 | 0.18 | 0.59 | 0.14 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Species Comparison | D |
|---|---|
| Redside shiner–longnose dace | 0.47 |
| Redside shiner–lake chub | 0.69 |
| Lake chub–longnose dace | 0.29 |
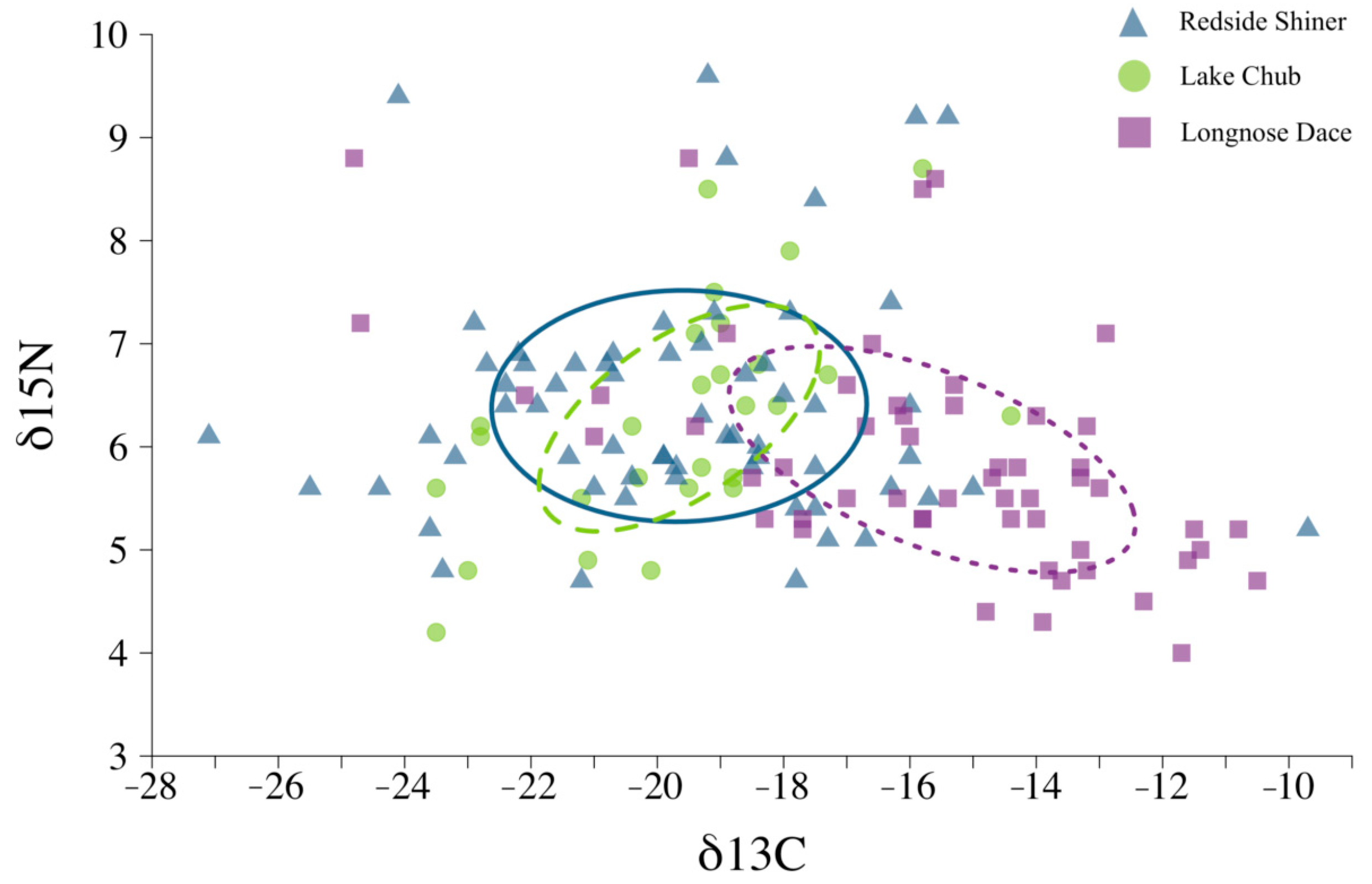
| Species | N | δ15N (‰) | δ13C (‰) |
|---|---|---|---|
| Redside shiner | 63 | 6.40 (6.13–6.67) | −19.65 (−20.38–−18.92) |
| Longnose dace | 27 | 5.88 (5.47–6.29) | −15.65 (−16.85–−14.45) |
| Lake chub | 54 | 6.28 (4.61–7.96) | −19.65 (−20.24–−19.06) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glassic, H.C.; Guy, C.S.; Koel, T.M. Diets and Stable Isotope Signatures of Native and Nonnative Leucisid Fishes Advances Our Understanding of the Yellowstone Lake Food Web. Fishes 2021, 6, 51. https://doi.org/10.3390/fishes6040051
Glassic HC, Guy CS, Koel TM. Diets and Stable Isotope Signatures of Native and Nonnative Leucisid Fishes Advances Our Understanding of the Yellowstone Lake Food Web. Fishes. 2021; 6(4):51. https://doi.org/10.3390/fishes6040051
Chicago/Turabian StyleGlassic, Hayley C., Christopher S. Guy, and Todd M. Koel. 2021. "Diets and Stable Isotope Signatures of Native and Nonnative Leucisid Fishes Advances Our Understanding of the Yellowstone Lake Food Web" Fishes 6, no. 4: 51. https://doi.org/10.3390/fishes6040051
APA StyleGlassic, H. C., Guy, C. S., & Koel, T. M. (2021). Diets and Stable Isotope Signatures of Native and Nonnative Leucisid Fishes Advances Our Understanding of the Yellowstone Lake Food Web. Fishes, 6(4), 51. https://doi.org/10.3390/fishes6040051