Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes


Abstract
1. Introduction
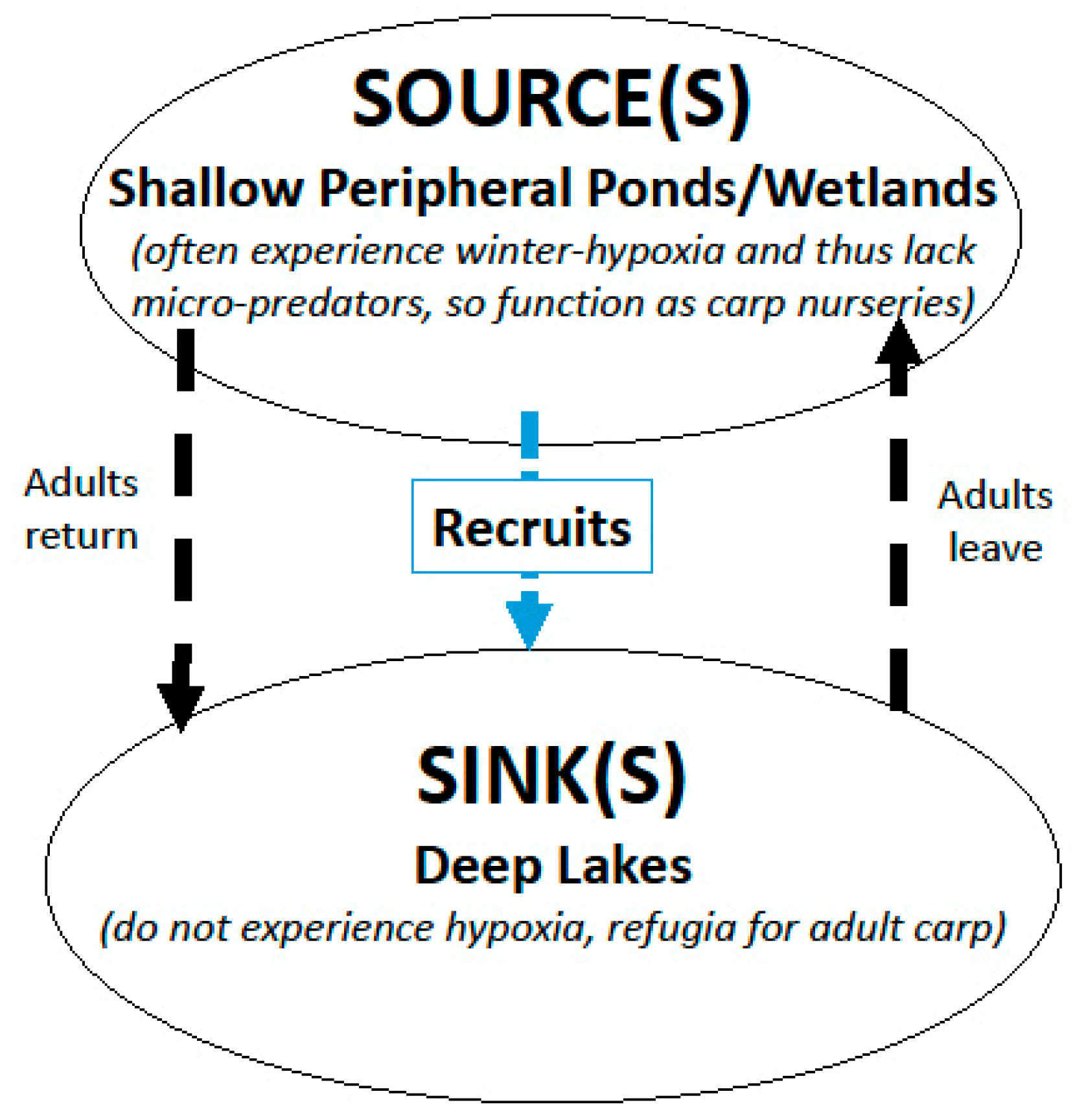
2. Overarching Strategy and Approach
- Step 1: Assessing the density and distribution of juvenile and adult carp. We found that before we could consider carp control, we first had to determine the location and number of adult and juvenile carp in the systems, the extent of their populations (carp movement patterns), and whether there were sources and sinks. We developed several techniques including different types of netting, electrofishing, genetic analysis, environmental DNA, and aging to conduct this process. These are described below in the two case studies and related publications.
- Step 2: Controlling and reducing the source(s) of juvenile carp while removing adult carp. While developing an understanding of how many carp there were, we realized that we could adopt these assessment techniques and approaches to affect control. Controlling sources required that we develop strategies different from those used for sinks. Many of these were adaptations of assessment strategies (ex. seining used for mark-recapture study were modified to include removal) and conducted as part of assessment itself. Different strategies had to be developed for different locales/carp populations. These are described in the case studies described below.
- Step 3. Monitoring and Adapting. Even after carp populations had been reduced and controlled, it was evident that continued systematic monitoring was necessary owing to the high fecundity of this species and how its reproductive success was influenced by variable winter conditions. If/as increases in carp numbers were noted, new techniques to address these increases had to be developed. This adaptive effort was strongly based on assessment techniques and is described below. These data are largely taken from agency records.
3. Case-Studies of Sustainable Carp Control
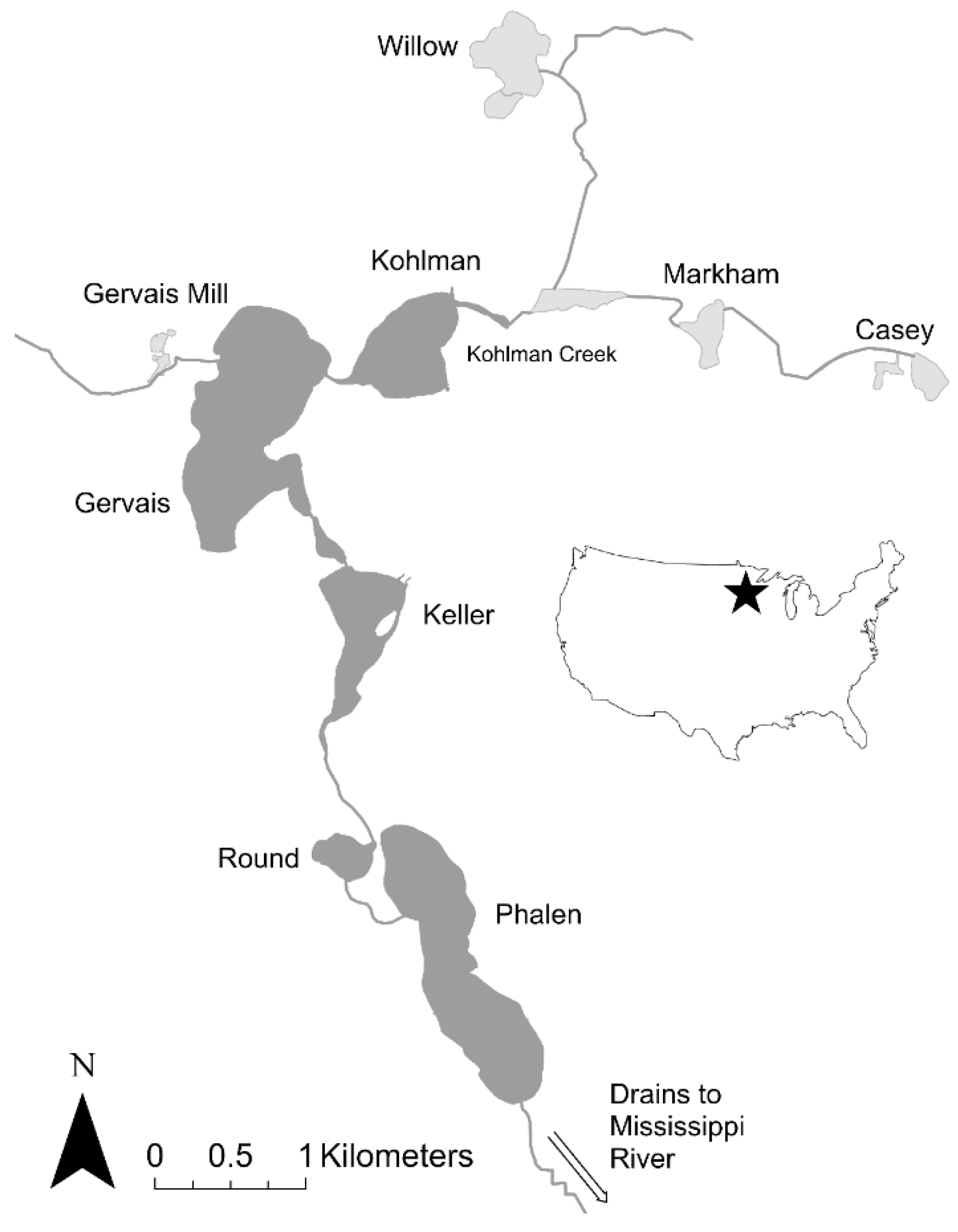
3.1. Case Study 1: The Phalen Chain of Lakes
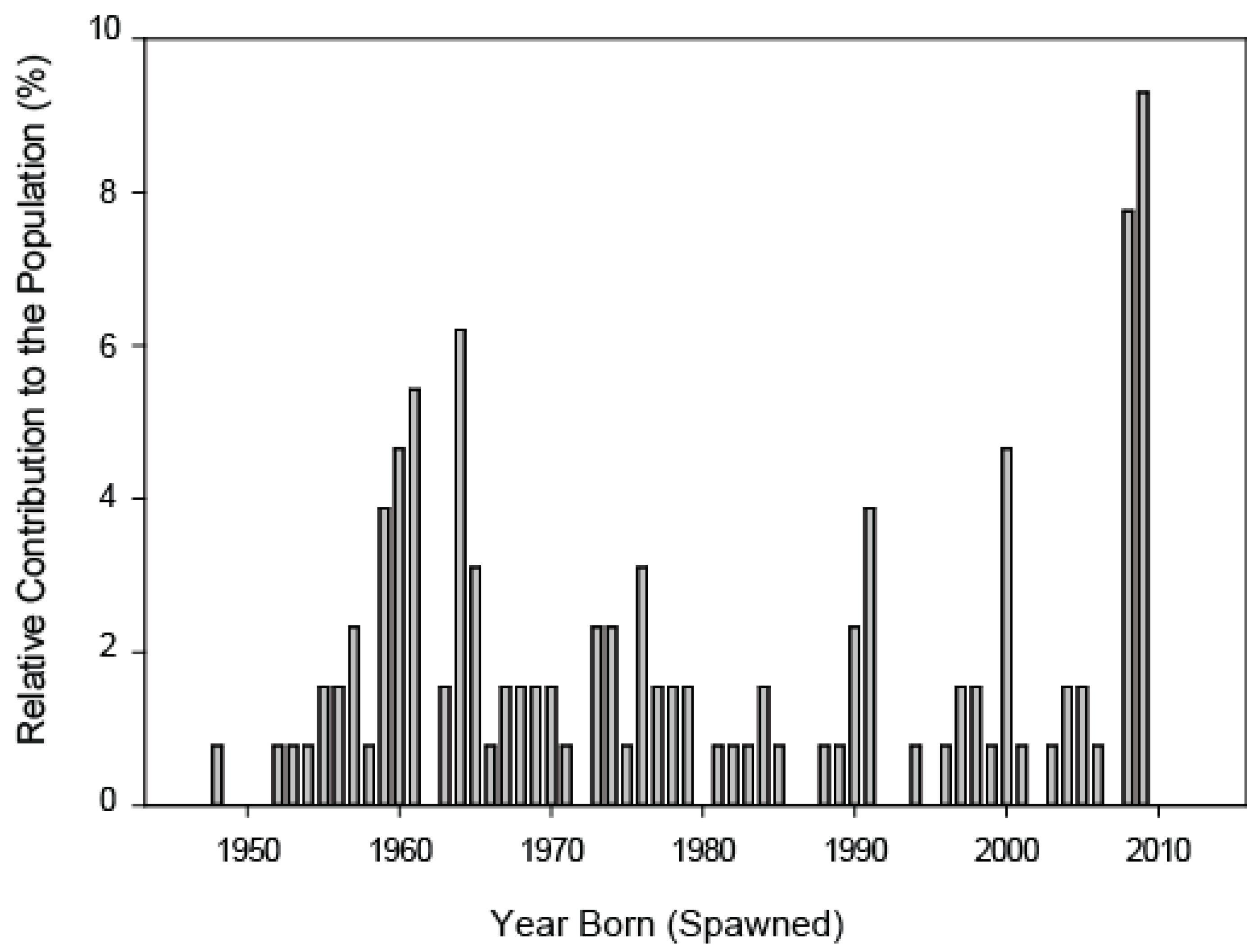
3.1.1. Step 1: Assessing the Density and Distribution of Adult and Juvenile Carp
3.1.2. Step 2: Controlling the Number of Adult Common Carp while Controlling Recruitment/Sources
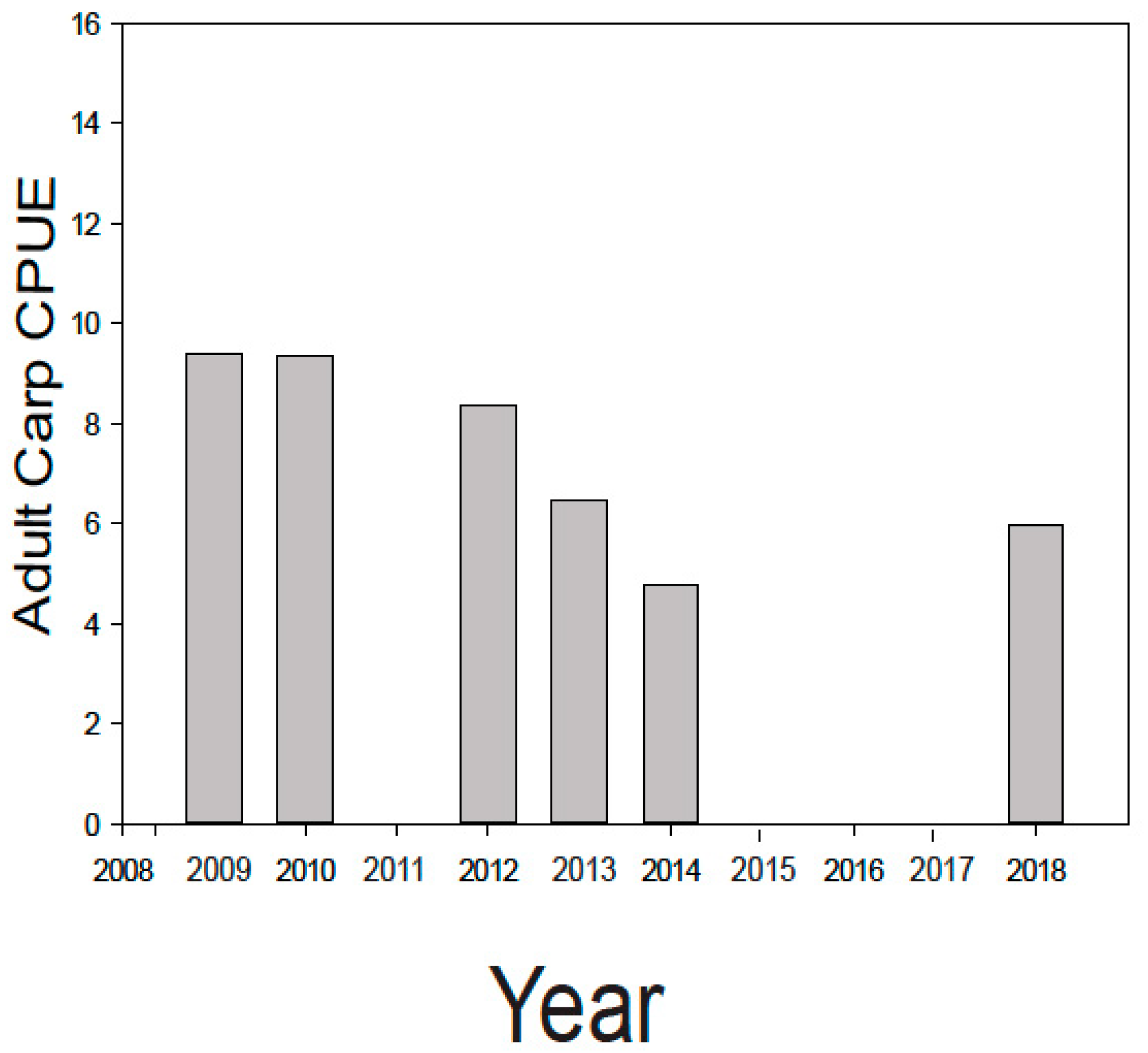
3.1.3. Step 3: Monitoring Carp Populations and Adapting New Management Approaches If/As Needed
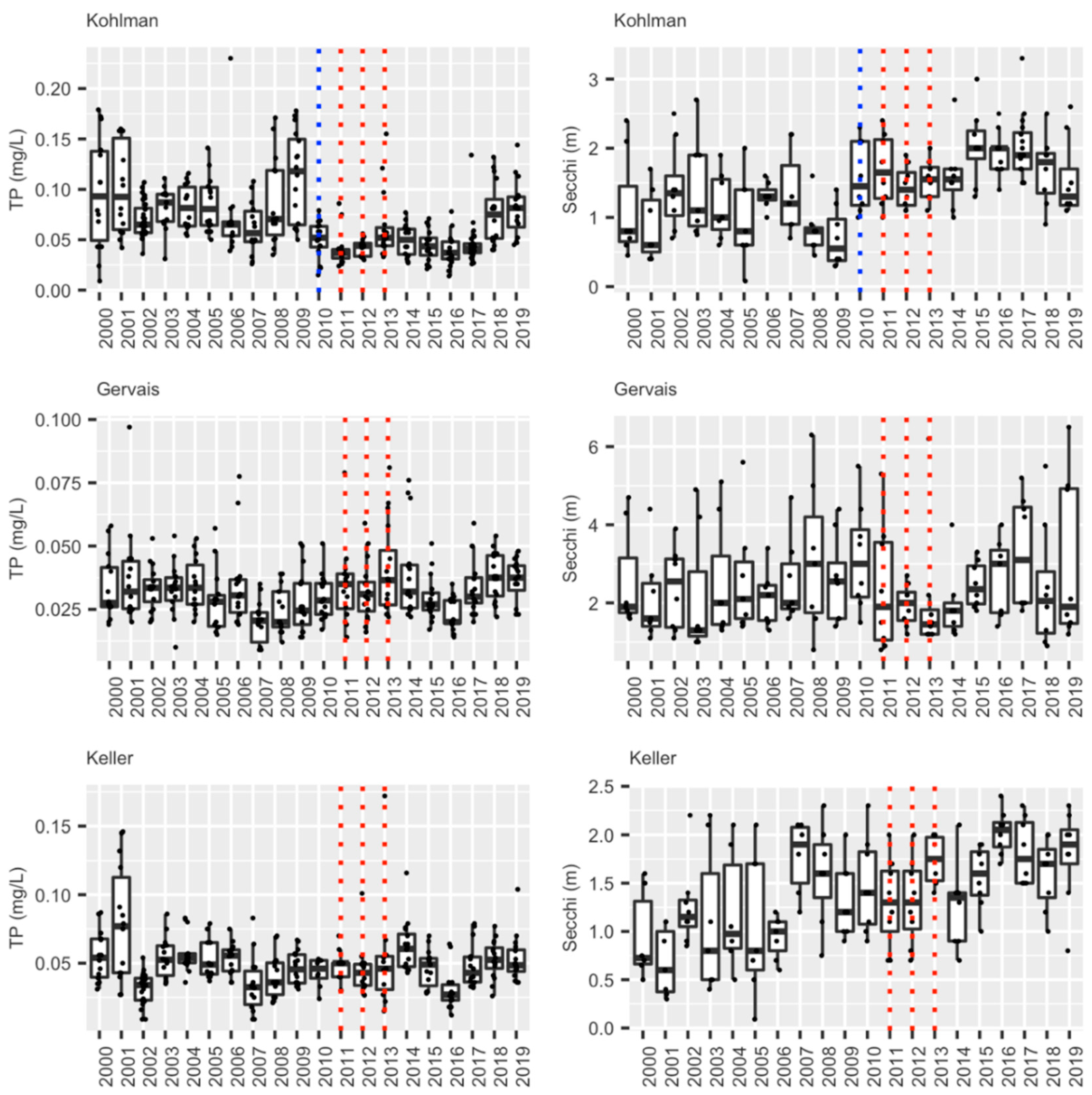
3.1.4. Summary: Carp, Water Quality, and Current Status
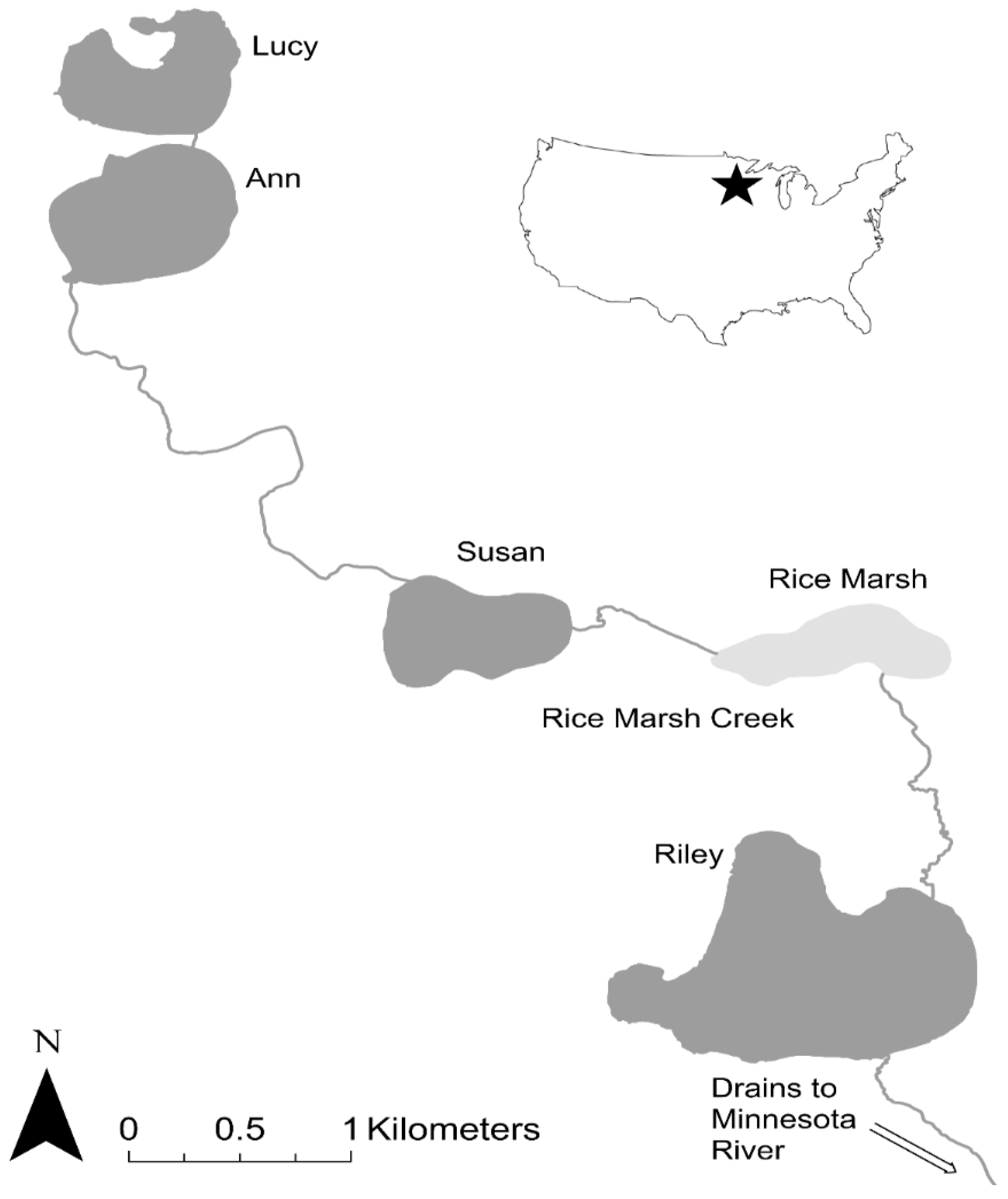
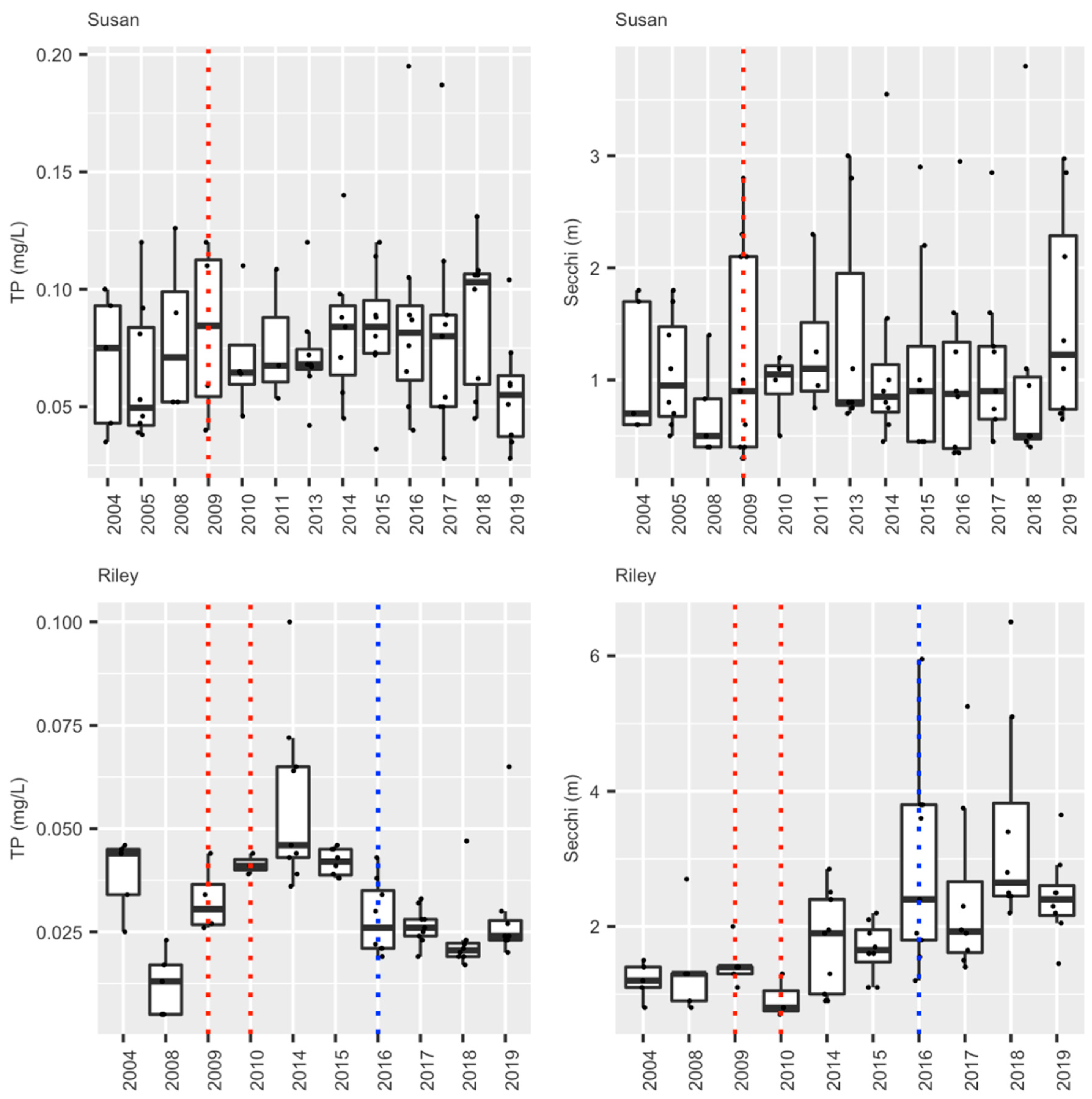
3.2. Study 2. The Riley Chain of Lakes
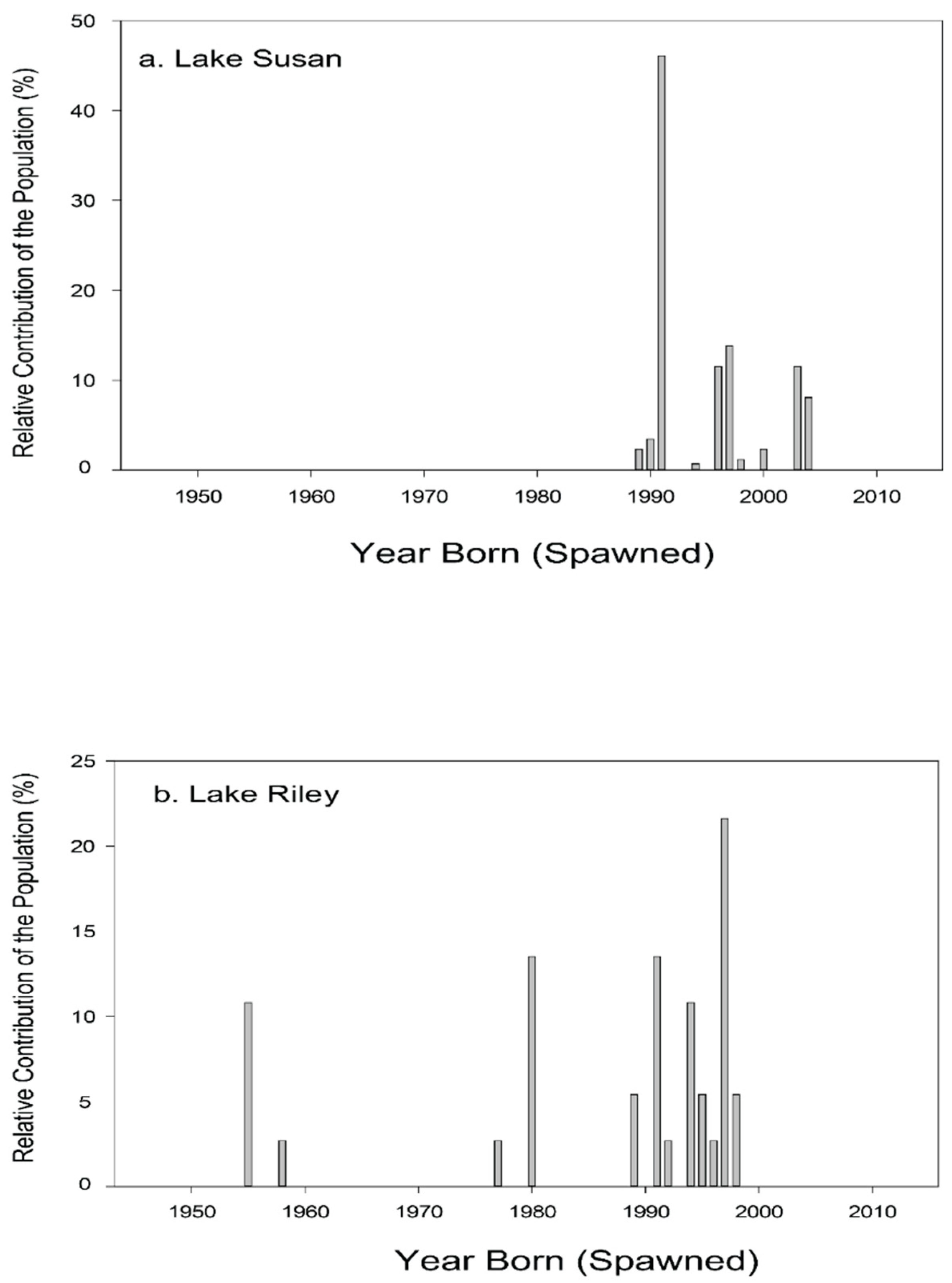
3.2.1. Step 1: Assessing the Density, Abundance, and Distribution of Adult and Juvenile Carp
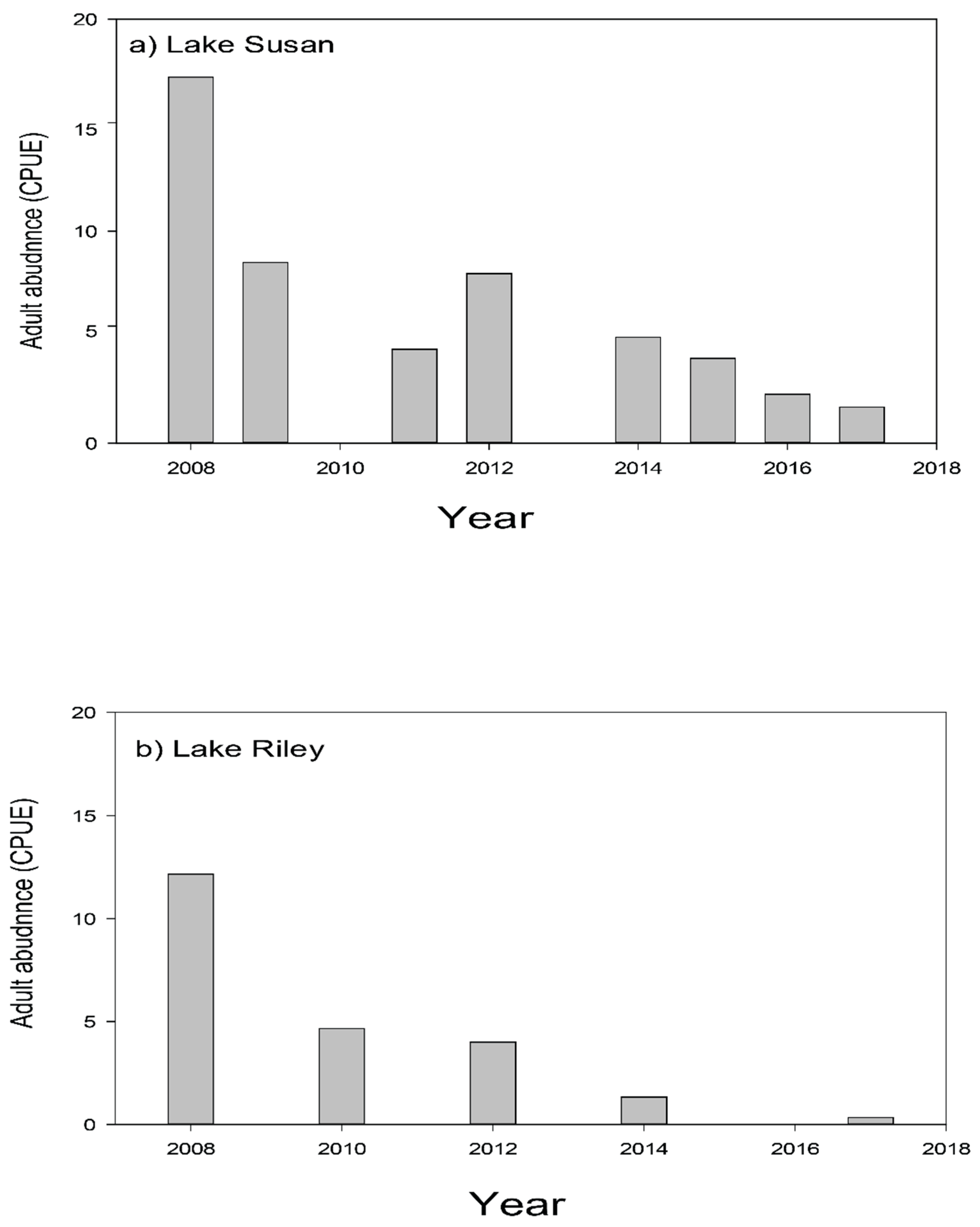
3.2.2. Step 2—Reducing and Controlling Adult Carp while Controlling Recruitment
3.2.3. Step 3—Monitoring Carp Populations and Using Adaptive Management Approaches
3.2.4. Summary: Carp, Water Quality, and Current Status
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-native fishes and climate change: Predicting species responses to warming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Balon, E.K. Origin and domestication of the wild carp, Cyprinus carpio: From Roman gourmets to the swimming flowers. Aquaculture 1995, 129, 3–48. [Google Scholar] [CrossRef]
- Sorensen, P.W.; Bajer, P.G. Carp, common. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmanek, M., Eds.; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Kohlmann, K.; Kersten, P. Deeper insight into the origin and spread of European common carp (Cyprinus carpio carpio) based on mitochondrial D-loop sequence polymorphisms. Aquaculture 2013, 376, 97–104. [Google Scholar] [CrossRef]
- Funge-Smith, S. Review of the State of the World Fisheries Resources: Inland Fisheries; FAO Fisheries and Aquaculture Circular: Rome, Italy, 2018; 253p. [Google Scholar]
- Bajer, P.G.; Sullivan, G.; Sorensen, P.W. Effects of a rapidly increasing population of common carp on vegetative cover and waterfowl in a recently restored Midwestern shallow lake. Hydrobiologia 2009, 632, 235–245. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Effects of Common Carp on Aquatic Ecosystems 80 Years after “Carp as a Dominant”: Ecological Insights for Fisheries Management. Rev. Fish. Sci. 2009, 17, 524–537. [Google Scholar] [CrossRef]
- Matsuzaki, S.S.; Usio, N.; Takamura, N.; Washitani, I. Contrasting impacts of invasive engineers on freshwater ecosystems: An experiment and meta-analysis. Oecologia 2008, 158, 673–686. [Google Scholar] [CrossRef]
- Verrill, C.R.; Berry, B.R., Jr. Effectiveness of a lake drawdown and electrical barrier for reducing common carp and bigmouth buffalo abudances. N. Am. J. Fish. Manag. 1995, 5, 137–141. [Google Scholar] [CrossRef]
- Meronek, T.G.; Bouchard, P.M.; Buckner, E.R.; Burri, T.M.; Demmerly, K.K.; Hatleli, D.C.; Klumb, R.A.; Schmidt, S.H.; Coble, D.W. A Review of Fish Control Projects. N. Am. J. Fish. Manag. 1996, 16, 63–74. [Google Scholar] [CrossRef]
- Roberts, J.; Tilzey, R. Controlling Carp: Options for Australia; CSIRO Press: Griffith, Australia, 1997; 141p. [Google Scholar]
- Weber, M.J.; Hennen, M.J.; Brown, M.L.; Lucchesi, D.O.; Sauver, T.R.S. Compensatory response of invasive common carp Cyprinus carpio to harvest. Fish. Res. 2016, 179, 168–178. [Google Scholar] [CrossRef]
- Balon, E.K. The common carp, Cyprinus carpio: Its wild origin, domestication in aquaculture, and selection as colored nishikigoi. Guelph Ichthyol. Rev. 1995, 3, 1–55. [Google Scholar]
- Bajer, P.G.; Cross, T.K.; Lechelt, J.D.; Chizinski, C.J.; Weber, M.J.; Sorensen, P.W. Across-ecoregion analysis suggests a hierarchy of ecological filters that regulate recruitment of a globally invasive fish. Divers. Distrib. 2015, 21, 500–510. [Google Scholar] [CrossRef]
- McCrimmon, H.R. Carp in Canada. In Bulletin Fisheries Research; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1968; 165p. [Google Scholar]
- Bajer, P.G.; Sorensen, P.W. Recruitment and abundance of an invasive fish, the common carp, is driven by its propensity to invade and reproduce in basins that experience winter-time hypoxia in interconnected lakes. Biol. Invasions 2009, 12, 1101–1112. [Google Scholar] [CrossRef]
- Billard, R. Carp: Biology and Culture; Praxis Publishing: Chichester, UK, 1999. [Google Scholar]
- Silbernagel, J.J.; Sorensen, P.W. Direct Field and Laboratory Evidence that a Combination of Egg and Larval Predation Controls Recruitment of Invasive Common Carp in Many Lakes of the Upper Mississippi River Basin. Trans. Am. Fish. Soc. 2013, 142, 1134–1140. [Google Scholar] [CrossRef]
- Koehn, J.D. Carp (Cyprinus carpio) as a powerful invader in Australian waterways. Freshw. Biol. 2004, 49, 882–894. [Google Scholar] [CrossRef]
- Lechelt, J.; Bajer, P. Elucidating the mechanism underlying the productivity-recruitment hypothesis in the invasive common carp. Aquat. Invasions 2016, 11, 469–482. [Google Scholar] [CrossRef]
- Dauphinais, J.D.; Miller, L.M.; Swanson, R.G.; Sorensen, P.W. Source-sink dynamics explain the distribution and persistence of an invasive population of common carp across a model Midwestern watershed. Biol. Invasions 2018, 20, 1961–1976. [Google Scholar] [CrossRef]
- Vilizzi, L.; Tarkan, A.S.; Copp, G.S. Experimental evidence from causal criteria analysis for the effects of common carp Cyprinus carpio on freshwater ecosystems: A global perspective. Rev. Fish. Sci. Aquac. 2015, 23, 253–290. [Google Scholar] [CrossRef]
- Bajer, P.G.; Beck, M.W.; Cross, T.K.; Koch, J.; Bartodziej, B.; Sorensen, P.W. Biological invasion by a benthivorous fish reduced the cover and species richness of aquatic plants in most lakes of a large North American ecoregion. Glob. Chang. Biol. 2016, 22, 3937–3947. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, H.R. Sources, Sinks, and Population Regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Travis, J.M.J.; Park, K. Spatial structure and the control of invasive alien species. Anim. Conserv. 2004, 7, 321–330. [Google Scholar] [CrossRef]
- Bajer, P.G.; Chizinski, C.J.; Silbernagel, J.J.; Sorensen, P.W. Variation in native micro-predator abundance explains recruitment of a mobile invasive fish, the common carp, in a naturally unstable environment. Biol. Invasions 2012, 14, 1919–1929. [Google Scholar] [CrossRef]
- Chizinski, C.J.; Bajer, P.G.; Headrick, M.E.; Sorensen, P.W. Different migratory strategies of invasive Common Carp and native Northern Pike in the American Midwest suggest an opportunity for selective management strategies. N. Am. J. Fish. Manag. 2016, 36, 769–779. [Google Scholar] [CrossRef]
- Bajer, P.G.; Sorensen, P.W. Using boat electrofishing to estimate the abundance of invasive Common Carp in small Midwestern lakes. N. Am. J. Fish. Manag. 2012, 32, 817–822. [Google Scholar] [CrossRef]
- Bajer, P.G.; Chizinski, C.J.; Sorensen, P.W. Using the Judas technique to locate and remove wintertime aggregations of invasive common carp. Fish. Manag. Ecol. 2011, 18, 497–505. [Google Scholar] [CrossRef]
- Osborne, J. Distribution, Abundance and Overwinter Survival of Young-of-the-Year Common Carp in a Midwestern Watershed. Master’s Thesis, University of Minnesota, Minneapolis, MN, USA, 2012, (unpublished). [Google Scholar]
- Brown, P.; Green, C.; Sivakumaran, K.P.; Giles, A.; Stoessel, D. Validating otolith annuli for use in age-determination of carp (Cyprinus carpio L.) from Victoria, Australia. Trans. Am. Fish. Soc. 2004, 133, 190–196. [Google Scholar] [CrossRef]
- Ghosal, R.; Eichmiller, J.J.; Witthuhn, B.A.; Sorensen, P.W. Attracting Common Carp to a bait site with food reveals strong positive relationships between fish density, feeding activity, environmental DNA, and sex pheromone release that could be used in invasive fish management. Ecol. Evol. 2018, 8, 6714–6727. [Google Scholar] [CrossRef]
- Bajer, P.G.; Lim, H.; Travaline, M.J.; Miller, B.D.; Sorensen, P.W. Cognitive aspects of food searching behavior in free-ranging wild Common Carp. Environ. Boil. Fishes 2010, 88, 295–300. [Google Scholar] [CrossRef]
- Eichmiller, J.J.; Bajer, P.G.; Sorensen, P.W. The Relationship between the Distribution of Common Carp and Their Environmental DNA in a Small Lake. PLoS ONE 2014, 9, e112611. [Google Scholar] [CrossRef]
- Huser, B.J.; Bajer, P.G.; Chizinski, C.J.; Sorensen, P.W. Effects of common carp (Cyprinus carpio) on sediment mixing depth and mobile phosphorus mass in the active sediment layer of a shallow lake. Hydrobiologia 2015, 763, 23–33. [Google Scholar] [CrossRef]
- Bajer, P.G.; Sorensen, P.W. Effects of common carp on phosphorus concentrations, water clarity, and vegetation density: A whole system experiment in a thermally stratified lake. Hydrobiologia 2014, 746, 303–311. [Google Scholar] [CrossRef]
- Bajer, P.G.; Parker, J.E.; Cross, T.K.; Venturelli, P.A.; Sorensen, P.W. Partial migration to seasonally-unstable habitat facilitates biological invasions in a predator-dominated system. Oikos 2015, 124, 1520–1526. [Google Scholar] [CrossRef]
- Knopik, J.M.; Newman, R.M. Transplanting aquatic macrophytes to restore the littoral community of a eutrophic lake after the removal of common carp. Lake Reserv. Manag. 2018, 34, 365–375. [Google Scholar] [CrossRef]
- Dunne, M.A.; Newman, R.M. Effect of light on macrophyte sprouting and assessment of viable seedbank to predict community composition. J. Aquat. Plant Manag. 2019, 57, 90–98. [Google Scholar]
- Banet, N. Partial Migration, Homing, Diel Activity, and Distribution of Adult Common Carp across a Large, Model Watershed in the North American Midwest. Master’s Thesis, University of Minnesota, Saint Paul, MN, USA, 2016, (unpublished). [Google Scholar]
- Colvin, M.E.; Pierce, C.L.; Stewart, T.W.; Grummer, S.E. Strategies to control a common carp population by pulsed commercial harvest. N. Am. J. Fish. Manag. 2012, 32, 1251–1264. [Google Scholar] [CrossRef]
- Poole, J.R.; Bajer, P.G. A small native predator reduces reproductive success of a large invasive fish as revealed by whole-lake experiments. PLoS ONE 2019, 14, e0214009. [Google Scholar] [CrossRef]
- Lechelt, J.; Kocian, M.; Bajer, P. Low downstream dispersal of young-of-year common carp from marshes into lakes in the Upper Mississippi Region and its implications for integrated pest management strategies. Manag. Biol. Invasions 2017, 8, 485–495. [Google Scholar] [CrossRef]
- Chapman, B.B.; Hulthén, K.; Brodersen, J.; Nilsson, P.A.; Skov, C.; Hansson, L.-A.; Brönmark, C. Partial migration in fishes: Causes and consequences. J. Fish Biol. 2012, 81, 456–478. [Google Scholar] [CrossRef]
- Sorensen, P.W.; Fine, J.M.; Dvornikovs, V.; Jeffrey, C.S.; Shao, F.; Wang, J.; Vrieze, L.A.; Anderson, K.R.; Hoye, T.R. Mixture of new sulfated steroids functions as a migratory pheromone in the sea lamprey. Nat. Chem. Biol. 2005, 1, 324–328. [Google Scholar] [CrossRef]
- Koel, T.M.; Arnold, J.L.; Bigelow, P.E.; Brenden, T.O.; Davis, J.D.; Detjens, C.R.; Doepke, P.D.; Ertel, B.D.; Glassic, H.C.; Gresswell, R.E.; et al. Yellowstone Lake ecosystem restoration: A case study for invasive fish management. Fishes 2020, 5, 18. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Size (ha) | Max Depth (m) | Adult Carp (CPUE) | Adult Carp (#) | Adult Carp Density | YOY Carp (CPUE) | Notes |
|---|---|---|---|---|---|---|---|
| Casey | 4.7 | 6.1 | 8.5 | 2585 | n.a. | 6.2 b | Winterkills |
| Markham | 7.0 | 2.0 | 8.2 | 211 | n.a. | 104 b | Winterkills |
| Willow | 21.1 | 2.5 | 0 | 0 | n.a. | 173 b | Winterkills |
| Kohlman | 29.9 | 2.7 | 11.2 a | 1668 a | 177 kg/ha c | 0 b | - |
| Gervais | 94.5 | 12.5 | 7.2 a | 3499 a | 177 kg/ha c | 0 b | - |
| G Mill Pond | 3.0 | 4.0 | n.s. | n.s. | n.s. | n.s. | - |
| Keller | 29.0 | 2.4 | 14.7 | 2103 | 177 kg/ha c | 0 b | - |
| Round | 14.3 | 2.5 | n.s. | n.s. | n.s. | n.s. | - |
| Phalen | 79.6 | 53 | 6.0 | 2504 | 106 kg/ha d | 0 b |
| Origin Lake | Summer 2009 | Fall 2009 | Winter 2009–2010 | Spring 2010 | Summer 2010 |
|---|---|---|---|---|---|
| Gervais Lake (Lake Where Detected) | |||||
| Kohlman | 1 (2%) | 0 (0%) | 19 (26%) | 6 (18%) | 4 (15%) |
| Gervais | 35 (71%) | 36 (61%) | 18 (25%) | 16 (47%) | 21 (78%) |
| Keller | 13 (27%) | 23 (39%) | 35 (49%) | 12 (35%) | 2 (7%) |
| Phalen | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Lake Keller | |||||
| Kohlman | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Gervais | 1 (9%) | 9 (11%) | 3 (33%) | 9 (17%) | 5 (8%) |
| Keller | 110 (91%) | 73 (89%) | 6 (67%) | 43 (83%) | 55 (92%) |
| Phalen | 0 (0) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Kohlman Lake | |||||
| Kohlman | 67 (70%) | 56 (84%) | 5 (83%) | 20 (77%) | 36 (68%) |
| Gervais | 21 (22%) | 11 (16%) | 1 (17%) | 5 (19%) | 7 (13%) |
| Keller | 8 (8%) | 0 (0%) | 0 (0%) | 1 (4%) | 10 (19%) |
| Phalen | 1 (1%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Lake Phalen | |||||
| Kohlman | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Gervais | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Keller | 1 (1%) | 26 (30%) | 4 (21%) | 3 (14%) | 12 (20%) |
| Phalen | 82 (99%) | 62 (70%) | 15 (79%) | 18 (86%) | 49 (80%) |
| Date | Removal Method | # Carp Caught | # Carp Removed |
|---|---|---|---|
| Winter 2010–11 | Gervais Winter Seine-1 a | 3537 b | 100 |
| Winter 2010–11 | Gervais Winter Seine-2 | 1509 | 1505 |
| Winter 2010–11 | Gervais Winter Seine-3 | 930 | 732 |
| Winter 2011–12 | Gervais Winter Seine | 73 | 71 |
| Fall 2012 | Kohlman Boxnet | 630 | 628 |
| Winter 2012–13 | Gervais Winter Seine | 827 | 825 |
| Spring 2011–2013 | Spawning Block/Trap | 381 | 351 |
| Summer 2013 | Keller Electrofishing | 8 | 8 |
| Summer 2013 | Gervais Electrofishing | 97 | 97 |
| Summer 2013 | Kohlman Electrofishing | 5 | 5 |
| Fall 2013 | Keller Boxnet | 446 | 446 |
| Fall 2013 | Gervais Boxnet | 192 | 192 |
| Summer 2013 | Drawdown Casey, Markham | - | 100+ |
| Summer 2014 | Drawdown Casey, Markham | - | 100+ |
| Winter 2014–2015 | Aerate Casey, Markham | - | - |
| Fall 2015 | Gervais Boxnet | 302 | 302 |
| Fall 2016 | Gervais Boxnet * | 324 | 324 * |
| Year | Lakes | ||||||
|---|---|---|---|---|---|---|---|
| - | Casey | Markham | Willow | Kohlman | Gervais | Keller | Phalen |
| 1999 | 0 D | 0 | - | 0 D | 0 D | 0 D | 0 D |
| 2000 | - | - | - | - | - | - | - |
| 2001 | 25 D | - | - | - | - | - | - |
| 2002 | - | - | - | - | - | - | - |
| 2003 | - | - | - | - | - | - | - |
| 2004 | - | - | - | 0.33 D | - | - | 0 D |
| 2005 | - | - | - | 1.3 D | 0.56 D | 0.5 D | - |
| 2006 | - | - | - | - | - | - | - |
| 2007 | 28 D | - | - | - | - | 0 D | - |
| 2008 | - | - | - | - | - | - | - |
| 2009 S | 6.2 * | 104 * | 173 * | 0 * | 0 * | 0 * | 0 * |
| 2010 | 3.8 | 47 | 5.4 W | 0 | 0 | - | - |
| 2011 | 2 | 0.8 | 0 | 0 | 0 | - | - |
| 2012 | 8 | 0.7 | 0 | 0 | 0 | - | - |
| 2013 | 0 | 68.8 DD | 0 | 0 | 0 | - | - |
| 2014 | -DD | -DD | - | - | - | - | - |
| 2015 | 0 | 0 | - | 1 | - | - | 0 |
| 2016 | 0 | 0 | - | 0 | - | - | - |
| 2017 | - | - | - | 0 | - | - | - |
| 2018 | 0 | 0 | - | 0 | - | 0 | 0 |
| Lake | Size (ha) | Max Depth(m) | Adult Carp (CPUE) | Adult Carp (#) | Adult Carp (kg/ha) | YOY Carp (CPUE) | Notes |
|---|---|---|---|---|---|---|---|
| Lucy a | 34.6 | 6.0 | 8.15 b | 808 c | 70 kg/ha d | n.s. | winterkills |
| Ann | 44.1 | 13.7 | 3.57 e | 662 c | 66 kg/ha d | 0 | - |
| Susan | 35.1 | 5.1 | 17.26 b | 4181 c | 307/kg/h e | 0.4 | aerated |
| RiceMarshL | 40.0 | 3.0 | 3.0 | 178 c | 16 kg/h | 0 | winterkills |
| Riley | 118.8 | 14.7 | 12.16 b | 6419 c | 176 kg/ha d | 0 | - |
| Original Lake | Spring 2006 | Summer 2006 | Winter 2006–7 | Summer 2007 | Winter 2007–8 |
|---|---|---|---|---|---|
| Lake Susan (Lake Where Detected) | |||||
| Susan | 16 (100%) | 10 (62%) | 15 (88%) | 24 (100%) | 26 (79%) |
| RiceM | - | 6 (38%) | 1 (6%) | 0 (0%) | 7 (21%) |
| Riley | - | 0 (0%) | 1 (6%) | 0 (0%) | 0 (0%) |
| Rice Marsh Lake | |||||
| Susan | - | 6 | 1 | 7 (0%) | 0 (0%) |
| RiceM | 15 (100%) | 15 (63%) | D | 0 (0%) | 11 (61%) |
| Riley | - | 3 | D | 7 (0%) | 7 (39%) |
| Lake Riley | |||||
| Susan | - | 0 (%) | 0 (0%) | 0 (0%) | 0 (0%) |
| RiceM | - | 3 (15%) | 0 (0%) | 0 (0%) | 1 (6%) |
| Riley | 19 (100%) | 16 (85%) | 18 (100%) | 24 (100%) | 17 (94%) |
| Year | Lakes (CPUE) | ||||
|---|---|---|---|---|---|
| - | Lucy | Ann | Susan | RiceMarsh | Riley |
| 1999 | - | - | - | - | 0 D |
| 2000 | 0 D | 0 D | - | - | - |
| 2001 | - | - | - | - | - |
| 2002 | - | - | - | - | - |
| 2003 | - | - | 0 D | - | - |
| 2004 | - | - | - | - | - |
| 2005 | - | - | - | - | 0 D |
| 2006 S | 0 D | 0 D | 0 | 0 | 0 |
| 2007 | - | - | 0 | T | 0 |
| 2008 | - | - | 0 | 0 | 0 |
| 2009 | 0 | 0 | 1.3 | 0 | 0 |
| 2010 | 0.8 | 0.6 | 1 | 1 | 1 |
| 2011 | 0.2 | 0 | 0.5 | 1 | 1 |
| 2012 | 0.2 | 0.2 | 0.8 | 1 | 0.8 |
| 2013 | 0.6 | 0.4 | 0.8 | 1 | 1 |
| 2014 | 0.6 | 0.4 | 0.6 | 0.8 | 0.8 |
| 2015 | - | - | 0 | 0 | 0 |
| 2016 | - | - | 0 W | 0.2 W | 0 |
| 2017 | - | 0 D | 0 D | - | - |
| Date | Removal Method | # Carp Caught | # Carp Removed |
|---|---|---|---|
| Winter 2008–2009 | Susan Winter Seine | 3981 | 3425 |
| Spring 2009–2011 | Spawning block/trap | 242 | 242 a |
| Winter 2008–2009 | Riley Winter Seine | 4040 | 2940 |
| Winter 2009–2010 | Riley Winter Seine | 376 | 376 |
| Winter 2009–2010 | Riley Winter Seine | 2303 | 2303 |
| Spring 2009–2011 | Spawning block/trap | 21 | 21 b |
| Fall 2011 | Rice Marsh Lake aerated | - | - |
| Winter 2010–2011 | Lucy Winterkills | 630 | 500 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorensen, P.W.; Bajer, P.G. Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes. Fishes 2020, 5, 36. https://doi.org/10.3390/fishes5040036
Sorensen PW, Bajer PG. Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes. Fishes. 2020; 5(4):36. https://doi.org/10.3390/fishes5040036
Chicago/Turabian StyleSorensen, Peter W., and Przemyslaw G. Bajer. 2020. "Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes" Fishes 5, no. 4: 36. https://doi.org/10.3390/fishes5040036
APA StyleSorensen, P. W., & Bajer, P. G. (2020). Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes. Fishes, 5(4), 36. https://doi.org/10.3390/fishes5040036