The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review



Abstract
1. Introduction
2. Characterisation
2.1. Proteins and Amino Acids
2.2. Lipids and Others (Astaxanthin and Bioactives)
3. Digestibility
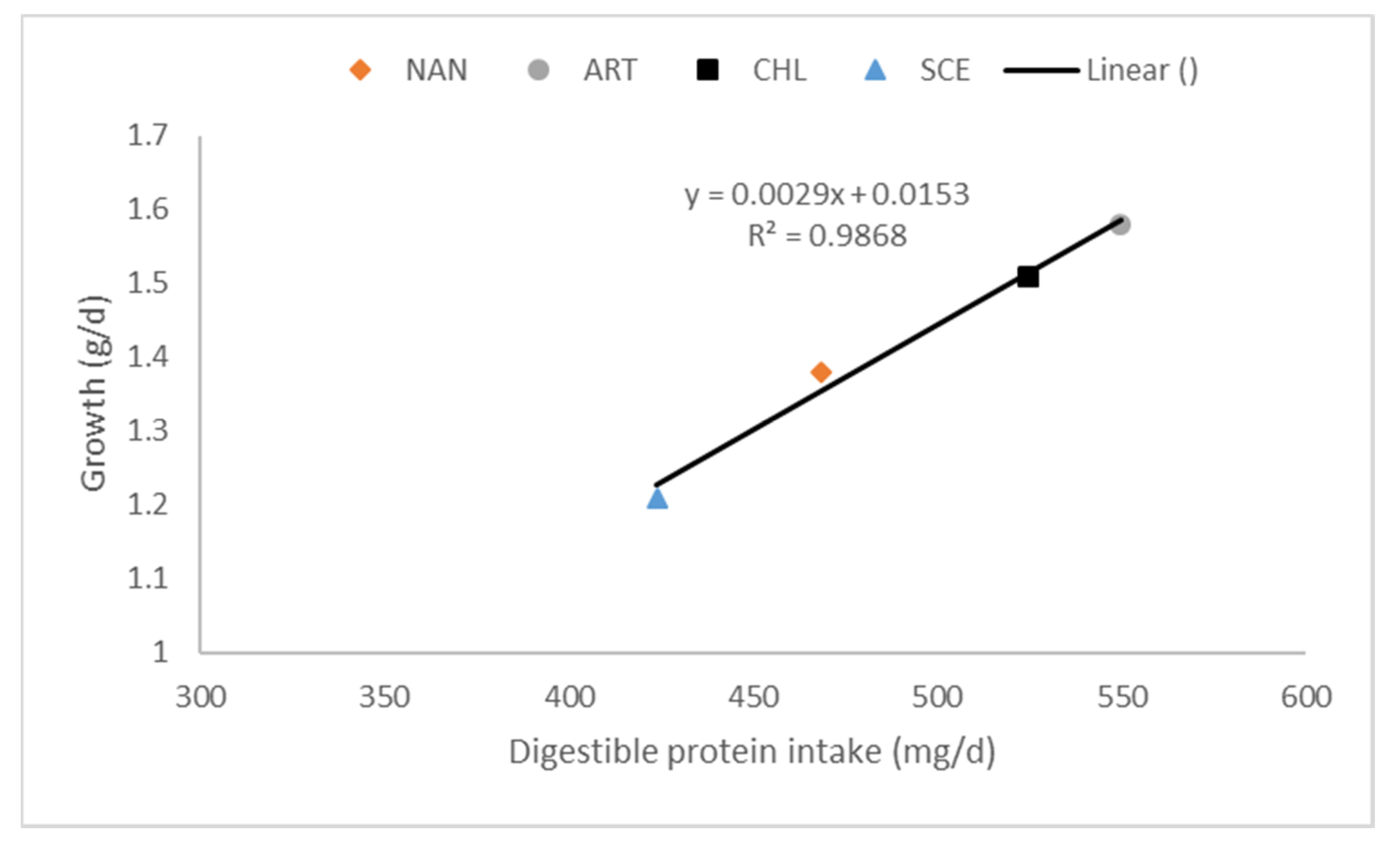
4. Utilisation and Palatability
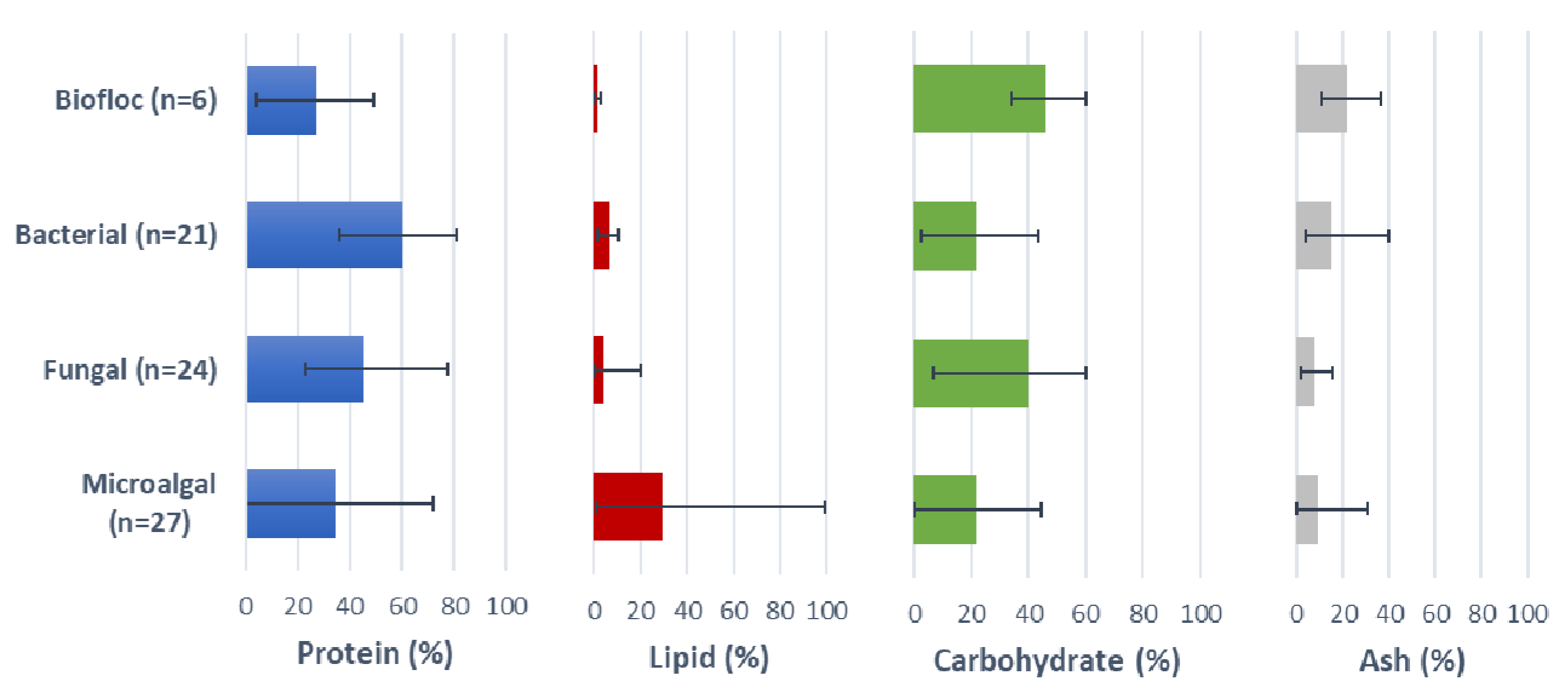
5. Microalgal Resources
5.1. Salmonids
5.2. Shrimp
5.3. Tilapia
5.4. Marine Species
6. Fungal Resources
7. Salmonids
7.1. Shrimp
7.2. Tilapia
7.3. Marine Species
8. Bacterial Resources
8.1. Salmonids
8.2. Shrimp
8.3. Tilapia
8.4. Marine Species
9. Biofloc Resources
9.1. Shrimp
9.2. Tilapia
9.3. Salmonids and Marine Species
10. Immunological and Health Allied Assessments
11. Processing Effects (Functionality)
12. Product Quality Influences
13. Next Steps
Author Contributions
Funding
Conflicts of Interest
References
- D’Mello, J.P.F. A Study of the Amino Acid Composition of Methane Utilizing Bacteria. J. Appl. Bacteriol. 1972, 35, 145–148. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F. The use of methane-utilising bacteria as a source of protein for young chicks1. Br. Poult. Sci. 1973, 14, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Matty, A.; Smith, P. Evaluation of a yeast, a bacterium and an alga as a protein source for rainbow trout: I. Effect of protein level on growth, gross conversion efficiency and protein conversion efficiency. Aquaculture 1978, 14, 235–246. [Google Scholar] [CrossRef]
- Kaushik, S.; Luquet, P. Influence of bacterial protein incorporation and of sulphur amino acid supplementation to such diets on growth of rainbow trout, Salmo gairdnerii Richardson. Aquaculture 1980, 19, 163–175. [Google Scholar] [CrossRef]
- Jones, S.W.; Karpol, A.; Friedman, S.; Maru, B.T.; Tracy, B.P. Recent advances in single cell protein use as a feed ingredient in aquaculture. Curr. Opin. Biotechnol. 2020, 61, 189–197. [Google Scholar] [CrossRef]
- Guedes, A.C.; Malcata, F.X. Nutritional value and uses of microalgae in aquaculture. Aquaculture 2012, 10, 59–78. [Google Scholar]
- Cohen, Z.; Ratledge, C. (Eds.) Single Cell Oils: Microbial and Algal Oils, 2nd ed.; AOCS Press: Urbana, IL, USA, 2010; p. 516. [Google Scholar]
- Øverland, M.; Tauson, A.-H.; Shearer, K.; Skrede, A. Evaluation of methane-utilising bacteria products as feed ingredients for monogastric animals. Arch. Anim. Nutr. 2010, 64, 171–189. [Google Scholar] [CrossRef]
- Tibbetts, S.M. The Potential for ‘Next-Generation’, Microalgae-Based Feed Ingredients for Salmonid Aquaculture in Context of the Blue Revolution. In Microalgal Biotechnology; IntechOpen: London, UK, 2018. [Google Scholar]
- Glencross, B.D.; Baily, J.; Berntssen, M.H.; Hardy, R.; MacKenzie, S.; Tocher, D.R. Risk assessment of the use of alternative animal and plant raw material resources in aquaculture feeds. Rev. Aquac. 2019, 12, 703–758. [Google Scholar] [CrossRef]
- Lee, Y.-K. Microalgal mass culture systems and methods: Their limitation and potential. Environ. Biol. Fishes 2001, 13, 307–315. [Google Scholar] [CrossRef]
- Knuckey, R.M.; Brown, M.R.; Robert, R.; Frampton, D.M. Production of microalgal concentrates by flocculation and their assessment as aquaculture feeds. Aquac. Eng. 2006, 35, 300–313. [Google Scholar] [CrossRef]
- Huntley, M.E.; Johnson, Z.; Brown, S.L.; Sills, D.L.; Gerber, L.; Archibald, I.; Machesky, S.C.; Granados, J.; Beal, C.; Greene, C. Demonstrated large-scale production of marine microalgae for fuels and feed. Algal Res. 2015, 10, 249–265. [Google Scholar] [CrossRef]
- Teuling, E.; Schrama, J.; Gruppen, H.; Wierenga, P.A. Characterizing emulsion properties of microalgal and cyanobacterial protein isolates. Algal Res. 2019, 39, 101471. [Google Scholar] [CrossRef]
- Teuling, E.; Wierenga, P.A.; Agboola, J.O.; Gruppen, H.; Schrama, J. Cell wall disruption increases bioavailability of Nannochloropsis gaditana nutrients for juvenile Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 499, 269–282. [Google Scholar] [CrossRef]
- Emerenciano, M.; Gaxiola, G.; Cuzon, G. Biofloc Technology (BFT): A Review for Aquaculture Application and Animal Food Industry. In Biomass Now—Cultivation and Utilization; IntechOpen: London, UK, 2013; pp. 301–328. [Google Scholar] [CrossRef]
- Bossier, P.; Ekasari, J. Biofloc technology application in aquaculture to support sustainable development goals. Microb. Biotechnol. 2017, 10, 1012–1016. [Google Scholar] [CrossRef]
- Glencross, B.D. A feed is still only as good as its ingredients—An update on the nutritional research strategies for optimal evaluation of ingredients for aquaculture feeds. Aquac. Nutr. 2020, 13, 17–34. [Google Scholar] [CrossRef]
- Glencross, B.D.; Booth, M.; Allan, G.L. A feed is only as good as its ingredients—A review of ingredient evaluation for aquaculture feeds. Aquac. Nutr. 2007, 13, 17–34. [Google Scholar] [CrossRef]
- Atalah, E.; Hernandez-Cruz, C.M.; Izquierdo, M.S.; Rosenlund, G.; Caballero, M.J.; Valencia, A.; Robaina, L. Two microalgae Crypthecodinium cohnii and Phaeodactylum tricornutum as alternative source of essential fatty acids in starter feeds for seabream (Sparus aurata). Aquaculture 2007, 270, 178–185. [Google Scholar] [CrossRef]
- Popovich, C.A.; Pistonesi, M.; Hegel, P.; Constenla, D.; Bielsa, G.B.; Martín, L.A.; Damiani, M.C.; Leonardi, P.I. Unconventional alternative biofuels: Quality assessment of biodiesel and its blends from marine diatom Navicula cincta. Algal Res. 2019, 39, 101438. [Google Scholar] [CrossRef]
- Cardona, E.; Gueguen, Y.; Magré, K.; Lorgeoux, B.; Piquemal, D.; Pierrat, F.; Noguier, F.; Saulnier, D. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar] [CrossRef]
- Pérez-Fuentes, J.A.; Pérez-Rostro, C.I.; Hernández-Vergara, M.P.; Monroy-Dosta, M.D.C. Variation of the bacterial composition of biofloc and the intestine of Nile tilapia Oreochromis niloticus, cultivated using biofloc technology, supplied different feed rations. Aquac. Res. 2018, 49, 3658–3668. [Google Scholar] [CrossRef]
- Galán, B.; Santos-Merino, M.; Nogales, J.; De la Cruz, F.; García, J.L. Microbial Oils as Nutraceuticals and Animal Feeds. In Health Consequences of Microbial Interactions with Hydrocarbons, Oils, and Lipids; Goldfine, H., Ed.; Springer Nature: Cham, Switzerland, 2019; pp. 401–445. [Google Scholar]
- Patil, V.; Reitan, K.I.; Knutsen, G.; Mortensen, L.M.; Källqvist, T.; Olsen, E.; Gislerød, H.R. Microalgae as source of polyunsaturated fatty acids for aquaculture. Plant Biol. 2005, 6, 57–65. [Google Scholar]
- Hemaiswarya, S.; Raja, R.; Kumar, R.R.; Ganesan, V.; Anbazhagan, C. Microalgae: A sustainable feed source for aquaculture. World J. Microbiol. Biotechnol. 2010, 27, 1737–1746. [Google Scholar] [CrossRef]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Chowdhury, M.A.K.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in aquafeeds for a sustainable aquaculture industry. Environ. Biol. Fishes 2017, 30, 197–213. [Google Scholar] [CrossRef]
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst. 2005, 1, 2. [Google Scholar] [CrossRef]
- Pisal, D.S.; Lele, S.S. Carotenoid production from microalga, Dunaliella salina. Indian J. Biotechnol. 2005, 4, 476–483. [Google Scholar]
- Moomaw, W.R.; Berzin, I.; Tzachor, A. Cutting Out the Middle Fish: Marine Microalgae as the Next Sustainable Omega-3 Fatty Acids and Protein Source. Ind. Biotechnol. 2017, 13, 234–243. [Google Scholar] [CrossRef]
- Hamilton, H.A.; Newton, R.; Auchterlonie, N.A.; Müller, D.B. Systems approach to quantify the global omega-3 fatty acid cycle. Nat. Food 2020, 1, 59–62. [Google Scholar] [CrossRef]
- Dunstan, G.A.; Volkman, J.K.; Barrett, S.M.; Garland, C.D. Changes in the lipid composition and maximisation of the polyunsaturated fatty acid content of three microalgae grown in mass culture. Environ. Biol. Fishes 1993, 5, 71–83. [Google Scholar] [CrossRef]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Scaife, M.A.; Armenta, R.E. Apparent digestibility of proximate nutrients, energy and fatty acids in nutritionally-balanced diets with partial or complete replacement of dietary fish oil with microbial oil from a novel Schizochytrium sp.(T18) by juvenile Atlantic salmon (Salmo salar L.). Aquaculture 2020, 520, 735003. [Google Scholar]
- Skrede, A.; Mydland, L.; Ahlstrøm, Ø.; Reitan, K.; Gislerød, H.; Overland, M. Evaluation of microalgae as sources of digestible nutrients for monogastric animals. J. Anim. Feed Sci. 2011, 20, 131–142. [Google Scholar] [CrossRef]
- Sarker, P.; Kapuscinski, A.R.; Lanois, A.J.; Livesey, E.D.; Bernhard, K.P.; Coley, M.L. Towards Sustainable Aquafeeds: Complete Substitution of Fish Oil with Marine Microalga Schizochytrium sp. Improves Growth and Fatty Acid Deposition in Juvenile Nile Tilapia (Oreochromis niloticus). PLoS ONE 2016, 11, e0156684. [Google Scholar] [CrossRef] [PubMed]
- Soto-Sierra, L.; Stoykova, P.; Nikolov, Z.L. Extraction and fractionation of microalgae-based protein products. Algal Res. 2018, 36, 175–192. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, X.; Ji, L.; Song, X.; Kuang, C. Changes of lipid content and fatty acid composition of Schizochytrium limacinum in response to different temperatures and salinities. Process. Biochem. 2007, 42, 210–214. [Google Scholar] [CrossRef]
- Øverland, M.; Skrede, A. Yeast derived from lignocellulosic biomass as a sustainable feed resource for use in aquaculture. J. Sci. Food Agric. 2016, 97, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Delgado, J.; Marquez-Reyes, J.M. Potential of microbial-derived nutrients for aquaculture development. Rev. Aquac. 2018, 10, 224–246. [Google Scholar] [CrossRef]
- Huyben, D.; Nyman, A.; Vidakovic, A.; Passoth, V.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Effects of dietary inclusion of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus on gut microbiota of rainbow trout. Aquaculture 2017, 473, 528–537. [Google Scholar] [CrossRef]
- Nalage, D.; Khedkar, G.; Kalyankar, A.; Sarkate, A.; Ghodke, S. Single cell proteins. In Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 4, pp. 790–794. [Google Scholar]
- Shurson, G. Yeast and yeast derivatives in feed additives and ingredients: Sources, characteristics, animal responses, and quantification methods. Anim. Feed Sci. Technol. 2018, 235, 60–76. [Google Scholar] [CrossRef]
- Vidakovic, A.; Langeland, M.; Sundh, H.; Sundell, K.; Olstorpe, M.; Vielma, J.; Kiessling, A.; Lundh, T. Evaluation of growth performance and intestinal barrier function in Arctic Charr (Salvelinus alpinus) fed yeast (Saccharomyces cerevisiae), fungi (Rhizopus oryzae) and blue mussel (Mytilus edulis). Aquac. Nutr. 2015, 22, 1348–1360. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Esteban, M.Á.; Cuesta, A.; Sun, Y.Z. Prebiotics and fish immune response: A review of current knowledge and future perspectives. Rev. Fish. Sci. Aquac. 2015, 23, 315–328. [Google Scholar] [CrossRef]
- Skrede, A.; Berge, G.; Storebakken, T.; Herstad, O.; Aarstad, K.; Sundstøl, F. Digestibility of bacterial protein grown on natural gas in mink, pigs, chicken and Atlantic salmon. Anim. Feed Sci. Technol. 1998, 76, 103–116. [Google Scholar] [CrossRef]
- Parsons, J.B.; Rock, C.O. Bacterial lipids: Metabolism and membrane homeostasis. Prog. Lipid Res. 2013, 52, 249–276. [Google Scholar] [CrossRef] [PubMed]
- Garay, L.A.; Boundy-Mills, K.L.; German, J.B. Accumulation of High-Value Lipids in Single-Cell Microorganisms: A Mechanistic Approach and Future Perspectives. J. Agric. Food Chem. 2014, 62, 2709–2727. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.; Roth, O.; De Schryver, P.; Bayer, T.; Garcia-Gonzalez, L.; Künzel, S.; Bossier, P.; Miest, J.J.; Clemmesen, C. Poly-β-hydroxybutyrate administration during early life: Effects on performance, immunity and microbial community of European sea bass yolk-sac larvae. Sci. Rep. 2017, 7, 15022. [Google Scholar] [CrossRef]
- Gouveia, L.; Choubert, G.; Pereira, N.; Santinha, J.; Empis, J.; Gomes, E. Pigmentation of gilthead seabream, Sparus aurata (L. 1875), using Chlorella vulgaris (Chlorophyta, Volvocales) microalga. Aquac. Res. 2002, 33, 987–993. [Google Scholar] [CrossRef]
- Waldenstedt, L.; Inborr, J.; Hansson, I.; Elwinger, K. Effects of astaxanthin-rich algal meal (Haematococcus pluvalis) on growth performance, caecal campylobacter and clostridial counts and tissue astaxanthin concentration of broiler chickens. Anim. Feed Sci. Technol. 2003, 108, 119–132. [Google Scholar] [CrossRef]
- Lewis, T.E.; Nichols, P.D.; McMeekin, T.A. The Biotechnological Potential of Thraustochytrids. Surg. Endosc. 1999, 1, 580–587. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, D.; Barrow, C.J.; Puri, M. Exploring potential use of Australian thraustochytrids for the bioconversion of glycerol to omega-3 and carotenoids production. Biochem. Eng. J. 2013, 78, 11–17. [Google Scholar] [CrossRef]
- McGinnis, K.M.; Dempster, T.A.; Sommerfeld, M.R. Characterization of the growth and lipid content of the diatom Chaetoceros muelleri. Environ. Biol. Fishes 1997, 9, 19–24. [Google Scholar] [CrossRef]
- Popovich, C.A.; Damiani, C.; Constenla, D.; Leonardi, P.I. Lipid quality of the diatoms Skeletonema costatum and Navicula gregaria from the South Atlantic Coast (Argentina): Evaluation of its suitability as biodiesel feedstock. Environ. Biol. Fishes 2011, 24, 1–10. [Google Scholar] [CrossRef]
- Ponis, E.; Parisi, G.; Le Coz, J.-R.; Robert, R.; Zittelli, G.; Tredici, M. Effect of the culture system and culture technique on biochemical characteristics of Pavlova lutheri and its nutritional value for Crassostrea gigas larvae. Aquac. Nutr. 2006, 12, 322–329. [Google Scholar] [CrossRef]
- Coutinho, P.; Rema, P.; Otero, A.; Pereira, O.; Fábregas, J. Use of biomass of the marine microalga Isochrysis galbana in the nutrition of goldfish (Carassius auratus) larvae as source of protein and vitamins. Aquac. Res. 2006, 37, 793–798. [Google Scholar] [CrossRef]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae yeasts as protein sources in diets for Atlantic salmon (Salmo salar). Aquaculture 2013, 402, 1–7. [Google Scholar] [CrossRef]
- Vidakovic, A.; Huyben, D.; Sundh, H.; Nyman, A.; Vielma, J.; Passoth, V.; Kiessling, A.; Lundh, T. Growth performance, nutrient digestibility and intestinal morphology of rainbow trout (Oncorhynchus mykiss) fed graded levels of the yeasts Saccharomyces cerevisiae and Wickerhamomyces anomalus. Aquac. Nutr. 2019, 26, 275–286. [Google Scholar] [CrossRef]
- Sakuradani, E.; Shimizu, S. Single cell oil production by Mortierella alpina. J. Biotechnol. 2009, 144, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Olvera-Novoa, M.A.; Dominguez-Cen, L.J.; Olivera-Castillo, L.; A Martinez-Palacios, C. Effect of the use of the microalga Spirulina maxima as fish meal replacement in diets for tilapia, Oreochromis mossambicus (Peters), fry. Aquac. Res. 1998, 29, 709–715. [Google Scholar] [CrossRef]
- Storebakken, T.; Baeverfjord, G.; Skrede, A.; Olli, J.J.; Berge, G.M. Bacterial protein grown on natural gas in diets for Atlantic salmon, Salmo salar, in freshwater. Aquaculture 2004, 241, 413–425. [Google Scholar] [CrossRef]
- Windass, J.D.; Worsey, M.J.; Pioli, E.M.; Pioli, D.; Barth, P.T.; Atherton, K.T.; Dart, E.C.; Byrom, D.; Powell, K.; Senior, P.J. Improved conversion of methanol to single-cell protein by Methylophilus methylotrophus. Nature 1980, 287, 396–401. [Google Scholar] [CrossRef]
- Hardy, R.W.; Patro, B.; Pujol-Baxley, C.; Marx, C.J.; Feinberg, L. Partial replacement of soybean meal with Methylobacterium extorquens single-cell protein in feeds for rainbow trout (Oncorhynchus mykiss Walbaum). Aquac. Res. 2018, 49, 2218–2224. [Google Scholar] [CrossRef]
- Schneider, O.; Sereti, V.; Machiels, M.A.; Eding, E.H.; Verreth, J.A.J. The potential of producing heterotrophic bacteria biomass on aquaculture waste. Water Res. 2006, 40, 2684–2694. [Google Scholar] [CrossRef]
- Moss, S.M.; Pruder, G.D. Characterization of organic particles associated with rapid growth in juvenile white shrimp, Penaeus vannamei Boone, reared under intensive culture conditions. J. Exp. Mar. Biol. Ecol. 1995, 187, 175–191. [Google Scholar] [CrossRef]
- Ekasari, J.; Azhar, M.; Surawidjaja, E.H.; Nuryati, S.; De Schryver, P.; Bossier, P. Immune response and disease resistance of shrimp fed biofloc grown on different carbon sources. Fish Shellfish. Immunol. 2014, 41, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Burford, M.A.; Thompson, P.J.; McIntosh, R.P.; Bauman, R.H.; Pearson, D.C. The contribution of flocculated material to shrimp (Litopenaeus vannamei) nutrition in a high-intensity, zero-exchange system. Aquaculture 2004, 232, 525–537. [Google Scholar] [CrossRef]
- Avnimelech, Y. Feeding with microbial flocs by tilapia in minimal discharge bio-flocs technology ponds. Aquaculture 2007, 264, 140–147. [Google Scholar] [CrossRef]
- Ballester, E.L.C.; Abreu, P.; Cavalli, R.O.; Emerenciano, M.; De Abreu, L. Effect of practical diets with different protein levels on the performance of Farfantepenaeus paulensis juveniles nursed in a zero exchange suspended microbial flocs intensive system. Aquac. Nutr. 2010, 16, 163–172. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Ballester, E.L.C.; Cavalli, R.O. Biofloc technology application as a food source in a limited water exchange nursery system for pink shrimp Farfantepenaeus brasiliensis (Latreille, 1817). Aquac. Res. 2011, 43, 447–457. [Google Scholar] [CrossRef]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 356, 351–356. [Google Scholar] [CrossRef]
- Tago, Y.; Aida, K. Exocellular mucopolysaccharide closely related to bacterial floc formation. Appl. Environ. Microbiol. 1977, 34, 308–314. [Google Scholar] [CrossRef]
- Kuhn, D.D.; Boardman, G.D.; Lawrence, A.L.; Marsh, L.; Flick, G.J. Microbial floc meal as a replacement ingredient for fish meal and soybean protein in shrimp feed. Aquaculture 2009, 296, 51–57. [Google Scholar] [CrossRef]
- Glencross, B.; Irvin, S.; Arnold, S.; Blyth, D.; Bourne, N.; Preston, N. Effective use of microbial biomass products to facilitate the complete replacement of fishery resources in diets for the black tiger shrimp, Penaeus monodon. Aquaculture 2014, 431, 12–19. [Google Scholar] [CrossRef]
- Glencross, B.; Arnold, S.; Irvin, S. Bioactive factors in microbial biomass have the capacity to offset reductions in the level of protein in the diet of black tiger shrimp, Penaeus monodon. Aquaculture 2015, 446, 74–79. [Google Scholar] [CrossRef]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takkriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. 2014, 21, 6. [Google Scholar] [CrossRef]
- Ende, S.S.; Noke, A. Heterotrophic microalgae production on food waste and by-products. J. Appl. Phycol. 2019, 31, 1565–1571. [Google Scholar] [CrossRef]
- Sharma, S.; Hansen, L.D.; Hansen, J.Ø.; Mydland, L.T.; Horn, S.J.; Øverland, M.; Eijsink, V.G.H.; Vuoristo, K.S. Microbial Protein Produced from Brown Seaweed and Spruce Wood as a Feed Ingredient. J. Agric. Food Chem. 2018, 66, 8328–8335. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Garland, C.D.; Jeffrey, S.W.; Jameson, I.D.; Leroi, J.M. The gross and amino acid compositions of batch and semi-continuous cultures of Isochrysis sp. (clone T. ISO), Pavlova lutheri and Nannochloropsis oculata. J. Appl. Phycol. 1993, 5, 285–296. [Google Scholar] [CrossRef]
- Huerlimann, R.; De Nys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 107, 245–257. [Google Scholar] [CrossRef]
- Chua, E.T.; Schenk, P.M. A biorefinery for Nannochloropsis: Induction, harvesting, and extraction of EPA-rich oil and high-value protein. Bioresour. Technol. 2017, 244, 1416–1424. [Google Scholar] [CrossRef]
- Hatlen, B.; Berge, G.M.; Odom, J.M.; Mundheim, H.; Ruyter, B. Growth performance, feed utilisation and fatty acid deposition in Atlantic salmon, Salmo salar L.; fed graded levels of high-lipid/high-EPA Yarrowia lipolytica biomass. Aquaculture 2012, 364, 39–47. [Google Scholar] [CrossRef]
- Xie, D.; Jackson, E.N.; Zhu, Q. Sustainable source of omega-3 eicosapentaenoic acid from metabolically engineered Yarrowia lipolytica: From fundamental research to commercial production. Appl. Microbiol. Biotechnol. 2015, 99, 1599–1610. [Google Scholar] [CrossRef]
- Katerina, K.; Mørkøre, T.; Nengas, I.; Berge, R.; Sweetman, J. Microalgae and organic minerals enhance lipid retention efficiency and fillet quality in Atlantic salmon (Salmo salar L.). Aquaculture 2016, 451, 47–57. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Yasumaru, F.; Lemos, D. In vitro prediction of digestible protein content of marine microalgae (Nannochloropsis granulata) meals for Pacific white shrimp (Litopenaeus vannamei) and rainbow trout (Oncorhynchus mykiss). Algal Res. 2017, 21, 76–80. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.-V.; Parry, D.L. The gross chemical composition and fatty acid composition of 18 species of tropical Australian microalgae for possible use in mariculture. Aquaculture 1999, 170, 147–159. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Ghosh, A.; Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.; Saini, S.; Bhowmick, T.K.; Gayen, K. Progress toward isolation of strains and genetically engineered strains of microalgae for production of biofuel and other value added chemicals: A review. Energy Convers. Manag. 2016, 113, 104–118. [Google Scholar] [CrossRef]
- Camacho-Rodríguez, J.; Macías-Sánchez, M.D.; Cerón-García, M.; Alarcón, F.J.; Molina-Grima, E. Microalgae as a potential ingredient for partial fish meal replacement in aquafeeds: Nutrient stability under different storage conditions. Environ. Biol. Fishes 2017, 30, 1049–1059. [Google Scholar] [CrossRef]
- Lorenz, R.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Shah, M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-Producing Green Microalga Haematococcus pluvialis: From Single Cell to High Value Commercial Products. Front. Plant Sci. 2016, 7, 300. [Google Scholar] [CrossRef]
- Sanderson, G.W.; Jolly, S.O. The value of Phaffia yeast as a feed ingredient for salmonid fish. Aquaculture 1994, 124, 193–200. [Google Scholar] [CrossRef]
- Li, P.; Gatlin, D.M. Nucleotide nutrition in fish: Current knowledge and future applications. Aquaculture 2006, 251, 141–152. [Google Scholar] [CrossRef]
- Casadei, E.; Bird, S.; Vecino, J.L.G.; Wadsworth, S.; Secombes, C.J. The effect of peptidoglycan enriched diets on antimicrobial peptide gene expression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2013, 34, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Casadei, E.; Bird, S.; Wadsworth, S.; Vecino, J.L.G.; Secombes, C.J. The longevity of the antimicrobial response in rainbow trout (Oncorhynchus mykiss) fed a peptidoglycan (PG) supplemented diet. Fish Shellfish. Immunol. 2015, 44, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Sellars, M.; Rao, M.; Polymeris, N.; Irvin, S.J.; Cowley, J.A.; Preston, N.P.; Glencross, B.D. Feed Containing Novacq Improves Resilience of Black Tiger Shrimp, Penaeus Monodon, to Gill-associated Virus-induced Mortality. J. World Aquac. Soc. 2015, 46, 328–336. [Google Scholar] [CrossRef]
- Gong, Y.; Guterres, H.; Huntley, M.; Sørensen, M.; Kiron, V. Digestibility of the defatted microalgae Nannochloropsis sp. and Desmodesmus sp. when fed to Atlantic salmon, Salmo salar. Aquac. Nutr. 2017, 24, 56–64. [Google Scholar] [CrossRef]
- Raji, A.A.; Jimoh, W.A.; Abu Bakar, N.H.; Taufek, N.H.M.; Muin, H.; Alias, Z.; Milow, P.; Razak, S.A. Dietary use of Spirulina (Arthrospira) and Chlorella instead of fish meal on growth and digestibility of nutrients, amino acids and fatty acids by African catfish. Environ. Biol. Fishes 2020, 1–8. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Vandenberg, G.W.; Proulx, E.; Sitek, A.J. Towards sustainable and ocean-friendly aquafeeds: Evaluating a fish-free feed for rainbow trout (Oncorhynchus mykiss) using three marine microalgae species. Elem. Sci. Anth. 2020, 8. [Google Scholar] [CrossRef]
- Teuling, E.; Schrama, J.; Gruppen, H.; Wierenga, P.A. Effect of cell wall characteristics on algae nutrient digestibility in Nile tilapia (Oreochromis niloticus) and African catfish (Clarus gariepinus). Aquaculture 2017, 479, 490–500. [Google Scholar] [CrossRef]
- Agboola, J.O.; Teuling, E.; Wierenga, P.A.; Gruppen, H.; Schrama, J. Cell wall disruption: An effective strategy to improve the nutritive quality of microalgae in African catfish (Clarias gariepinus). Aquac. Nutr. 2019, 25, 783–797. [Google Scholar] [CrossRef]
- Rumsey, G.; Kinsella, J.; Shetty, K.; Hughes, S. Effect of high dietary concentrations of brewer’s dried yeast on growth performance and liver uricase in rainbow trout (Oncorhynchus mykiss). Anim. Feed Sci. Technol. 1991, 33, 177–183. [Google Scholar] [CrossRef]
- Langeland, M.; Vidakovic, A.; Vielma, J.; Lindberg, J.E.; Kiessling, A.; Lundh, T. Digestibility of microbial and mussel meal for Arctic charr (Salvelinus alpinus) and Eurasian perch (Perca fluviatilis). Aquac. Nutr. 2016, 22, 485–495. [Google Scholar] [CrossRef]
- Storebakken, T.; Kvien, I.; Shearer, K.; Grisdale-Helland, B.; Helland, S.; Berge, G. The apparent digestibility of diets containing fish meal, soybean meal or bacterial meal fed to Atlantic salmon (Salmo salar): Evaluation of different faecal collection methods. Aquaculture 1998, 169, 195–210. [Google Scholar] [CrossRef]
- Aas, T.S.; Grisdale-Helland, B.; Terjesen, B.F.; Helland, S.J. Improved growth and nutrient utilisation in Atlantic salmon (Salmo salar) fed diets containing a bacterial protein meal. Aquaculture 2006, 259, 365–376. [Google Scholar] [CrossRef]
- Øverland, M.; Romarheim, O.H.; Hovin, M.; Storebakken, T.; Skrede, A. Apparent total tract digestibility of unprocessed and extruded diets containing basic and autolyzed bacterial protein meal grown on natural gas in mink and rainbow trout. Anim. Feed Sci. Technol. 2006, 129, 237–251. [Google Scholar]
- Ekasari, J.; Suprayudi, M.A.; Elas, P.; Senja, R.K. The digestibility of biofloc meal from African catfish culture medium as a feed raw material for Pacific white shrimp. J. Akuakultur Indones. 2019, 18, 1–8. [Google Scholar] [CrossRef]
- Neto, H.S.; Santaella, S.T.; Nunes, A.J.P. Bioavailability of crude protein and lipid from biofloc meals produced in an activated sludge system for white shrimp, Litopenaeus vannamei. Rev. Bras. de Zootec. 2015, 44, 269–275. [Google Scholar] [CrossRef]
- Liu, W.; Luo, G.; Tan, H.; Sun, D. Effects of sludge retention time on water quality and bioflocs yield, nutritional composition, apparent digestibility coefficients treating recirculating aquaculture system effluent in sequencing batch reactor. Aquac. Eng. 2016, 72, 58–64. [Google Scholar] [CrossRef]
- Carter, C.G.; Bransden, M.P.; Lewis, T.E.; Nichols, P.D. Potential of thraustochytrids to partially replace fish oil in Atlantic salmon feeds. Mar. Biotechnol. 2003, 5, 480–492. [Google Scholar] [CrossRef]
- Miller, M.; Nichols, P.D.; Carter, C.G. Replacement of fish oil with thraustochytrid Schizochytrium sp. L oil in Atlantic salmon parr (Salmo salar L.) diets. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 148, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Katerina, K.; Østbye, T.-K.K.; Krasnov, A.; Torgersen, J.S.; Mørkøre, T.; Sweetman, J. Metabolism, health and fillet nutritional quality in Atlantic salmon (Salmo salar) fed diets containing n-3-rich microalgae. J. Nutr. Sci. 2015, 4. [Google Scholar] [CrossRef]
- Katerina, K.; Berge, G.M.; Turid, M.; Aleksei, K.; Grete, B.; Trine, Y.; Mats, C.; John, S.; Bente, R. Microalgal Schizochytrium limacinum Biomass Improves Growth and Filet Quality When Used Long-Term as a Replacement for Fish Oil, in Modern Salmon Diets. Front. Mar. Sci. 2020, 7, 57. [Google Scholar] [CrossRef]
- Betiku, O.C.; Barrows, F.T.; Ross, C.; Sealey, W.M. The effect of total replacement of fish oil with DHA-Gold® and plant oils on growth and fillet quality of rainbow trout (Oncorhynchus mykiss) fed a plant-based diet. Aquac. Nutr. 2016, 22, 158–169. [Google Scholar] [CrossRef]
- Sørensen, M.; Berge, G.M.; Reitan, K.I.; Ruyter, B. Microalga Phaeodactylum tricornutum in feed for Atlantic salmon (Salmo salar)—Effect on nutrient digestibility, growth and utilization of feed. Aquaculture 2016, 460, 116–123. [Google Scholar] [CrossRef]
- Schøyen, H.F.; Frøyland, J.R.K.; Sahlström, S.; Knutsen, S.H.; Skrede, A. Effects of autolysis and hydrolysis of bacterial protein meal grown on natural gas on chemical characterization and amino acid digestibility. Aquaculture 2005, 248, 27–33. [Google Scholar] [CrossRef]
- Yarnold, J.; Karan, H.; Oey, M.; Hankamer, B. Microalgal Aquafeeds As Part of a Circular Bioeconomy. Trends Plant Sci. 2019, 24, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Turchini, G.; Torstensen, B.E.; Ng, W.-K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Sprague, M.; Walton, J.; Campbell, P.; Strachan, F.; Dick, J.; Bell, J. Replacement of fish oil with a DHA-rich algal meal derived from Schizochytrium sp. on the fatty acid and persistent organic pollutant levels in diets and flesh of Atlantic salmon (Salmo salar, L.) post-smolts. Food Chem. 2015, 185, 413–421. [Google Scholar] [CrossRef]
- Chang, K.J.; Parrish, C.C.; Simon, C.J.; Revill, A.T.; Nichols, P.D. Feeding Whole Thraustochytrid Biomass to Cultured Atlantic Salmon (Salmo salar) Fingerlings: Culture Performance and Fatty Acid Incorporation. J. Mar. Sci. Eng. 2020, 8, 207. [Google Scholar] [CrossRef]
- Norambuena, F.; Hermon, K.; Skrzypczyk, V.; Emery, J.A.; Sharon, Y.; Beard, A.; Turchini, G. Algae in Fish Feed: Performances and Fatty Acid Metabolism in Juvenile Atlantic Salmon. PLoS ONE 2015, 10, e0124042. [Google Scholar] [CrossRef]
- Patnaik, S.; Samocha, T.M.; Davis, D.; Bullis, R.; Browdy, C. The use of HUFA-rich algal meals in diets for Litopenaeus vannamei. Aquac. Nutr. 2006, 12, 395–401. [Google Scholar] [CrossRef]
- Samocha, T.; Patnaik, S.; Davis, D.A.; Bullis, R.A.; Browdy, C.L. Use of commercial fermentation products as a highly unsaturated fatty acid source in practical diets for the Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2009, 41, 961–967. [Google Scholar] [CrossRef]
- Samocha, T.; Davis, D.; Roy, L.; Carpenter, B.; Bullis, R. The effect of non-marine HUFA supplementation with fish oil removal on growth and survival of the Pacific white shrimp, Litopenaeus vannamei. Aquac. Nutr. 2010, 17, 518–525. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Deng, D.F.; Dominy, W. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 2012, 354, 50–55. [Google Scholar] [CrossRef]
- Hende, S.V.D.; Claessens, L.; De Muylder, E.; Boon, N.; Vervaeren, H. Microalgal bacterial flocs originating from aquaculture wastewater treatment as diet ingredient for Litopenaeus vannamei (Boone). Aquac. Res. 2014, 47, 1075–1089. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Filer, K.; Xue, Y.; Ai, Q.; Mai, K. Evaluation of Schizochytrium meal in microdiets of Pacific white shrimp (Litopenaeus vannamei) larvae. Aquac. Res. 2016, 48, 2328–2336. [Google Scholar] [CrossRef]
- Allen, K.M.; Habte-Tsion, H.M.; Thompson, K.R.; Filer, K.; Tidwell, J.H.; Kumar, V. Freshwater microalgae (Schizochytrium sp.) as a substitute to fish oil for shrimp feed. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Araújo, B.C.; Mata-Sotres, J.A.; Viana, M.T.; Tinajero, A.; Braga, A. Fish oil-free diets for Pacific white shrimp Litopenaeus vannamei: The effects of DHA-EPA supplementation on juvenile growth performance and muscle fatty acid profile. Aquaculture 2019, 511, 734276. [Google Scholar] [CrossRef]
- Guimarães, A.M.; Schleder, D.D.; Nagata, M.; Nóbrega, R.O.; Fracalossi, D.M.; Seiffert, W.Q.; Vieira, F.D.N. Aurantiochytrium sp. meal can replace fish oil in practical diets for the juvenile Pacific white shrimp. Aquac. Nutr. 2019, 25, 798–807. [Google Scholar] [CrossRef]
- Adissin, O.; Manabu, I.; Shunsuke, K.; Saichiro, Y.; Moss, A.S.; Dossou, S. Effects of dietary Nannochloropsis sp. powder and lipids on the growth performance and fatty acid composition of larval and postlarval kuruma shrimp, Marsupenaeus japonicus. Aquac. Nutr. 2019, 26, 186–200. [Google Scholar] [CrossRef]
- Basri, N.A.; Shaleh, S.R.M.; Matanjun, P.; Noor, N.M.; Shapawi, R. The potential of microalgae meal as an ingredient in the diets of early juvenile Pacific white shrimp, Litopenaeus vannamei. Environ. Biol. Fishes 2014, 27, 857–863. [Google Scholar] [CrossRef]
- Gbadamosi, O.; Lupatsch, I. Effects of dietary Nannochloropsis salina on the nutritional performance and fatty acid profile of Nile tilapia, Oreochromis niloticus. Algal Res. 2018, 33, 48–54. [Google Scholar] [CrossRef]
- Mahmoud, E.; El-Sayed, B.; Mahsoub, Y.; El-Murr, A.; Neamat-Allah, A. Effect of Chlorella vulgaris enriched diet on growth performance, hemato-immunological responses, antioxidant and transcriptomics profile disorders caused by deltamethrin toxicity in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 102, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Hussein, E.E.-S.; Dabrowski, K.; Lee, B.-J.; El-Saidy, D.M.S.D. Enhancing the growth of Nile tilapia larvae/juveniles by replacing plant (gluten) protein with algae protein. Aquac. Res. 2012, 44, 937–949. [Google Scholar] [CrossRef]
- Hussein, E.E.; Dabrowski, K.; El-Saidy, D.M.S.D.; Lee, B. Effect of dietary phosphorus supplementation on utiization of algae in the grow-out diet of Nile tilapia Oreochromis niloticus. Aquac. Res. 2014, 45, 1533–1544. [Google Scholar] [CrossRef]
- Sarker, P.; Kapuscinski, A.R.; Bae, A.Y.; Donaldson, E.; Sitek, A.J.; Fitzgerald, D.S.; Edelson, O.F. Towards sustainable aquafeeds: Evaluating substitution of fishmeal with lipid-extracted microalgal co-product (Nannochloropsis oculata) in diets of juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0201315. [Google Scholar] [CrossRef]
- Harel, M.; Koven, W.; Lein, I.; Bar, Y.; Behrens, P.; Stubblefield, J.; Zohar, Y.; Place, A.R. Advanced DHA, EPA and ArA enrichment materials for marine aquaculture using single cell heterotrophs. Aquaculture 2002, 213, 347–362. [Google Scholar] [CrossRef]
- Glencross, B.; Rutherford, N. A determination of the quantitative requirements for docosahexaenoic acid for juvenile barramundi (Lates calcarifer). Aquac. Nutr. 2010, 17, e536–e548. [Google Scholar] [CrossRef]
- Eryalçın, K.M.; Roo, J.; Saleh, R.; Atalah, E.; Benítez, T.; Betancor, M.; Hernandez-Cruz, M.D.C.; Izquierdo, M.S.; Eryalçın, K.M.; Hernandez-Cruz, M.C. Fish oil replacement by different microalgal products in microdiets for early weaning of gilthead sea bream (Sparus aurata, L.). Aquac. Res. 2012, 44, 819–828. [Google Scholar] [CrossRef]
- Haas, S.; Bauer, J.L.; Adakli, A.; Meyer, S.; Lippemeier, S.; Schwarz, K.; Schulz, C. Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.). Environ. Biol. Fishes 2015, 28, 1011–1021. [Google Scholar] [CrossRef]
- Ganuza, E.; Benítez-Santana, T.; Atalah, E.; Vega-Orellana, O.M.; Ganga, R.; Izquierdo, M.S. Crypthecodinium cohnii and Schizochytrium sp. as potential substitutes to fisheries-derived oils from seabream (Sparus aurata) microdiets. Aquaculture 2008, 277, 109–116. [Google Scholar] [CrossRef]
- Perez-Velazquez, M.; Gatlin, D.; González-Félix, M.L.; Garcia-Ortega, A. Partial replacement of fishmeal and fish oil by algal meals in diets of red drum Sciaenops ocellatus. Aquaculture 2018, 487, 41–50. [Google Scholar] [CrossRef]
- Nasseri, A.; Rasoul-Ami, S.; Morowvat, M.H.; Ghasemi, Y. Single Cell Protein: Production and Process. Am. J. Food Technol. 2011, 6, 103–116. [Google Scholar] [CrossRef]
- Ravindra, P. Value-added food: Single cell protein. Biotechnol. Adv. 2000, 18, 459–479. [Google Scholar]
- Huyben, D.; Vidakovic, A.; Nyman, A.; Langeland, M.; Lundh, T.; Kiessling, A. Effects of dietary yeast inclusion and acute stress on post-prandial whole blood profiles of dorsal aorta-cannulated rainbow trout. Fish Physiol. Biochem. 2016, 43, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Sun, L.; Moccia, R.; Kiessling, A.; Dicksved, J.; Lundh, T. Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 2018, 124, 1377–1392. [Google Scholar] [CrossRef] [PubMed]
- Mahnken, C.V.; Spinelli, J.; Waknitz, F. Evaluation of an alkane yeast (Candida sp.) as a substitute for fish meal in Oregon Moist Pellet: Feeding trials with coho salmon (Oncorhynchus kisutch) and rainbow trout (Salmo gairdneri). Aquaculture 1980, 20, 41–56. [Google Scholar] [CrossRef]
- Rumsey, G.L.; Hughes, S.G.; Kinsella, J.L. Use of dietary yeast Saccharomyces cerevisiae nitrogen by lake trout. J. World Aquac. Soc. 1990, 21, 205–209. [Google Scholar] [CrossRef]
- Hauptman, B.S.; Barrows, F.T.; Block, S.S.; Gaylord, T.G.; Paterson, J.A.; Rawles, S.D.; Sealey, W.M. Evaluation of grain distillers dried yeast as a fish meal substitute in practical-type diets of juvenile rainbow trout, Oncorhynchus mykiss. Aquaculture 2014, 432, 7–14. [Google Scholar] [CrossRef]
- Sealey, W.M.; O’Neill, T.J.; Peach, J.T.; Gaylord, G.; Barrows, F.T.; Block, S.S. Refining Inclusion Levels of Grain Distiller’s Dried Yeast in Commercial-type and Plant-based Diets for Juvenile Rainbow Trout, Oncorhynchus mykiss. J. World Aquac. Soc. 2015, 46, 434–444. [Google Scholar] [CrossRef]
- Fox, I.H. Metabolic basis for disorders of purine nucleotide degradation. Metabolism 1981, 30, 616–634. [Google Scholar] [CrossRef]
- Clifford, A.J.; Story, D.L. Levels of purines in foods and their metabolic effects in rats. J. Nutr. 1976, 106, 435–442. [Google Scholar] [CrossRef]
- Waslien, C.I.; Calloway, D.H.; Margen, S.; Costa, F. Uric acid levels in men fed algae and yeast as protein sources. J. Food Sci. 1970, 35, 294–298. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; de La Higuera, M.; Varela, G. Alterations of erythrocytes of the rainbow trout (Salmo gairdneri) by the use of Hansenula anomala yeast as sole protein source. Comp. Biochem. Physiol. Part A Physiol. 1982, 72, 693–696. [Google Scholar]
- Huyben, D.; Vidakovic, A.; Langeland, M.; Nyman, A.; Lundh, T.; Kiessling, A. Effects of dietary yeast inclusion and acute stress on postprandial plasma free amino acid profiles of dorsal aorta-cannulated rainbow trout. Aquac. Nutr. 2017, 24, 236–246. [Google Scholar] [CrossRef]
- Blomqvist, J.; Pickova, J.; Tilami, S.K.; Sampels, S.; Mikkelsen, N.; Brandenburg, J.; Sandgren, M.; Passoth, V. Oleaginous yeast as a component in fish feed. Sci. Rep. 2018, 8, 15945. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Boqvist, S.; Passoth, V.; Renström, L.; Bengtsson, U.A.; Andréoletti, O.; Kiessling, A.; Lundh, T.; Vågsholm, I. Screening of intact yeasts and cell extracts to reduce Scrapie prions during biotransformation of food waste. Acta Veter Scand. 2018, 60, 9. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Rajan, D.K. Potential uses of fungal polysaccharides as immunostimulants in fish and shrimp aquaculture: A review. Aquaculture 2019, 500, 250–263. [Google Scholar] [CrossRef]
- Gamboa-Delgado, J.; Fernández-Díaz, B.; Nieto-López, M.G.; Cruz-Suárez, L.E. Nutritional contribution of torula yeast and fish meal to the growth of shrimp Litopenaeus vannamei as indicated by natural nitrogen stable isotopes. Aquaculture 2016, 453, 116–121. [Google Scholar] [CrossRef]
- Achupallas, J.; Zhou, Y.; Davis, D.A. Pond production of Pacific white shrimp, Litopenaeus vannamei, fed grain distillers dried yeast. Aquac. Nutr. 2015, 22, 1222–1229. [Google Scholar] [CrossRef]
- Achupallas, J.M.; Zhou, Y.; Davis, D.A. Use of Grain Distillers Dried Yeast in Practical Diets for Juvenile Pacific White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2016, 47, 220–229. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, W.; Huang, X.; Guo, T.; Wen, W.; Feng, L.; Wei, L. The effect of replacement of fish meal by yeast extract on the digestibility, growth and muscle composition of the shrimp Litopenaeus vannamei. Aquac. Res. 2017, 48, 311–320. [Google Scholar] [CrossRef]
- Qiu, X.; Davis, D.A. Evaluation of flash dried yeast as a nutritional supplement in plant-based practical diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2017, 23, 1244–1253. [Google Scholar] [CrossRef]
- Xiong, J.; Jin, M.; Yuan, Y.; Luo, J.-X.; Lu, Y.; Zhou, Q.-C.; Liang, C.; Tan, Z.-L. Dietary nucleotide-rich yeast supplementation improves growth, innate immunity and intestinal morphology of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1425–1435. [Google Scholar] [CrossRef]
- Li, P.; Wang, X.; Murthy, S.; Gatlin, D.M.; Castille, F.L.; Lawrence, A.L. Effect of Dietary Supplementation of Brewer’s Yeast and GroBiotic®-A on Growth, Immune Responses, and Low-Salinity Tolerance of Pacific White Shrimp Litopenaeus vannamei Cultured in Recirculating Systems. J. Appl. Aquac. 2009, 21, 110–119. [Google Scholar] [CrossRef]
- Babu, D.T.; Antony, S.P.; Joseph, S.P.; Bright, A.R.; Philip, R. Marine yeast Candida aquaetextoris S527 as a potential immunostimulant in black tiger shrimp Penaeus monodon. J. Invertebr. Pathol. 2013, 112, 243–252. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Trinh, L.T.; Chau, D.T.; Baruah, K.; Lundh, T.; Kiessling, A. Spent brewer’s yeast as a replacement for fishmeal in diets for giant freshwater prawn (Macrobrachium rosenbergii), reared in either clear water or a biofloc environment. Aquac. Nutr. 2019, 25, 970–979. [Google Scholar] [CrossRef]
- Hai, N.V. Research findings from the use of probiotics in tilapia aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 592–597. [Google Scholar] [CrossRef]
- Ozório, R.O.A.; Portz, L.; Borghesi, R.; Cyrino, J.E.P. Effects of Dietary Yeast (Saccharomyces cerevisiae) Supplementation in Practical Diets of Tilapia (Oreochromis niloticus). Animals 2012, 2, 16–24. [Google Scholar] [CrossRef]
- Al-Hafedh, Y.S.; Alam, A. Replacement of Fishmeal by Single Cell Protein Derived from Yeast Grown on Date (Phoenix dactylifera) Industry Waste in the Diet of Nile Tilapia (Oreochromis niloticus) Fingerlings. J. Appl. Aquac. 2013, 25, 346–358. [Google Scholar] [CrossRef]
- Fayeofori, G.B.-M.; Bob-Manuel, F.G. A comparative study of the effect of yeast single cell protein on growth, feed utilization and condition factor of the African catfish Clarias gariepinus (Burchell) and tilapia, Oreochromis niloticus (Linnaeus) fingerlings. Afr. J. Agric. Res. 2014, 9, 2005–2011. [Google Scholar] [CrossRef]
- Abass, D.A.; Obirikorang, K.A.; Campion, B.B.; Edziyie, R.E.; Skov, P.V. Dietary supplementation of yeast (Saccharomyces cerevisiae) improves growth, stress tolerance, and disease resistance in juvenile Nile tilapia (Oreochromis niloticus). Aquac. Int. 2018, 26, 843–855. [Google Scholar] [CrossRef]
- Berto, R.D.S.; Pereira, G.D.V.; Mouriño, J.L.P.; Martins, M.L.; Fracalossi, D.M. Yeast extract on growth, nutrient utilization and haemato-immunological responses of Nile tilapia. Aquac. Res. 2015, 47, 2650–2660. [Google Scholar] [CrossRef]
- Trosvik, K.A.; Rawles, S.D.; Thompson, K.R.; Metts, L.A.; Gannam, A.; Twibell, R.; Webster, C. Growth and Body Composition of Nile Tilapia, Oreochromis niloticus, Fry Fed Organic Diets Containing Yeast Extract and Soybean Meal as Replacements for Fish Meal, with and without Supplemental Lysine and Methionine. J. World Aquac. Soc. 2012, 43, 635–647. [Google Scholar] [CrossRef]
- Olvera-Novoa, M.; Martinez-Palacios, C.; Olivera-Castillo, L. Utilization of torula yeast (Candida utilis) as a protein source in diets for tilapia (Oreochromis mossambicus Peters) fry. Aquac. Nutr. 2002, 8, 257–264. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Hasan, M.R.; Metian, M. Demand and supply of feed ingredients for farmed fish and crustaceans: Trends and prospects. FAO Fish. Aquac. Tech. Paper 2011, 564, 1–87. [Google Scholar]
- Nhi, N.H.Y.; Da, C.T.; Lundh, T.; Lan, T.T.; Kiessling, A. Comparative evaluation of Brewer’s yeast as a replacement for fishmeal in diets for tilapia (Oreochromis niloticus), reared in clear water or biofloc environments. Aquaculture 2018, 495, 654–660. [Google Scholar] [CrossRef]
- Ribeiro, C.D.S.; Moreira, R.G.; Cantelmo, O.A.; Esposito, E. The use ofKluyveromyces marxianusin the diet of Red-Stirling tilapia (Oreochromis niloticus, Linnaeus) exposed to natural climatic variation: Effects on growth performance, fatty acids, and protein deposition. Aquac. Res. 2012, 45, 812–827. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Gonçalves, P. Partial replacement of fishmeal by brewers yeast (Saccaromyces cerevisae) in diets for sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2001, 202, 269–278. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Guedes, M.; Vachot, C.; Kaushik, S. The effect of nucleic acids on growth, ureagenesis and nitrogen excretion of gilthead sea bream Sparus aurata juveniles. Aquaculture 2006, 253, 608–617. [Google Scholar] [CrossRef]
- Li, P.; Gatlin, D.M. Evaluation of brewers yeast (Saccharomyces cerevisiae) as a feed supplement for hybrid striped bass (Morone chrysops × M. saxatilis). Aquaculture 2003, 219, 681–692. [Google Scholar] [CrossRef]
- Rosas, V.T.; Poersch, L.; Romano, L.A.; Tesser, M.B. Feasibility of the use of Spirulina in aquaculture diets. Rev. Aquac. 2018, 11, 1367–1378. [Google Scholar] [CrossRef]
- Belay, A.; Kato, T.; Ota, Y. Spirulina (Arthrospira): Potential application as an animal feed supplement. J. Appl. Phycol. 1996, 8, 303–311. [Google Scholar] [CrossRef]
- Matassa, S.; Boon, N.; Pikaar, I.; Verstraete, W. Microbial protein: Future sustainable food supply route with low environmental footprint. Microb. Biotechnol. 2016, 9, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Perera, W.M.K.; Carter, C.G.; Houlihan, D.F. Apparent absorption efficiencies of amino acids in rainbow trout, Oncorhynchus mykiss (Walbaum), fed diets containing bacterial single-cell protein. Aquac. Nutr. 1995, 1, 95–103. [Google Scholar] [CrossRef]
- Aas, T.S.; Hatlen, B.; Grisdale-Helland, B.; Terjesen, B.F.; Bakke-McKellep, A.M.; Helland, S.J. Effects of diets containing a bacterial protein meal on growth and feed utilisation in rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 357–368. [Google Scholar] [CrossRef]
- Kiessling, A.; Askbrandt, S. Nutritive value of two bacterial strains of single-cell protein for rainbow trout (Oncorhynchus mykiss). Aquaculture 1993, 109, 119–130. [Google Scholar] [CrossRef]
- Berge, G.M.; Baeverfjord, G.; Skrede, A.; Storebakken, T. Bacterial protein grown on natural gas as protein source in diets for Atlantic salmon, Salmo salar, in saltwater. Aquaculture 2005, 244, 233–240. [Google Scholar] [CrossRef]
- Teimouri, M.; Amirkolaie, A.K.; Yeganeh, S. The effects of Spirulina platensis meal as a feed supplement on growth performance and pigmentation of rainbow trout (Oncorhynchus mykiss). Aquaculture 2013, 396, 14–19. [Google Scholar] [CrossRef]
- Hanel, R.; Broekman, D.; De Graaf, S.; Schnack, D. Partial Replacement of Fishmeal by Lyophylized Powder of the Microalgae Spirulina platensis in Pacific White Shrimp Diets. Open Mar. Biol. J. 2007, 1, 1–5. [Google Scholar] [CrossRef]
- Silva-Neto, J.F.; Nunes, A.J.P.; Sabry-Neto, H.; Sá, M.V.C. Spirulina meal has acted as a strong feeding attractant for Litopenaeus vannamei at a very low dietary inclusion level. Aquac. Res. 2012, 43, 430–437. [Google Scholar] [CrossRef]
- Macias-Sancho, J.; Poersch, L.H.; Bauer, W.; Romano, L.A.; Wasielesky, W.; Tesser, M.B. Fishmeal substitution with Arthrospira (Spirulina platensis) in a practical diet for Litopenaeus vannamei: Effects on growth and immunological parameters. Aquaculture 2014, 426, 120–125. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Yun, H.; Won, S.; Kim, S.; Farris, N.W.; Bai, S.C. Evaluation of a single-cell protein as a dietary fish meal substitute for whiteleg shrimp Litopenaeus vannamei. Fish. Sci. 2018, 85, 147–155. [Google Scholar] [CrossRef]
- Imelda, J.; Paulraj, R. Efficacy of bacterial fermented oilcake mix as fishmeal substitute in the diet of tiger shrimp, Penaeus monodon (Fabricius) post larvae. Indian J. Fish. 2007, 54, 379–387. [Google Scholar]
- Hamidoghli, A.; Won, S.; Farris, N.W.; Bae, J.; Choi, W.; Yun, H.; Bai, S.C. Solid state fermented plant protein sources as fish meal replacers in whiteleg shrimp Litopaeneus vannamei. Anim. Feed Sci. Technol. 2020, 264, 114474. [Google Scholar] [CrossRef]
- Velasquez, S.F.; Chan, M.A.; Abisado, R.G.; Traifalgar, R.F.M.; Tayamen, M.M.; Maliwat, G.C.F.; Ragaza, J.A. Dietary Spirulina (Arthrospira platensis) replacement enhances performance of juvenile Nile tilapia (Oreochromis niloticus). J. Appl. Phycol. 2016, 28, 1023–1030. [Google Scholar] [CrossRef]
- Aas, T.S.; Hatlen, B.; Grisdale-Helland, B.; Terjesen, B.F.; Penn, M.; Bakke-McKellep, A.M.; Helland, S.J. Feed intake, growth and nutrient utilization in Atlantic halibut (Hippoglossus hippoglossus) fed diets containing a bacterial protein meal. Aquac. Res. 2007, 38, 351–360. [Google Scholar] [CrossRef]
- Shapawi, R.; Ting, T.E.; Al-Azad, S. Inclusion of Purple Non-sulfur Bacterial Biomass in Formulated Feed to Promote Growth, Feed Conversion Ratio and Survival of Asian Seabass Lates calcarifer Juveniles. J. Fish. Aquat. Sci. 2012, 7, 475–480. [Google Scholar] [CrossRef][Green Version]
- Deboutteville, J.D.; Batstone, D.J.; Kawasaki, M.; Stegman, S.; Salini, M.; Tabrett, S.; Smullen, R.; Barnes, A.; Hülsen, T. Mixed culture purple phototrophic bacteria is an effective fishmeal replacement in aquaculture. Water Res. X 2019, 4, 100031. [Google Scholar] [CrossRef]
- Bauer, W.; Prentice-Hernández, C.; Tesser, M.B.; Wasielesky, W., Jr.; Poersch, L.H. Substitution of fishmeal with microbial floc meal and soy protein concentrate in diets for the pacific white shrimp Litopenaeus vannamei. Aquaculture 2012, 342, 112–116. [Google Scholar] [CrossRef]
- Arnold, S.J.; Smullen, R.; Briggs, M.; West, M.; Glencross, B. The combined effect of feed frequency and ration size of diets with and without microbial biomass on the growth and feed conversion of juvenile Penaeus monodon. Aquac. Nutr. 2015, 22, 1340–1347. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Forster, I.; Conquest, L.; Dominy, W. Enhanced growth effects on shrimp (Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to a formulated diet. Aquac. Nutr. 2008, 14, 533–543. [Google Scholar] [CrossRef]
- Kuhn, D.D.; Lawrence, A.L.; Boardman, G.D.; Patnaik, S.; Marsh, L.; Flick, G.J. Evaluation of two types of bioflocs derived from biological treatment of fish effluent as feed ingredients for Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2010, 303, 28–33. [Google Scholar] [CrossRef]
- Azim, M.; Little, D. The biofloc technology (BFT) in indoor tanks: Water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus). Aquaculture 2008, 283, 29–35. [Google Scholar] [CrossRef]
- Azim, M.; Little, D.; Bron, J.E. Microbial protein production in activated suspension tanks manipulating C:N ratio in feed and the implications for fish culture. Bioresour. Technol. 2008, 99, 3590–3599. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.J.; Blyth, D.; Fatan, N.A.; Suri, S.K. Microbial biomass (Novacq™) stimulates feeding and improves the growth performance on extruded low to zero-fishmeal diets in tilapia (GIFT strain). Aquaculture 2019, 501, 319–324. [Google Scholar] [CrossRef]
- Suphantharika, M. Preparation of spent brewer’s yeast β-glucans with a potential application as an immunostimulant for black tiger shrimp, Penaeus monodon. Bioresour. Technol. 2003, 88, 55–60. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Øverland, M.; Mydland, L.T.; Skrede, A.; Landsverk, T. Bacteria Grown on Natural Gas Prevent Soybean Meal-Induced Enteritis in Atlantic Salmon. J. Nutr. 2010, 141, 124–130. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Landsverk, T.; Mydland, L.T.; Skrede, A.; Øverland, M. Cell wall fractions from Methylococcus capsulatus prevent soybean meal-induced enteritis in Atlantic salmon (Salmo salar). Aquaculture 2013, 402, 13–18. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2016, 48, 2438–2452. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Guardiola, F.; Rojas, M.; Ascencio, F.; Esteban, M. Ángeles Dietary administration of microalgae Navicula sp. affects immune status and gene expression of gilthead seabream (Sparus aurata). Fish Shellfish. Immunol. 2013, 35, 883–889. [Google Scholar] [CrossRef]
- Burgents, J.E.; Burnett, K.G.; Burnett, L.E. Disease resistance of Pacific white shrimp, Litopenaeus vannamei, following the dietary administration of a yeast culture food supplement. Aquaculture 2004, 231, 1–8. [Google Scholar] [CrossRef]
- Sahlmann, C.; Djordjevic, B.; Lagos, L.; Mydland, L.T.; Morales-Lange, B.; Hansen, J.Ø.; Ånestad, R.; Mercado, L.; Bjelanovic, M.; Press, C.M.; et al. Yeast as a protein source during smoltification of Atlantic salmon (Salmo salar L.), enhances performance and modulates health. Aquaculture 2019, 513. [Google Scholar] [CrossRef]
- Vasanth, G.; Viswanath, K.; Kulkarni, A.; Dahle, D.; Lokesh, J.; Kitani, Y. A Microbial Feed Additive Abates Intestinal Inflammation in Atlantic Salmon. Front. Immunol. 2015, 6, 409. [Google Scholar] [CrossRef]
- Grassi, T.L.M.; Oliveira, D.L.; Paiva, N.M.; Diniz, J.C.P.; Bosco, A.M.; Pereira, A.A.F.; Menezes, A.R.P.; Valadares, T.C.; Pastor, R.C.P.; Ciarlini, P.C.; et al. Microbial biomass as an antioxidant for tilapia feed. Aquac. Res. 2018, 49, 2881–2890. [Google Scholar] [CrossRef]
- Zhao, P.; Huang, J.; Wang, X.H.; Song, X.L.; Yang, C.H.; Zhang, X.G.; Wang, G.C. The application of bioflocs technology in high-intensive, zero exchange farming systems of Marsupenaeus japonicus. Aquaculture 2012, 354, 97–106. [Google Scholar] [CrossRef]
- Samuelsen, T.A.; Oterhals, Å.; Kousoulaki, K. High lipid microalgae (Schizochytrium sp.) inclusion as a sustainable source of n-3 long-chain PUFA in fish feed—Effects on the extrusion process and physical pellet quality. Anim. Feed Sci. Technol. 2018, 236, 14–28. [Google Scholar] [CrossRef]
- Øverland, M.; Romarheim, O.H.; Ahlstrøm, Ø.; Storebakken, T.; Skrede, A. Technical quality of dog food and salmon feed containing different bacterial protein sources and processed by different extrusion conditions. Anim. Feed Sci. Technol. 2007, 134, 124–139. [Google Scholar] [CrossRef]
- Lu, J.; Takeuchi, T.; Ogawa, H. Flesh quality of tilapia Oreochromis niloticus fed solely on raw Spirulina. Fish. Sci. 2003, 69, 529–534. [Google Scholar] [CrossRef]
- Lopes da Silva, T.; Moniz, P.; Silva, C.; Reis, A. The Dark Side of Microalgae Biotechnology: A Heterotrophic Biorefinery Platform Directed to ω-3 Rich Lipid Production. Microorganisms 2019, 7, 670. [Google Scholar] [CrossRef]
- Sillman, J.; Nygren, L.; Kahiluoto, H.; Ruuskanen, V.; Tamminen, A.; Bajamundi, C.; Nappa, M.; Wuokko, M.; Lindh, T.; Vainikka, P.; et al. Bacterial protein for food and feed generated via renewable energy and direct air capture of CO2: Can it reduce land and water use? Glob. Food Secur. 2019, 22, 25–32. [Google Scholar] [CrossRef]
- Ketzer, F.; Skarka, J.; Rösch, C. Critical Review of Microalgae LCA Studies for Bioenergy Production. BioEnergy Res. 2017, 11, 95–105. [Google Scholar] [CrossRef]
- Guedes, A.C.; Pinto, I.S.; Malcata, F. Application of Microalgae Protein to Aquafeed. In Handbook of Marine Microalgae; Academic Press: London, UK, 2015; pp. 93–125. [Google Scholar]
- Chauton, M.S.; Reitan, K.I.; Norsker, N.H.; Tveteras, R.; Kleivdal, H.T. A techno-economic analysis of industrial production of marine microalgae as a source of EPA and DHA-rich raw material for aquafeed: Research challenges and possibilities. Aquaculture 2015, 436, 95–103. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | (Super)Phylum | Class | Genus | Species | Application | Data Source |
|---|---|---|---|---|---|---|
| Microalgae | Chlorophyta | Trebouxiophyceae | Chlorella | vulgaris | SCP | [50] |
| Chlorophyceae | Haematococcus | pluvialis | Bioactive | [51] | ||
| Heterokonta | Labyrinthulomycetes | Schizochytrium | sp. | SCO, SCP | [52,53] | |
| Eustigmatophyceae | Nannochloropsis | gaditana | SCP | [15] | ||
| Coscinodiscophyceae | Chaetoceros | muelleri | SCO | [54] | ||
| Coscinodiscophyceae | Skeletonema | costatum | SCO | [55] | ||
| Bacillariophyceae | Navicula | gregaria | SCO | [55] | ||
| Haptophyta | Pavlovophyceae | Pavlova | lutheri | SCO | [32,56] | |
| Prymnesiophyceae | Isochrysis | galbana | SCO, SCP | [32,57] | ||
| Dinoflagellata | Dinophyceae | Crypthecodinium | cohnii | SCO | [20] | |
| Fungal | Ascomycota | Saccharomycetes | Saccharomyces | cerevisiae | SCP | [41,58] |
| Saccharomycetes | Wikerhamomyces | anomalus | SCP | [41,59] | ||
| Saccharomycetes | Candida | utilis | SCP | [58] | ||
| Saccharomycetes | Kluyveromyces | marxianus | SCP | [58] | ||
| Mucoromycota | Mortierellales | Mortierella | alpina | SCO | [60] | |
| Bacteria | Cyanobacteria | Cyanophyceae | Spirulina | maxima | SCP | [61] |
| Proteobacteria | Gammaproteobacteria | Methylococcus | capsulatus | SCP | [62] | |
| Betaproteobacteria | Methylophilus | methylotrophus | SCP | [63] | ||
| Alphaproteobacteria | Methylobacterium | extorquens | SCP | [64] |
| Group | Microalgal | Microalgal | Fungal | Fungal | Fungal | Bacterial | Bacterial | Bacterial | Bacterial |
|---|---|---|---|---|---|---|---|---|---|
| Species | Chlorella | Nannochloropsis | Saccharomyces | Wikerhamomyces | Candida | Spirulina | Methylophilus | Methylococcus | Methylobacterium |
| Material | Biomass | Biomass | Biomass | Biomass | Biomass | Biomass | Biomass | Biomass | Biomass |
| Data Source | [36] | [35] | [41] | [41] | [80] | [36] | [5] | [62] | [64] |
| Dry matter | 95.0 | 94.9 | 94.0 | 93.0 | 96.4 | 82.2 | 100 | 98.6 | 96.3 |
| Moisture | 5.0 | 5.1 | 6.0 | 7.0 | 3.6 | 17.8 | 0.0 | 1.4 | 3.7 |
| Protein | 54.5 | 47.7 | 46.6 | 42.2 | 33.3 | 61.3 | 81.3 | 68.1 | 50.9 |
| Lipid | 9.4 | 8.4 | 1.0 | 0.9 | 2.1 | 5.5 | 7.2 | 10.4 | 0.8 |
| Ash | 5.3 | 7.5 | 6.3 | 7.0 | 9.8 | 6.9 | 9.1 | 8.0 | 4.1 |
| Carbohydrates | 25.8 | 31.3 | 40.1 | 42.9 | 51.2 | 8.5 | 2.4 | 12.1 | 40.5 |
| Energy (kJ/g) | 20.9 | 19.9 | 18.3 | 17.7 | 17.5 | 18.1 | 22.4 | 22.2 | 19.3 |
| Sum Amino Acids | - | 37.6 | 42.4 | 36.0 | 31.7 | - | 68.2 | 55.9 | 42.9 |
| Alanine | - | 2.5 | 2.3 | 2.0 | 2.1 | - | 5.8 | 4.4 | 3.8 |
| Arginine | 2.9 | 2.3 | 2.0 | 1.6 | 1.7 | 4.1 | 3.7 | 3.8 | 3.3 |
| Aspartic acid | - | 3.3 | 3.5 | 3.1 | 3.4 | - | 7.2 | 5.5 | 4.4 |
| Cysteine | - | 0.3 | 0.0 | 0.0 | 0.3 | - | 0.6 | 0.5 | 0.3 |
| Glutamic acid | - | 4.6 | 13.2 | 11.4 | 4.3 | - | 8.6 | 6.5 | 6.3 |
| Glycine | - | 1.8 | 1.2 | 0.9 | 1.6 | - | 4.6 | 3.1 | 2.5 |
| Histidine | 1.0 | 0.7 | 0.9 | 0.7 | 0.6 | 1.0 | 1.5 | 1.3 | 1.1 |
| Isoleucine | 1.5 | 1.7 | 1.9 | 1.6 | 1.7 | 2.6 | 3.5 | 2.7 | 1.7 |
| Leucine | 4.2 | 3.2 | 3.1 | 2.6 | 2.6 | 4.7 | 5.7 | 5.0 | 3.3 |
| Lysine | 4.6 | 2.3 | 3.3 | 2.7 | 2.3 | 3.1 | 4.9 | 3.5 | 2.5 |
| Methionine | 1.0 | 0.9 | 0.3 | 0.2 | 0.4 | 1.4 | 2.0 | 1.5 | 0.9 |
| Phenylalanine | 2.3 | 1.9 | 2.2 | 1.8 | 1.5 | 2.5 | 3.3 | 3.0 | 2.0 |
| Proline | - | 4.7 | 1.8 | 1.5 | 1.4 | - | 2.4 | 2.5 | 1.9 |
| Serine | - | 1.6 | 1.3 | 1.5 | 1.9 | - | 2.7 | 2.4 | 1.9 |
| Taurine | - | 0.0 | 0.0 | 0.2 | - | - | - | - | - |
| Threonine | 2.0 | 1.7 | 1.6 | 1.5 | 2.1 | 2.7 | 3.7 | 2.8 | 2.0 |
| Tryptophan | 1.5 | 0.8 | 0.0 | 0.0 | 0.5 | 1.2 | 0.7 | 1.3 | - |
| Tyrosine | - | 1.1 | 1.6 | 1.4 | 1.1 | - | 2.8 | 2.2 | 1.4 |
| Valine | 2.4 | 2.2 | 2.5 | 2.0 | 2.1 | 0.3 | 4.6 | 3.7 | 2.8 |
| Group | Microalgal | Microalgal | Microalgal | Microalgal | Microalgal | Microalgal | Microalgal | Fungal |
|---|---|---|---|---|---|---|---|---|
| Species | Schizochytrium | Schizochytrium | Crypthecodinium | Chlorella | Isochrysis | Pavlova | Nannochloropsis | Yarrowia |
| Material | Biomass | Oil | Biomass | Whole-cell | Biomass | Biomass | Biomass | Biomass |
| Data Source | [86] | [34] | [33] | [87] | [88] | [32] | [32] | [84] |
| Dry matter | 98.4 | 98.9 | 100.0 | 96.4 | 100.0 | - | - | 95.3 |
| Moisture | 1.6 | 1.1 | - | 3.6 | 0.0 | - | - | 4.7 |
| Protein | 13.2 | 0.0 | - | 30.4 | 29.5 | - | - | 29.8 |
| Lipid | 61.4 | 98.6 | 50.0 | 26.0 | 23.4 | - | - | 20.3 |
| Ash | 4.4 | 0.3 | - | 3.3 | 13.2 | - | - | 2.1 |
| Carbohydrate | 19.4 | 0.0 | - | 36.7 | 33.9 | - | - | 43.1 |
| Energy | 28.8 | 39.1 | - | 23.5 | 21.8 | - | - | 22.3 |
| Fatty Acids | ||||||||
| C14:0 | 4.1 | 12.0 | 16.0 | 0.7 | 17.3 | 10.0 | 4.8 | 0.3 |
| C15:0 | 2.0 | 1.5 | 0.1 | 0.1 | 0.0 | 0.2 | 0.5 | - |
| C16:0 | 52.8 | 26.3 | 25.0 | 17.1 | 12.0 | 20.8 | 20.3 | 10.7 |
| C18:0 | 1.5 | 0.9 | 0.0 | 1.9 | 1.1 | 0.2 | 0.3 | 6.6 |
| C20:0 | 0.2 | 0.0 | 0.0 | 0.2 | - | - | - | 0.7 |
| C22:0 | 0.1 | 0.0 | 0.0 | 0.1 | - | - | - | 1.0 |
| Total Saturates | 61.4 | 41.9 | 41.1 | 21.1 | 30.3 | 31.2 | 26.2 | 19.3 |
| C16:1n-7 | 0.1 | 4.5 | 0.4 | 1.6 | 3.1 | 19.9 | 21.2 | 1.5 |
| C18:1n-9 | 0.1 | 0.7 | 16.0 | 30.6 | 6.9 | 0.5 | 4.1 | 8.8 |
| C18:1n-7 | - | 2.9 | - | 0.5 | 1.1 | 3.3 | 0.5 | |
| C20:1n-11 | 0.1 | 0 | - | - | - | - | - | 0.2 |
| C22:1n-11 | 0.2 | 0 | - | - | - | - | - | 0.9 |
| Total Monoenes | 0.8 | 8.1 | 16.4 | 42.1 | 11.3 | 23.8 | 27.0 | 11.4 |
| C18:2n-6 | 0.1 | 0.3 | 0.5 | 19.7 | 4.0 | 0.4 | 1.8 | 22.9 |
| C18:3n-6 | 0.0 | 0.0 | 0.0 | - | 1.0 | 0.2 | 1.0 | |
| C20:2n-6 | 0.1 | 0.0 | 0.0 | - | - | - | - | 2.7 |
| C20:3n-6 | 0.1 | 0.0 | 0.0 | - | - | - | - | 3.3 |
| C20:4n-6 | 0.3 | 0.0 | 0.0 | - | 0.0 | 0.6 | 7.4 | 0.5 |
| C22:4n-6 | 0.0 | 0.0 | 0.0 | 0.0 | - | - | - | |
| C22:5n-6 | 6.9 | 7.5 | 0.0 | 0.1 | 2.1 | 0.9 | - | |
| Total n-6 PUFA * | 7.4 | 7.8 | 0.5 | 25.6 | 7.1 | 2.1 | 10.2 | 29.4 |
| C18:3n-3 | 0.0 | 0.0 | 0.4 | 13.6 | 5.7 | 1.1 | - | 2.3 |
| C18:4n-3 | 0.0 | 0.2 | 0.0 | 0.1 | 19.0 | 6.6 | - | 0.3 |
| C20:4n-3 | 0.1 | 0.4 | 0.0 | 0.1 | - | - | - | 3.5 |
| C20:5n-3 | 0.3 | 0.8 | 0.1 | - | 0.9 | 22.9 | 34.7 | 30.2 |
| C22:5n-3 | 0.1 | 0.0 | 0.0 | - | 0.0 | 0.1 | - | 0.9 |
| C22:6n-3 | 28.5 | 40.9 | 39.0 | - | 9.9 | 7.9 | - | 0.0 |
| Total n-3 PUFA | 29.0 | 42.2 | 39.5 | 14.5 | 35.5 | 38.6 | 34.7 | 37.2 |
| Group | SCP Species | Processing | Test Species | Data | Digestibility (Protein) | Inclusion (g/kg) | Data Source |
|---|---|---|---|---|---|---|---|
| Microalgae | Nannochloropsis | Cold-pelleting | Salmo salar | Ingredient | 73% | 300 | [99] |
| Desmodesmus | Cold-pelleting | Salmo salar | Ingredient | 54% | 300 | [99] | |
| Nannochloropsis | Extruded | Salmo salar | Ingredient | 72% | 300 | [99] | |
| Desmodesmus | Extruded | Salmo salar | Ingredient | 67% | 300 | [99] | |
| Spirulina | Dried (undefined) | Oreochromis niloticus | Ingredient | 86% | 300 | [36] | |
| Chlorella | Dried (undefined) | Oreochromis niloticus | Ingredient | 80% | 300 | [36] | |
| Schizocytrium | Dried (undefined) | Oreochromis niloticus | Ingredient | 82% | 300 | [36] | |
| Nannochloropsis | Drum dried | Oreochromis niloticus | Ingredient | 75% | 300 | [102] | |
| Arthrospira (Spirulina) | Dried (undefined) | Oreochromis niloticus | Ingredient | 83% | 300 | [102] | |
| Chlorella | Dried (undefined) | Oreochromis niloticus | Ingredient | 81% | 300 | [102] | |
| Scenedesmus | Dried (undefined) | Oreochromis niloticus | Ingredient | 67% | 300 | [102] | |
| Nannochloropsis | Drum dried | Oreochromis niloticus | Ingredient | 62% | 300 | [15] | |
| Pasteurised | Oreochromis niloticus | Ingredient | 61% | 300 | [15] | ||
| Freeze-dried | Oreochromis niloticus | Ingredient | 61% | 300 | [15] | ||
| Frozen/Thawed | Oreochromis niloticus | Ingredient | 66% | 300 | [15] | ||
| Bead milled | Oreochromis niloticus | Ingredient | 78% | 300 | [15] | ||
| Commercial | Oreochromis niloticus | Ingredient | 73% | 300 | [15] | ||
| Nannochloropsis | Dried (undefined) | Oncorhynchus mykiss | Ingredient | 69% | 300 | [101] | |
| Isochrysis | Dried (undefined) | Oncorhynchus mykiss | Ingredient | 87% | 300 | [101] | |
| Fishmeal | - | Clarius gariepinus | Ingredient | 98% | 300 | [100] | |
| Spirulina | Spray-drying | Clarius gariepinus | Ingredient | 99% | 300 | [100] | |
| Chlorella | Spray-drying | Clarius gariepinus | Ingredient | 99% | 300 | [100] | |
| Fungal | Saccharomyces | Intact | Oncorhynchus mykiss | Ingredient | 63% | n.s. | [104] |
| Disrupted | Oncorhynchus mykiss | Ingredient | 85% | n.s. | [104] | ||
| Extract | Oncorhynchus mykiss | Ingredient | 81% | n.s. | [104] | ||
| Nucleoprotein complex | Oncorhynchus mykiss | Ingredient | 83% | n.s. | [104] | ||
| Protein isolate | Oncorhynchus mykiss | Ingredient | 87% | n.s. | [104] | ||
| Reference | extruded | Salmo salar | Diet | 88% | - | [58] | |
| Saccharomyces | extruded | Salmo salar | Diet | 73% | 345 | [58] | |
| Kluyveromyces | extruded | Salmo salar | Diet | 86% | 302 | [58] | |
| Candida | extruded | Salmo salar | Diet | 88% | 283 | [58] | |
| Rhizopus | dried | Salvelinus alpinus | Ingredient | 94% | 300 | [105] | |
| Saccharomyces | dried | Salvelinus alpinus | Ingredient | 86% | 300 | [105] | |
| Saccharomyces-Extracted | autolysis + centrifugation | Salvelinus alpinus | Ingredient | 98% | 300 | [105] | |
| Candida | Spray dried | Salmo salar | Ingredient | 41% | 300 | [80] | |
| Reference | extruded | Oncorhynchus mykiss | Diet | 91% | - | [59] | |
| Saccharomyces | extruded | Oncorhynchus mykiss | Diet | 87% | 321 | [59] | |
| Wickerhamomyces | extruded | Oncorhynchus mykiss | Diet | 87% | 355 | [59] | |
| Bacterial | Methylophilus | dried | Oncorhynchus mykiss | Diet | 78% | 0 | [5] |
| dried | Oncorhynchus mykiss | Diet | 79% | 70 | [5] | ||
| dried | Oncorhynchus mykiss | Diet | 80% | 140 | [5] | ||
| dried | Oncorhynchus mykiss | Diet | 81% | 210 | [5] | ||
| dried | Oncorhynchus mykiss | Diet | 84% | 280 | [5] | ||
| dried | Oncorhynchus mykiss | Diet | 82% | 350 | [5] | ||
| Methylococcus + Alcaligenes + Bacilus | Spray-dried | Salmo salar | Ingredient | 82% | 193/370 | [46] | |
| Methylococcus + Alcaligenes + Bacilus | Reference | Salmo salar | Diet | 90% | - | [106] | |
| Spray-dried | Salmo salar | Diet | 88% | 200 | [106] | ||
| Methylococcus + Alcaligenes + Bacilus | Spray-dried | Salmo salar | Diet | 90% | 0 | [62] | |
| Spray-dried | Salmo salar | Diet | 88% | 50 | [62] | ||
| Spray-dried | Salmo salar | Diet | 88% | 99 | [62] | ||
| Spray-dried | Salmo salar | Diet | 87% | 193 | [62] | ||
| Spray-dried | Salmo salar | Diet | 84% | 370 | [62] | ||
| Methylococcus + Alcaligenes + Bacilus | Spray-dried | Salmo salar | Diet | 88% | 0 | [107] | |
| Spray-dried | Salmo salar | Diet | 87% | 180 | [107] | ||
| Spray-dried | Salmo salar | Diet | 85% | 360 | [107] | ||
| Methylococcus + Ralstonia + Aneurinibacillus + Brevibacillus | Reference | Oncorhynchus mykiss | Diet | 89% | 0 | [108] | |
| Spray-dried | Oncorhynchus mykiss | Diet | 89% | 75 | [108] | ||
| Spray-dried | Oncorhynchus mykiss | Diet | 88% | 147 | [108] | ||
| Autolysate | Oncorhynchus mykiss | Diet | 89% | 73 | [108] | ||
| Autolysate | Oncorhynchus mykiss | Diet | 88% | 147 | [108] | ||
| Biofloc | Molasses stimulated catfish water | Oven dried | Litopenaeus vannamei | Ingredient | 77% | 300 | [109] |
| Activated sludge from shrimp pond | Oven dried | Litopenaeus vannamei | Ingredient | 26% | 300 | [110] | |
| RAS effluent --> Sequence Batch Reactor | Untreated | Oreochromis niloticus | Diet | 68% | n.s. | [111] |
| Group | SCO Species | Material | Processing | Test Species | Data | Digestibility | Inclusion (g/kg) | Data Source | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Sats | Monos | n-6 | n-3 | ||||||||
| Microalgae | Schizochytrium | Biomass | Dried | Salmo salar 36 g | Diet | 94% | 98% | 100% | 100% | 100% | 100 | [112] |
| Canola | Oil | - | Salmo salar 36 g | Diet | 96% | 100% | 100% | 99% | 100% | 169 | [112] | |
| Fish (Jack mackerel) | Oil | - | Salmo salar 36 g | Diet | 96% | 99% | 100% | 100% | 100% | 91 | [112] | |
| Microalgae | Schizochytrium (S) | Oil | Undefined | Salmo salar 40 g | Diet | - | 71% | 75% | 93% | 97% | 130 | [113] |
| Palm | Oil | Undefined | Salmo salar 40 g | Diet | - | 78% | 92% | 95% | 97% | 130 | [113] | |
| Palm + S | Oil | Undefined | Salmo salar 40 g | Diet | - | 71% | 91% | 95% | 96% | 104 + 26 | [113] | |
| Fish (Jack mackerel) | Oil | Undefined | Salmo salar 40 g | Diet | - | 76% | 87% | 92% | 96% | 130 | [113] | |
| Microalgae | Schizochytrium | Oil | Enzymatic | Salmo salar 32 g | Diet | 90% | 76% | 92% | 96% | 98% | 90 | [34] |
| Microalgae | Schizochytrium + YE | Biomass | Reference | Salmo salar 213 g | Diet | 96% | 96% | 99% | 99% | 99% | 0 | [114] |
| +Fungal | Biomass | Dried | Salmo salar 213 g | Diet | 96% | 95% | 99% | 99% | 99% | 10 | [114] | |
| Biomass | Dried | Salmo salar 213 g | Diet | 94% | 86% | 99% | 99% | 99% | 60 | [114] | ||
| Biomass | Dried | Salmo salar 213 g | Diet | 88% | 67% | 99% | 99% | 99% | 150 | [114] | ||
| Microalgae | Schizochytrium | Biomass | Spray-dried | Salmo salar 400 g | Diet | 94% | 93% | 99% | 99% | 99% | 0 | [86] |
| Biomass | Spray-dried | Salmo salar 400 g | Diet | 93% | 87% | 99% | 99% | 99% | 25 | [86] | ||
| Biomass | Spray-dried | Salmo salar 400 g | Diet | 92% | 79% | 99% | 99% | 99% | 50 | [86] | ||
| Microalgae | Schizochytrium | Biomass | Spray-dried | Salmo salar 65–270 g | Diet | 91% | 78% | 99% | 98% | 99% | 52 | [115] |
| Fishoil | - | Salmo salar 65–270 g | Diet | 95% | 96% | 99% | 98% | 99% | 0 | [115] | ||
| Biomass | Spray-dried | Salmo salar >860 g | Diet | 87% | 63% | 92% | 95% | 96% | 62 | [115] | ||
| Fishoil | - | Salmo salar >860 g | Diet | 85% | 81% | 84% | 90% | 92% | 0 | [115] | ||
| Microalgae | Crypthecodinium | Biomass | Spray-dried | Oncorhynchus mykiss | Ingred. | 98% | - | - | - | - | 300 | [116] |
| Microalgae | Spirulina | Biomass | Dried | Oreochromis niloticus | Ingred. | 95% | 76% | 76% | 84% | - | 300 | [36] |
| Chlorella | Biomass | Dried | Oreochromis niloticus | Ingred. | 94% | 75% | 70% | 77% | 39% | 300 | [36] | |
| Schizochytrium | Biomass | Dried | Oreochromis niloticus | Ingred. | 98% | 52% | 85% | 92% | 97% | 300 | [36] | |
| Microalgae | Phaeodactylum | Biomass | Reference | Salmo salar | Diet | 96% | 97% | 99% | 96% | 99% | 0 | [117] |
| Biomass | Dried | Salmo salar | Diet | 96% | 97% | 98% | 96% | 99% | 30 | [117] | ||
| Biomass | Dried | Salmo salar | Diet | 95% | 96% | 98% | 96% | 99% | 60 | [117] | ||
| Biomass | Dried | Salmo salar | Diet | 94% | 95% | 98% | 97% | 99% | 120 | [117] | ||
| Microalgae | Nannochloropsis | Biomass | Dried | Oreochromis niloticus | Ingred. | 50% | - | - | - | - | 300 | [15] |
| Biomass | Pasteruised | Oreochromis niloticus | Ingred. | 56% | - | - | - | - | 300 | [15] | ||
| Biomass | Freeze-dried | Oreochromis niloticus | Ingred. | 58% | - | - | - | - | 300 | [15] | ||
| Biomass | Frozen/Thawed | Oreochromis niloticus | Ingred. | 53% | - | - | - | - | 300 | [15] | ||
| Biomass | Bead milled | Oreochromis niloticus | Ingred. | 82% | - | - | - | - | 300 | [15] | ||
| Biomass | Commercial | Oreochromis niloticus | Ingred. | 66% | - | - | - | - | 300 | [15] | ||
| Microalgae | Nannochloropsis | Biomass | Dried | Oncorhynchus mykiss | Ingred. | 60% | 56% | 45% | 76% | 64% | 300 | [101] |
| Isochrysis | Biomass | Dried | Oncorhynchus mykiss | Ingred. | 63% | 59% | 72% | 98% | 93% | 300 | [101] | |
| Microalgae | Chlorella | Biomass | Whole-cell dried | Salmo salar | Ingred. | 67% | 61% | 59% | 63% | 62% | 60–300 | [87] |
| Biomass | Cell-ruptured | Salmo salar | Ingred. | 85% | 62% | 88% | 93% | 93% | 60–300 | [87] | ||
| Fungal | Yarrowia | Biomass | Whole-cell dried | Salmo salar | Diet | 95% | 87% | 97% | 96% | 94% | 100 | [84] |
| Biomass | Whole-cell dried | Salmo salar | Diet | 94% | 81% | 97% | 96% | 90% | 200 | [84] | ||
| Biomass | Whole-cell dried | Salmo salar | Diet | 91% | 77% | 96% | 93% | 89% | 300 | [84] | ||
| Oil | Fishoil + Rape | Salmo salar | Diet | 96% | 90% | 97% | 96% | 96% | 73 + 126 | [84] | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glencross, B.D.; Huyben, D.; Schrama, J.W. The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review. Fishes 2020, 5, 22. https://doi.org/10.3390/fishes5030022
Glencross BD, Huyben D, Schrama JW. The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review. Fishes. 2020; 5(3):22. https://doi.org/10.3390/fishes5030022
Chicago/Turabian StyleGlencross, Brett D., David Huyben, and Johan W. Schrama. 2020. "The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review" Fishes 5, no. 3: 22. https://doi.org/10.3390/fishes5030022
APA StyleGlencross, B. D., Huyben, D., & Schrama, J. W. (2020). The Application of Single-Cell Ingredients in Aquaculture Feeds—A Review. Fishes, 5(3), 22. https://doi.org/10.3390/fishes5030022