Accumulation of Geosmin and 2-methylisoborneol in European Whitefish Coregonus Lavaretus and Rainbow Trout Oncorhynchus Mykiss in RAS



Abstract
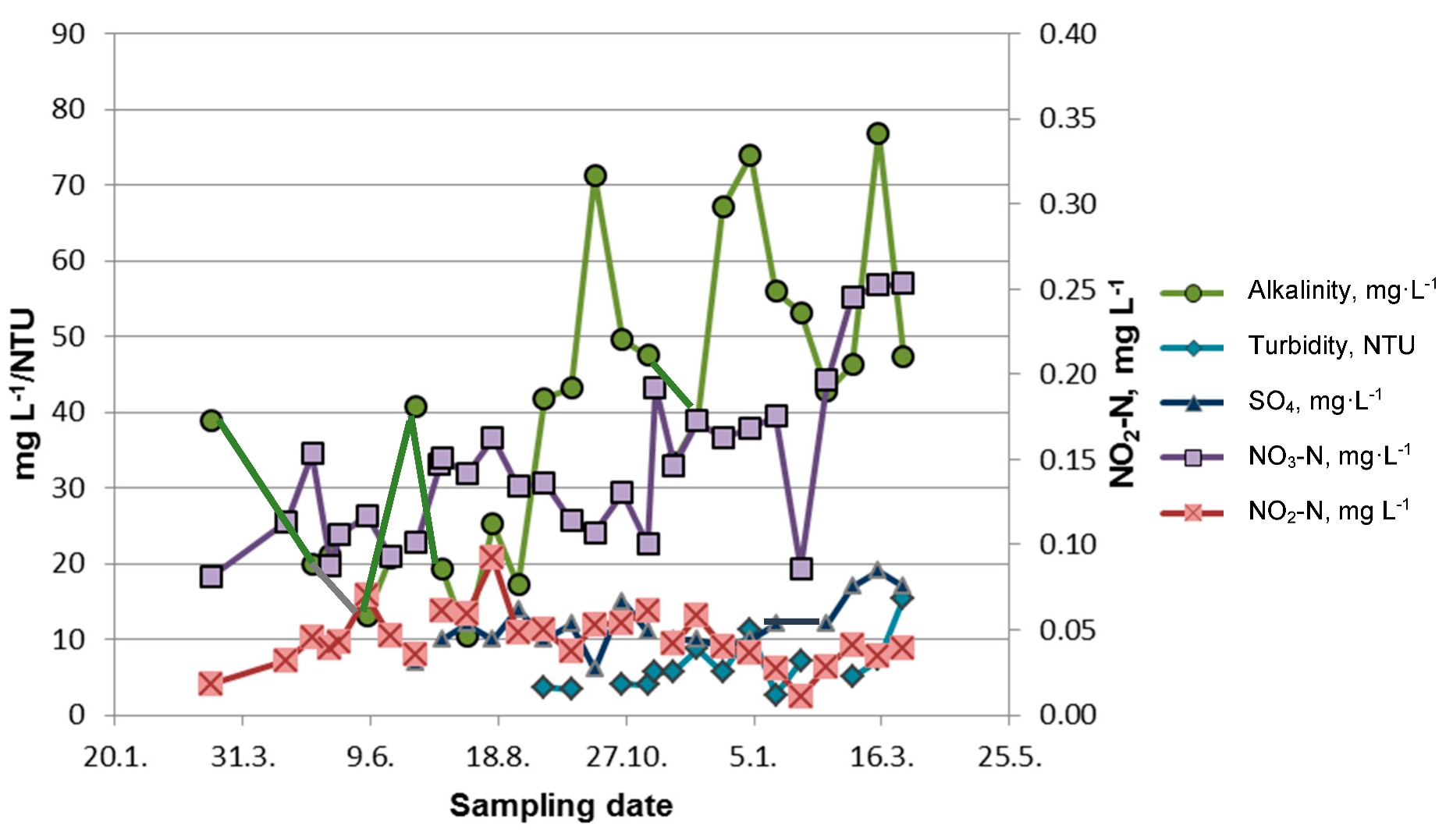
1. Introduction
2. Results
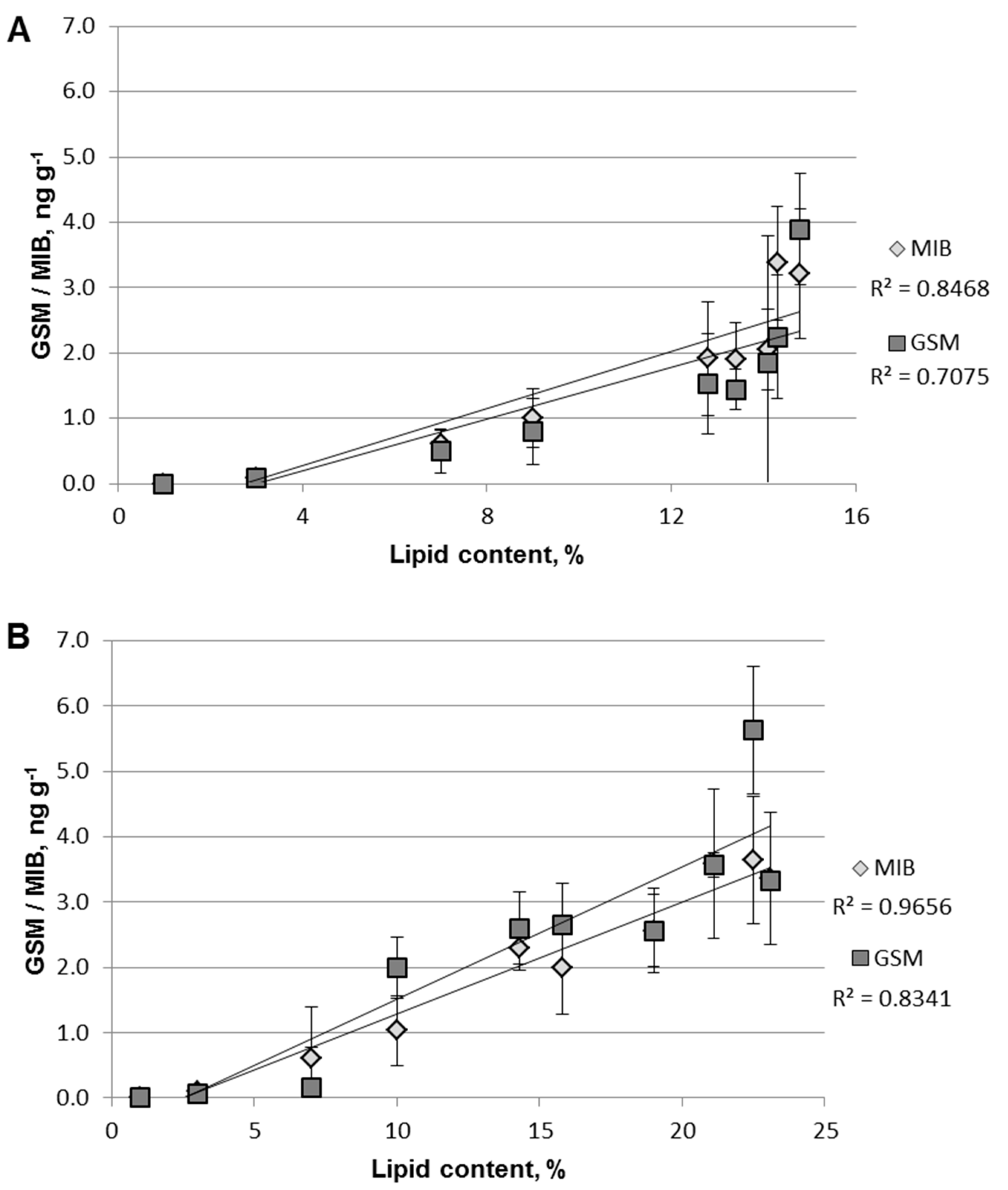
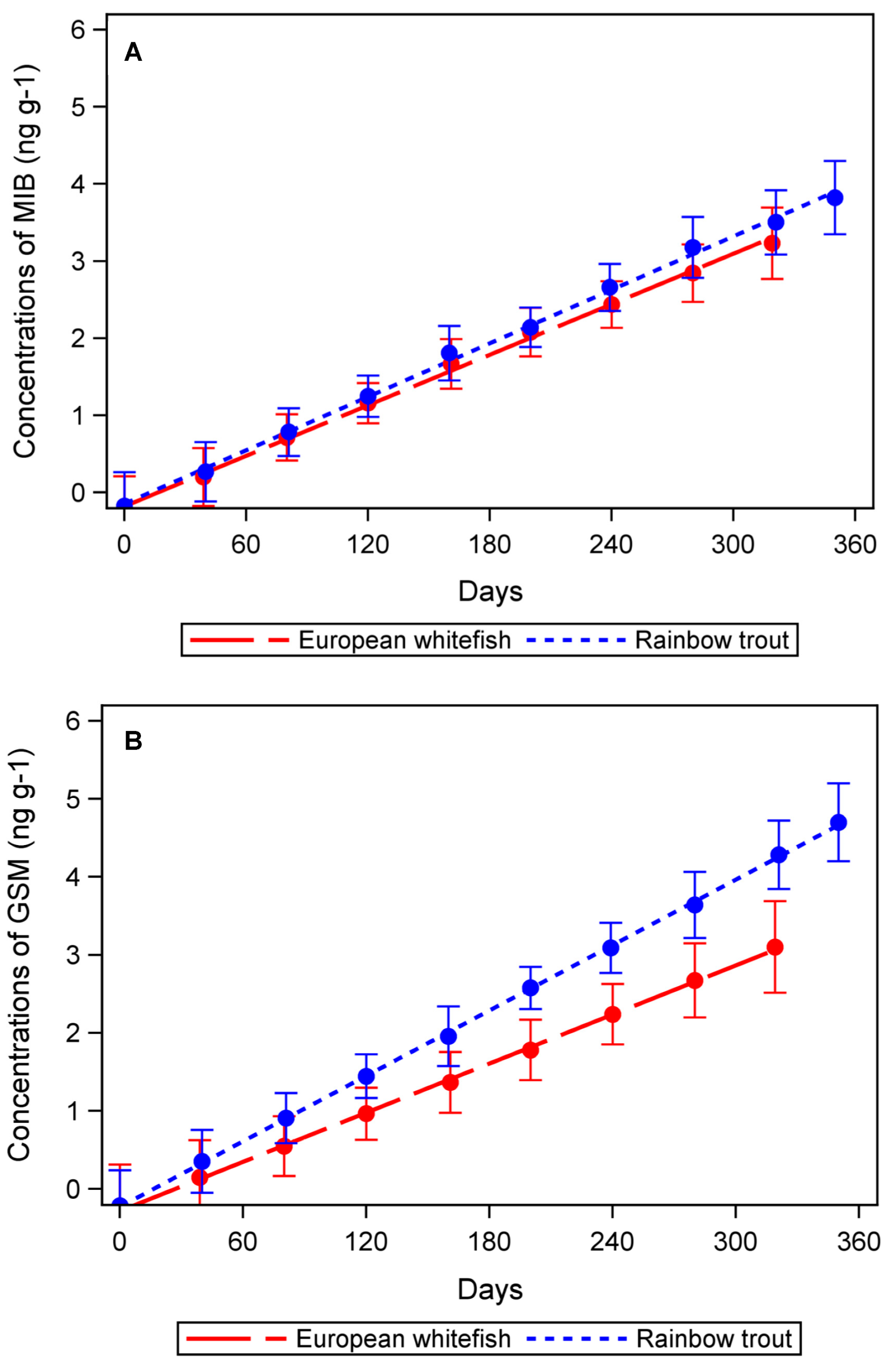
2.1. GSM and MIB in Fish Fillet
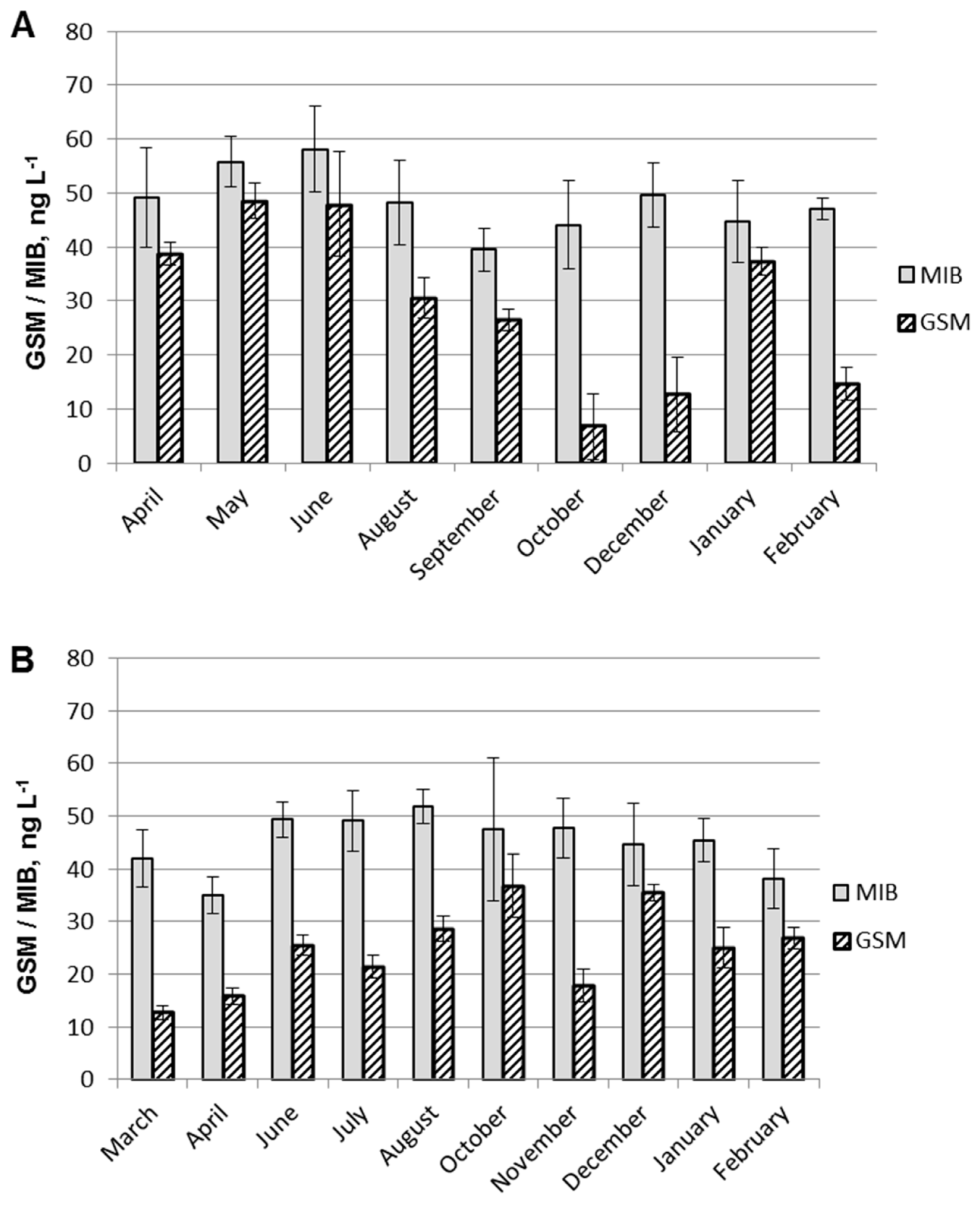
2.2. GSM and MIB in Water
2.3. Statistical Analyses of GSM and MIB Concentrations
3. Discussion
4. Materials and Methods
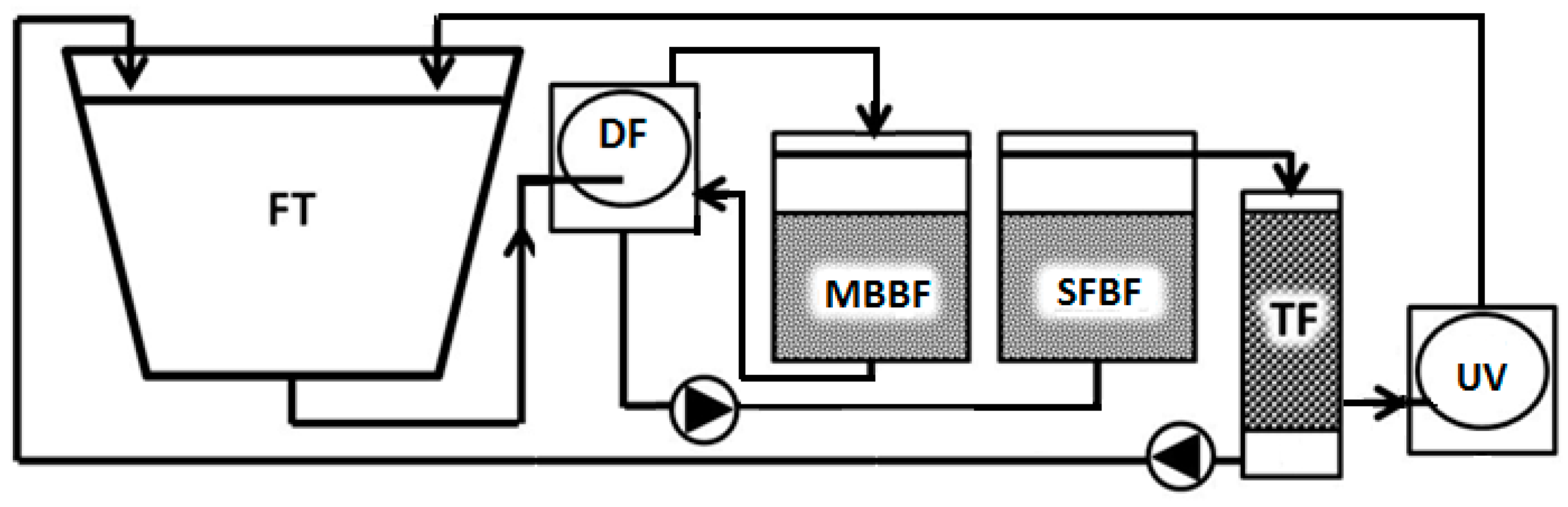
4.1. Experimental Setup
4.2. Fish and Feeding
4.3. Sample Collection for Off-flavor Analyses
4.4. Chemical Analyses
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dalsgaard, J.; Lund, I.; Thorarinsdottir, R.; Drengstig, A.; Arvonen, K.; Pedersen, P.B. Farming different species in RAS in Nordic countries: Current status and future perspectives. Aquac. Eng. 2013, 53, 2–13. [Google Scholar] [CrossRef]
- Burr, G.S.; Wolters, W.R.; Schrader, K.K.; Summerfelt, S.T. Impact of depuration of earthy-musty off-flavors on fillet quality of Atlantic salmon, Salmo salar, cultured in a recirculating aquaculture system. Aquac. Eng. 2012, 50, 28–36. [Google Scholar] [CrossRef]
- Hathurusingha, P.I.; Davey, K.R. Experimental validation of a time-dependent model for chemical taste taint accumulation as geosmin (GSM) and 2-methylisoborneol (MIB) in commercial RAS farmed barramundi (Lates calcarifer). Ecol. Model. 2016, 340, 17–27. [Google Scholar] [CrossRef]
- Lindholm-Lehto, P.C.; Vielma, J.; Pakkanen, H.; Alén, R. Depuration of geosmin- and 2-methylisoborneol-induced off-flavors in recirculating aquaculture system (RAS) farmed European whitefish Coregonus lavaretus. J. Food Sci. Technol. 2019, 56, 4585–4594. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.S. Off-flavor problems in aquaculture. Rev. Fish. Sci. 2000, 8, 45–88. [Google Scholar] [CrossRef]
- Petersen, M.A.; Md. Alam, A.; Md. Rahman, M.; Md. Ali, L.; Mahmud, S.; Schlüter, L.; Jørgensen, N.O.G. Geosmin off-flavour in pond-raised fish in southern Bangladesh and occurrence of potential off-flavour producing organisms. Aquac. Environ. Interact. 2014, 5, 107–116. [Google Scholar] [CrossRef][Green Version]
- Liu, H.; Li, H.; Wei, H.; Zhu, X.; Han, D.; Jin, J.; Yang, Y.; Xie, S. Biofloc formation improves water quality and fish yield in a freshwater pond aquaculture system. Aquaculture 2019, 506, 256–269. [Google Scholar] [CrossRef]
- Wang, R.; Dong, L.; Chao, X.J.; Bing, W.Y. Seasonal occurrence and species specificity of fishy and musty odor in Huajiang Reservoir in winter, China. Water Res. Ind. 2015, 11, 13–26. [Google Scholar] [CrossRef]
- Gerber, N.N. Geosmin from microorganisms in trans-1,10-dimethyl-trans-9-decalol. Tetrahedron Lett. 1968, 25, 2971–2974. [Google Scholar] [CrossRef]
- Gerber, N.N. A volatile metabolite of actinomycetes: 2-methylisoborneol. J. Antibiot. 1969, 22, 508–509. [Google Scholar] [CrossRef]
- Polak, E.H.; Provasi, J. Odor sensitivity to geosmin enantiomers. Chem. Senses 1992, 17, 23–26. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Bode, H.B.; Mahmud, T.; Müller, R.; Schulz, S. A novel type of geosmin biosynthesis in myxobacteria. J. Org. Chem. 2005, 70, 5174–5182. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, F.; Medger, A.; Börnick, H.; Opitz, M.; Lang, K.; Göttfert, M.; Röske, I. Identification and expression analyses of putative sesquiterpene synthase genes in Phormidium sp. and prevalence of geoA-like genes in a drinking water reservoir. Appl. Environ. Microbiol. 2007, 73, 6988–6993. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Li, L.; Zhang, T.; Zheng, L.; Dai, G.; Liu, L.; Song, L. Contribution of Streptomyces in sediment to earthy odor in the overlying water in Xionghe reservoir, China. Water Res. 2010, 44, 6085–6094. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.E.; Gobas, A.P.C.; Mackay, D. Model of organic chemical uptake and clearance by fish from food and water. Environ. Sci. Technol. 1990, 24, 1203–1213. [Google Scholar] [CrossRef]
- Nichols, J.W.; McKim, J.M.; Anderdsen, M.E.; Gargas, M.L.; Clewell, H.J.; Erickson, R.J. A physiologically based toxicokinetic model for the uptake and disposition of waterborne organic chemicals in fish. Toxicol. Appl. Pharmacol. 1990, 106, 433–447. [Google Scholar] [CrossRef]
- Arnot, J.A.; Gobas, F.A.P.C. A review of bioconcentration factor (BCF) and bioaccumulation factor (BAF) assessments for organic chemicals in aquatic organisms. Environ. Rev. 2006, 14, 257–297. [Google Scholar] [CrossRef]
- Howgate, P. Tainting of farmed fish by geosmin and 2-methyliso-borneol: A review of sensory aspects and of uptake/depuration. Aquaculture 2004, 234, 155–181. [Google Scholar] [CrossRef]
- Watson, S.B.; Monis, P.; Baker, P.; Giglio, S. Biochemistry and genetics of taste- and odor producing cyanobacteria. Harmful Algae 2016, 54, 112–127. [Google Scholar] [CrossRef]
- Lukassen, M.B. Off-Flavour Producing Bacteria in Aquaculture. Ph.D. Thesis, Aalborg University, Aalborg, Denmark, 2017. [Google Scholar]
- Schram, E.; van Kooten, T.; van de Heul, J.W.; Schrama, J.W.; Verreth, J.A.J.; Murk, A.J. Geosmin depuration from European eel (Anguilla anguilla) is not affected by the water renewal rate of depuration tanks. Aquac. Res. 2017, 48, 4646–4655. [Google Scholar] [CrossRef]
- Schram, E.; Schrama, J.W.; van Kooten, T.; Kwadijijk, C.J.A.F.; Kampen, H.; van de Heul, J.W.; Verreth, J.A.J.; Murk, A.J. Experimental validation of geosmin uptake in rainbow trout, Oncorhynchus mykiss (Waldbaum) suggests biotransformation. Aquac. Res. 2018, 49, 668–675. [Google Scholar] [CrossRef]
- Papp, Z.G.; Kerepeczki, É.; Pekár, F.; Gál, D. Natural origins of off-flavours in fish related to feeding habits. Water Sci. Technol. 2007, 55, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Powell, A.; Scolding, J.W.S. Direct application of ozone in aquaculture systems. Rev. Aquac. 2016, 10, 424–438. [Google Scholar] [CrossRef]
- Spiliotopoulou, A.; Rojas-Tirado, P.; Chhetri, R.K.; Kaarsholm, K.M.S.; Martin, R.; Pedersen, P.B.; Pedersen, L.-F.; Andersen, H.R. Ozonation control and effects of ozone on water quality in recirculating aquaculture systems. Water Res. 2018, 133, 289–298. [Google Scholar] [CrossRef]
- Fotiou, T.; Triantis, T.M.; Kaloudis, T.; Hiskia, A. Evaluation of the photocatalytic activity of TiO2 based catalysts for the degradation and mineralization of cyanobacterial toxins and water off-odor compounds under UV-A, solar and visible light. Chem. Eng. J. 2015, 261, 17–26. [Google Scholar] [CrossRef]
- Xue, Q.; Liu, Y.; Zhou, Q.; Utsumi, M.; Zhang, Z.; Sugiura, N. Photocatalytic degradation of geosmin by Pd nanoparticle modified WO3 catalyst under simulated solar light. Chem. Eng. J. 2016, 283, 614–621. [Google Scholar] [CrossRef]
- Azaria, S.; Nir, S.; van Rijn, J. Combined adsorption and degradation of the off-flavor compound 2-methylisoborneol in sludge derived from a recirculating aquaculture system. Chemosphere 2017, 169, 69–77. [Google Scholar] [CrossRef]
- Hanson, T.R. Off-Flavors in Aquaculture, ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2003. [Google Scholar] [CrossRef][Green Version]
- Zimba, P.V.; Schrader, K.K.; Hyldig, G.; Strobel, B.W.; Jørgensen, N.O.G. Evaluation of geosmin and 2-methylisoborneol off-flavour in smoked rainbow trout fillets using instrumental and sensory analyses. Aquac. Res. 2012, 43, 149–153. [Google Scholar] [CrossRef]
- Sarker, P.; Pilote, A.; Auffret, M.; Proulx, É.; Villemur, R.; Deschamps, M.-H.; Vandenberg, G. Reducing geosmin off-flavor compounds and waste outputs through dietary phosphorus management in rainbow trout aquaculture. Aquac. Environ. Interact. 2014, 6, 105–117. [Google Scholar] [CrossRef]
- Robertson, R.F.; Jauncey, K.; Beveridge, M.C.M.; Lawton, L.A. Depuration rates and the sensory threshold concentration of geosmin responsible for earthy-musty taint in rainbow trout, Onchorhynchus mykiss. Aquaculture 2005, 245, 89–99. [Google Scholar] [CrossRef]
- Petersen, M.A.; Hyldig, G.; Strobel, B.W.; Henriksen, N.; Jørgensen, N.O.G. Chemical and sensory quantification of geosmin and 2-methylisoborneol in rainbow trout (Oncorhynchus mykiss) from recirculated aquacultures in relation to concentrations in basin water. J. Agric. Food Chem. 2011, 59, 12561–12568. [Google Scholar] [CrossRef] [PubMed]
- Parinet, J.; Rodriguez, M.J.; Sérodes, J. Influence of water quality on the presence of off-flavour compounds (geosmin and 2-methylisoborneol). Water Res. 2010, 44, 5847–5856. [Google Scholar] [CrossRef] [PubMed]
- Blancheton, J.P.; Attramadal, K.J.K.; Michaud, L.; Roque d’Orbcastel, E.; Vadstein, O. Insight into bacterial population in aquaculture systems and its implication. Aquac. Eng. 2013, 53, 30–39. [Google Scholar] [CrossRef]
- Schram, E.; Verdegem, M.C.J.; Widjaja, R.T.O.B.H.; Kloet, C.J.; Foss, A.; Schelvis-Smit, R.; Roth, B.; Imsland, A.K. Impact of increased flow rate on specific growth rate of juvenile turbot (Scophthalmus maximus, Rafinesque 1810). Aquaculture 2009, 292, 46–52. [Google Scholar] [CrossRef]
- Guttman, L.; van Rijn, J. 2-Methylisoborneol and geosmin uptake by organic sludge derived from a recirculating aquaculture system. Water Res. 2009, 43, 474–480. [Google Scholar] [CrossRef]
- Davidson, J.; Schrader, K.; Ruan, E.; Swift, B.; Aalhus, J.; Juarez, M.; Wolters, W.; Burr, G.; Good, C.; Summerfelt, S.T. Evaluation of depuration procedures to mitigate the off-flavor compounds geosmin and 2-methylisoborneol from Atlantic salmon Salmo salar raised to market-size in recirculating aquaculture systems. Aquac. Eng. 2014, 61, 27–34. [Google Scholar] [CrossRef]
- Podduturi, R.; Petersen, M.A.; Vestergaard, M.; Jørgensen, N.O.G. Geosmin fluctuations and potential hotspots for elevated levels in recirculated aquaculture system (RAS): A case study from pikeperch (Stizostedion lucioperca) production in Denmark. Aquaculture 2020, 514, 734501. [Google Scholar] [CrossRef]
- Lukassen, M.B.; Saunders, A.M.; Sindilariu, P.D.; Nielsen, J.L. Quantification of novel geosmin-producing bacteria in aquaculture systems. Aquaculture 2017, 497, 304–310. [Google Scholar] [CrossRef]
- Koskela, J.; Jobling, M.; Savolainen, R. Influence of dietary fat level on feed intake, growth and fat deposition in the whitefish Coregonus lavaretus. Aquac. Int. 1998, 6, 95–102. [Google Scholar] [CrossRef]
- Quinton, C.D.; Kause, A.; Ruohonen, K.; Koskela, J. Genetic relationships of body composition and feed utilization traits in European whitefish (Coregonus lavaretus L.) and implications for selective breeding in fishmeal and soybean meal-based diet environments. J. Anim. Sci. 2007, 85, 3198–3208. [Google Scholar] [CrossRef]
- Wu, J.-T.; Jüttner, F. Differential partitioning of geosmin and 2-methylisoborneol between cellular constituents in Oscillatoria tenuis. Arch. Microbiol. 1988, 150, 580–583. [Google Scholar] [CrossRef]
- Directive 2010/63/EU of the European Parliament and of the Council on the Protection of Animals Used for Scientific Purposes. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32010L0063&from=EN (accessed on 1 March 2020).
- Hsu, J. Multiple Comparisons; Chapman and Hall, CRC: New York, NY, USA, 1996. [Google Scholar] [CrossRef]
- Kenward, M.G.; Roger, J.H. An improved approximation to the precision of fixed effects from restricted maximum likelihood. Comput. Stat. Data Anal. 2009, 53, 2583–2595. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Measurement | Unit |
|---|---|---|
| pH | 7.0 ± 0.3 | - |
| Temperature | 14.8 ± 0.9 | °C |
| Oxygen | 7.3 ± 0.6 | mg·L−1 |
| Alkalinity | 40.6 ± 19.2 | mg·L−1 |
| TAN | 0.39 ± 0.1 | mg·L−1 |
| NO2-N | 0.04 ± 0.02 | mg·L−1 |
| NO3-N | 32.8 ± 10.8 | mg·L−1 |
| SO42− | 11.7 ± 3.4 | mg·L−1 |
| Turbidity | 7.0 ± 3.2 | NTU |
| Characteristics | Value | Unit |
|---|---|---|
| Water renewal | 750 | L·kg−1 feed |
| Fish quantity per tank: European whitefish Rainbow trout | 3570–274 5500–83 | pcs |
| Fish density: European whitefish Rainbow trout | 1.0–56.1 7.5–54.5 | kg·tank−1 |
| Average fish size: European whitefish Rainbow trout | 0.28–242 1.36–657 | g |
| Feed quantity: European whitefish Rainbow trout | 0.04–0.33 0.04–0.50 | kg·d−1 |
| Feed pellet size: European whitefish Rainbow trout | 0.2–3.5 0.2–5.0 | mm |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindholm-Lehto, P.; Koskela, J.; Kaseva, J.; Vielma, J. Accumulation of Geosmin and 2-methylisoborneol in European Whitefish Coregonus Lavaretus and Rainbow Trout Oncorhynchus Mykiss in RAS. Fishes 2020, 5, 13. https://doi.org/10.3390/fishes5020013
Lindholm-Lehto P, Koskela J, Kaseva J, Vielma J. Accumulation of Geosmin and 2-methylisoborneol in European Whitefish Coregonus Lavaretus and Rainbow Trout Oncorhynchus Mykiss in RAS. Fishes. 2020; 5(2):13. https://doi.org/10.3390/fishes5020013
Chicago/Turabian StyleLindholm-Lehto, Petra, Juha Koskela, Janne Kaseva, and Jouni Vielma. 2020. "Accumulation of Geosmin and 2-methylisoborneol in European Whitefish Coregonus Lavaretus and Rainbow Trout Oncorhynchus Mykiss in RAS" Fishes 5, no. 2: 13. https://doi.org/10.3390/fishes5020013
APA StyleLindholm-Lehto, P., Koskela, J., Kaseva, J., & Vielma, J. (2020). Accumulation of Geosmin and 2-methylisoborneol in European Whitefish Coregonus Lavaretus and Rainbow Trout Oncorhynchus Mykiss in RAS. Fishes, 5(2), 13. https://doi.org/10.3390/fishes5020013