Transmission Strategies Used by Gyrodactylus gasterostei (Monogenea) on Its Host, the Three-Spined Stickleback Gasterosteus aculeatus


Abstract
1. Introduction
2. Results
2.1. Experiment 1. Parasite Migration from Dead Hosts
2.2. Experiment 2. Parasite Choice of Live or Dead Hosts
2.3. Experiment 3. One-to-One Fish Cohabitation
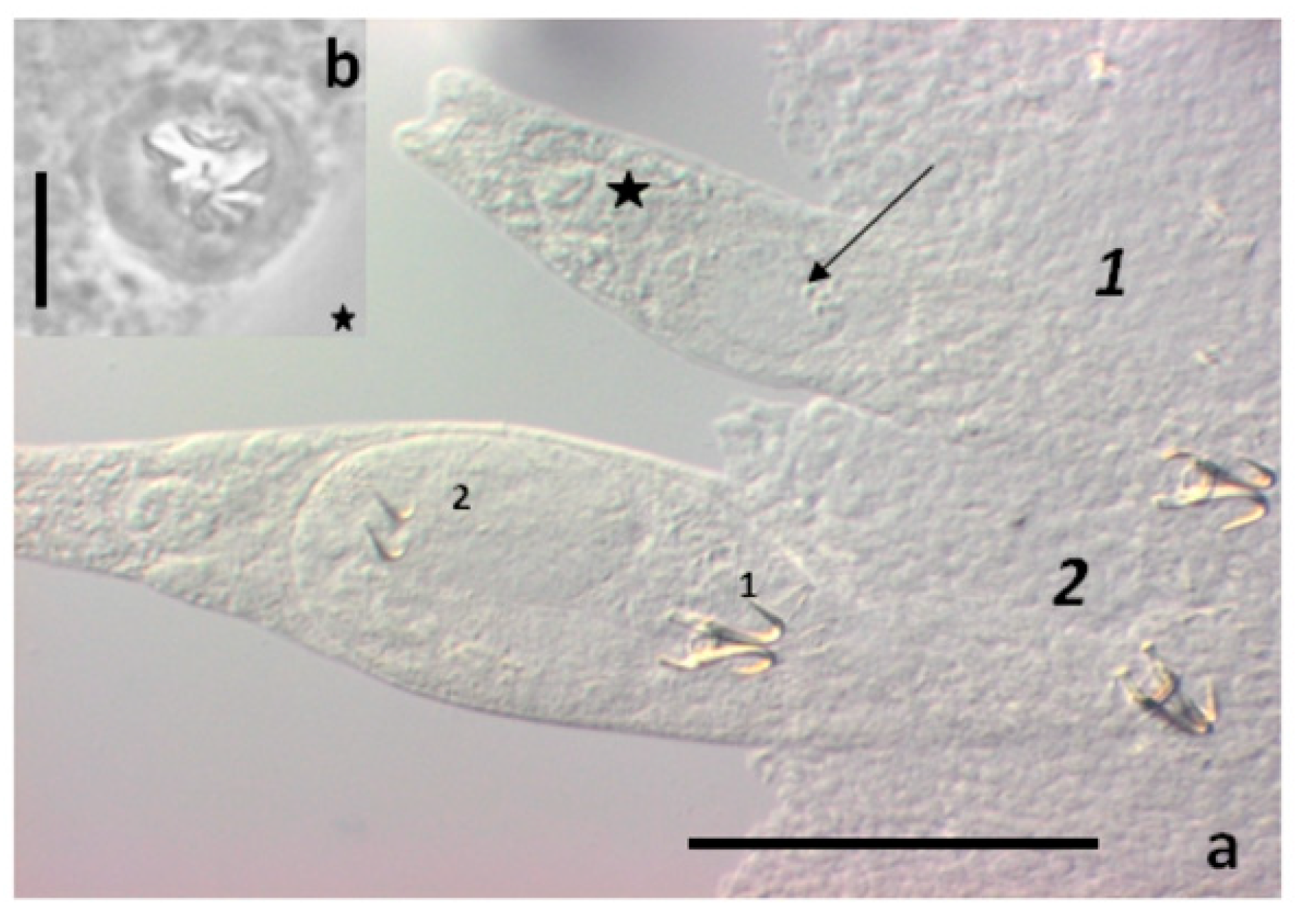
2.4. Experiment 4. Video Monitoring of Gyrodactylid Transmission In Vivo
3. Discussion
4. Materials and Methods
4.1. Source of Hosts and Parasites
4.2. Parasite Free-Hosts
4.3. Experiment 1. Parasite Migration from Dead Hosts
4.4. Experiment 2. Parasite Choice of Live or Dead Hosts
4.5. Experiment 3. One-to-One Fish Cohabitation
4.6. Experiment 4. Video Monitoring of Gyrodactylid Transmission In Vivo
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Harris, P.D. Interactions between reproduction and population biology in gyrodactylid monogeneans—A review. Bull. Fr. Peche Piscic. 1993, 1, 47–65. [Google Scholar] [CrossRef]
- Bakke, T.A.; Harris, P.D.; Cable, J. Host specificity dynamics: Observations on gyrodactylid monogeneans. Int. J. Parasitol. 2002, 32, 281–308. [Google Scholar] [CrossRef]
- Cable, J.; Harris, P.D. Gyrodactylids developmental biology historical review, current status and future trends. Int. J. Parasitol. 2002, 32, 255–280. [Google Scholar] [CrossRef]
- Harris, P.D. Changes in the site specificity of Gyrodactylus turnbulli Harris, 1986 (Monogenea) during infections of individual guppies (Poecilia reticulata Peters, 1859). Can. J. Zool. 1988, 66, 2854–2857. [Google Scholar] [CrossRef]
- Cable, J.; Scott, E.C.G.; Tinsley, R.C.; Harris, P.D. Behaviour favoring transmission in the viviparous monogenean Gyrodactylus turnbulli. J. Parasitol. 2002, 88, 183–184. [Google Scholar] [CrossRef]
- El-Naggar, M.M.; El-Naggar, A.M.; Kearn, G.C. Swimming in Gyrodactylus rysavyi (Monogenea: Gyrodactylidae) from the Nile catfish Clarias gariepinus. Acta Parasitol. 2004, 49, 102–107. [Google Scholar]
- Dmitrieva, E.V. Transmission triggers and pathways in Gyrodactylus sphinx (Monogenea, Gyrodactylidae). Vestnik Zool. 2003, 37, 67–72. [Google Scholar]
- Bakke, T.A.; Cable, J.; Harris, P.D. The biology of gyrodactylid monogeneans: The “Russian Doll-killers”. Adv. Parasitol. 2007, 64, 161–376. [Google Scholar] [PubMed]
- Grano-Maldonado, M.I. The Biological and Behavioural Basis of Host Selection in Transmission of Gyrodactylus (Monogenea). Ph.D. Thesis, Stirling Univeristy, Stirling, UK, 2011; p. 228. [Google Scholar]
- Grano-Maldonado, M.I.; Palaiokostas, C. Does the anaesthetic influence behavioural transmission of the monogenean Gyrodactylus gasterostei Gläser, 1974 off the host? Helminthologia 2015, 52, 144–147. [Google Scholar] [CrossRef][Green Version]
- Olstad, K.; Cable, J.; Robertsen, G.; Bakke, T.A. Unpredicted transmission strategy of Gyrodactylus salaris (Monogenea: Gyrodactylidae): Survival and infectivity of parasites on dead hosts. Parasitology 2006, 133, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.D. Observations on the development of the male reproductive system in Gyrodactylus gasterostei Gläser, 1974 (Monogenea, Gyrodactylidae). Parasitology 1985, 91, 519–529. [Google Scholar] [CrossRef]
- Raeymaekers, J.; Wegner, K.; Huyse, T.; Volckaert, F. Infection dynamics of the monogenean parasite Gyrodactylus gasterostei on sympatric and allopatric populations of the three-spined stickleback Gasterosteus aculeatus. Folia Parasitol. 2011, 58, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Grano-Maldonado, M.I.; Bron, J.; Longshaw, M.; Shinn, A. The accidental transfer of Gyrodactylus (Monogenea) during short duration fish transportation. Fish Pathol. 2011, 46, 71–79. [Google Scholar] [CrossRef]
- Grano-Maldonado, M.I. Ultrastructure of the external sensory apparatus of Gyrodactylus gasterostei Gläser, 1974. Microsc. Res. Tech. 2014, 77, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Grano-Maldonado, M.I. Gyrodactylus gasterostei a difficult meal to swallow for the three-spined sticklebacks, Gasterosteus aculeatus L. Scanning 2014, 36, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Brooker, A.J.; Grano-Maldonado, M.I.; Irving, S.; Bron, J.; Longshaw, M.; Shinn, A.P. The effect of octopaminergic compounds on the behaviour and transmission of Gyrodactylus. Parasite Vectors 2011, 4, 207. [Google Scholar] [CrossRef] [PubMed]
- Grano-Maldonado, M.I. Ultrastructure study of the stored lipid reserves in Gyrodactylus gasterostei Gläser, 1974 using confocal microscopy. J. Microsc. Ultrastruct. 2017. [Google Scholar] [CrossRef]
- Lester, R.J.G.; Adams, J.R. Gyrodactylus alexanderi: Reproduction, mortality and effect on its host Gasterosteus aculeatus. Can. J. Zool. 1974, 52, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Scott, M. Reproductive potential of Gyrodactylus bullatarudis (Monogenea) on guppies (Poecilia reticulata). Parasitology 1982, 85, 217–236. [Google Scholar] [CrossRef]
- Schelkle, B.; Paladini, G.; Shinn, A.; King, S.; Johnson, M.; van Oosterhout, C.; Mohammed, R.; Cable, J. Ieredactylus rivuli gen. et sp. nov. (Monogenea, Gyrodactylidae) from Ivulus hartii (Cyprinodontiformes, Rivulidae) in Trinida. Acta Parasitol. 2011, 56, 360–370. [Google Scholar] [CrossRef]
- Lavin, P.; McPhail, J. The evolution of freshwater diversity in the three-spined stickleback (Gasterosteus aculeatus): Site-specific differentation of trophic morphology. Can. J. Zool. 1985, 63, 2632–2638. [Google Scholar] [CrossRef]
- Bell, M.; Foster, S. The Evolutionary Biology of the Three spined Stickleback; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Grano-Maldonado, M.I.; Bruno de Sousa, C.; Rodriguez-Santiago, A. First insights into the ultrastructure of myosin and actin bands using Transmission Electron Microscopy in Gyrodactylus (Monogenea). J. Microsc. Ultrastruct. 2017. [Google Scholar] [CrossRef]
- Malmberg, G. The excretory system and marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Arch. Zool. 1970, 23, 1–237. [Google Scholar]
- De Roij, J.; Harris, P.; MacColl, A. Divergent resistance to a monogenean flatworm among three-spined stickleback populations. Funct. Ecol. 2011, 25, 211–217. [Google Scholar] [CrossRef]
- Harris, P.D.; Shinn, A.P.; Cable, J.; Bakke, T.A. Nominal species of the genus Gyrodactylus von Nordmann 1832 (Monogenea: Gyrodactylidae), with a list of principal host species. Syst. Parasitol. 2004, 59, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Shinn, A.P.; Hansen, H.; Olstad, K.; Bachmann, L.; Bakke, T.A. The use of morphometric characters to discriminate species of laboratory-reared and wild populations of Gyrodactylus salaris and G. thymalli (Monogenea). Folia Parasitol. 2004, 51, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.D.; Shinn, A.P.; Cable, J.; Bakke, T.A.; Bron, J.E. GyroDb: Gyrodactylid monogeneans on the web. Trends Parasitol. 2008, 24, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Shinn, A.P.; Harris, P.D.; Cable, J.; Bakke, T.A.; Paladini, G.; Bron, J.E. GyroDb—A Home for Gyrodactylids on the Web. Editors 2010. GyroDb. World Wide Web Electronic publicatIon, Version (06/2010). Available online: www.gyrodb.net (accessed on 1 August 2016).
- Konijnendijk, N.; Raeymaekers, J.A.; Vandeuren, S.; Jacquemin, L.; Volckaert, F.A. Testing for local adaptation in the Gasterosteus–Gyrodactylus host–parasite system. Evol. Ecol Res. 2013, 15, 489–502. [Google Scholar]
- Neiman, M.; Sharbel, T.F.; Schwander, T. Genetic causes of transitions from sexual reproduction to asexuality in plants and animals. J. Evol. Biol. 2014, 27, 1346–1359. [Google Scholar] [CrossRef] [PubMed]
- Lumme, J.; Ziętara, M.S. Horizontal transmission of the ectoparasite Gyrodactylus arcuatus (Monogenea: Gyrodactylidae) to the next generation of the three-spined stickleback Gasterosteus aculeatus. Folia Parasitol. 2018, 65, 006. [Google Scholar] [CrossRef] [PubMed]
- Tytell, E.D. Median fin function in bluegill sunfish, Lepomis macrochirus: Streamwise vortex structure during steady swimming. J. Exp. Biol. 2006, 209, 1516–1534. [Google Scholar] [CrossRef] [PubMed]
- Bozkurttas, M.; Dong, H.; Mittal, R.; Madden, P.; Lauder, G.V. Hydrodynamic performance of deformable fish fins and flapping foils. In Proceedings of the 44th AIAA Aerospace Sciences Meeting and Exhibit, Reno, Nevada, 9–12 January 2006. 2006 AIAA Paper 2006-1392. [Google Scholar]
- Drucker, E.G.; Lauder, G.V. Function of pectoral fins in rainbow trout: Behavioral repertoire and hydrodynamic forces. J. Exp. Biol. 2003, 206, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Pahor-Filho, E.; Miranda Filho, K.C.; Klosterhoff, M.; Romano, L.A.; Pereira Júnior, J. Histopathological and behaviour effects of formaldehyde treatment in juvenile mullet, Mugil liza (Valenciennes). Aquac. Res. 2015, 46, 3040–3045. [Google Scholar] [CrossRef]
- Harris, P.D. Ecological and genetic evidence for clonal reproduction in Gyrodactylus gasterostei Gläser, 1974. Int. J. Parasitol. 1998, 28, 1595–1607. [Google Scholar] [CrossRef]


{kind=link}
| PRS | [A] n = 240 | [B] n = 213 | p-Value (A vs. B) |
|---|---|---|---|
| N MCO/ND | 36 (15.0%) | 9 (4.2%) | <0.001 *** |
| N MCO/D | 136 (56.7%) | 53 (24.9%) | <0.001 *** |
| MCO/ND | 25 (10.4%) | 67 (31.5%) | <0.001 *** |
| MCO/D | 43 (17.92%) | 84 (39.4%) | <0.001 *** |
| Total N MCO | 172 (71.7%) | 62 (29.1%) | <0.001 *** |
| Total MCO | 68 (28.3%) | 151 (70.9%) | <0.001 *** |
| Location | Maturity Status of the Gyrodactylid | ||||
|---|---|---|---|---|---|
| N MCO/ND | N MCO/D | MCO/ND | MCO/D | Total | |
| Dead-un | 4 (4.2%) | 1 (0.2%) | 8 (2.1%) | 1 (0.3%) | 14 (1.1%) *** |
| Live-un | 1 (1.0%) | 6 (1.1%) | 11 (2.9%) | 5 (1.7%) | 23 (1.7%) *** |
| Live-inf | 79 (82.3%) | 510 (93.4%) | 346 (90.6%) | 274 (93.8%) | 1209 (91.9%) |
| Dislodged | 12 (12.5%) | 29 (5.3%) | 17 (4.5%) | 12 (4.1%) | 70 (5.3%) |
| Total | 96 | 546 | 382 | 292 | 1316 |
| Location | MCO/D | % | NMCO/D | % | MCO/ND | % | NMCO/ND | % | Total DS | Total | % |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Live-un | 46 | 33.3% | 21 | 15.21% | 49 | 35.50% | 22 | 15.94% | 138 (10.49%) | 1315 | 10.49 |
| Live-inf | 291 | 27.6% | 447 | 42.36% | 184 | 17.44% | 133 | 12.60% | 1055 (80.22%) | 80.22 | |
| Dislodged | 18 | 14.8% | 13 | 10.65% | 66 | 54.09% | 25 | 20.49% | 122 (9.2%) | 9.27 | |
| Dead-un | 104 | 35.86% | 56 | 19.3% | 85 | 29.31% | 45 | 15.51% | 290 (21.96%) | 1320 | 21.96 |
| Live-inf | 283 | 30.49% | 399 | 42.99% | 162 | 17.45% | 84 | 9.05% | 928 (70.30%) | 70.30 | |
| Dislodged | 25 | 24.50% | 9 | 8.82% | 53 | 51.96% | 15 | 14.70% | 102 (7.72%) | 7.72 | |
| Control | 314 | 27.86% | 430 | 38.15% | 243 | 21.56% | 140 | 12.42% | 1127 (90.74%) | 1242 | 90.74 |
| Dislodged | 28 | 24.34% | 25 | 21.73% | 47 | 40.86% | 15 | 13.04% | 115 (9.2%) | 9.25 | |
| Total | 1109 | 1400 | 889 | 479 | 3877 | 3877 | |||||
| Average | 7.701 | 9.72 | 6.17 | 3.32 | |||||||
| SD | 11.136 | 17.54 | 8.00 | 5.70 | |||||||
| % | 28.60 | 36.11 | 22.93 | 12.35 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grano-Maldonado, M.I.; Moreno-Navas, J.; Rodriguez-Santiago, M.A. Transmission Strategies Used by Gyrodactylus gasterostei (Monogenea) on Its Host, the Three-Spined Stickleback Gasterosteus aculeatus. Fishes 2018, 3, 20. https://doi.org/10.3390/fishes3020020
Grano-Maldonado MI, Moreno-Navas J, Rodriguez-Santiago MA. Transmission Strategies Used by Gyrodactylus gasterostei (Monogenea) on Its Host, the Three-Spined Stickleback Gasterosteus aculeatus. Fishes. 2018; 3(2):20. https://doi.org/10.3390/fishes3020020
Chicago/Turabian StyleGrano-Maldonado, Mayra I., Juan Moreno-Navas, and Maria Amparo Rodriguez-Santiago. 2018. "Transmission Strategies Used by Gyrodactylus gasterostei (Monogenea) on Its Host, the Three-Spined Stickleback Gasterosteus aculeatus" Fishes 3, no. 2: 20. https://doi.org/10.3390/fishes3020020
APA StyleGrano-Maldonado, M. I., Moreno-Navas, J., & Rodriguez-Santiago, M. A. (2018). Transmission Strategies Used by Gyrodactylus gasterostei (Monogenea) on Its Host, the Three-Spined Stickleback Gasterosteus aculeatus. Fishes, 3(2), 20. https://doi.org/10.3390/fishes3020020