
Molecular Characterization and Antiviral Function Against GCRV of Complement Factor D in Barbel Chub (Squaliobarbus curriculus)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish, Cell Line, and Virus
2.2. RNA Extraction, cDNA Synthesis, and Quantitative PCR
2.3. Cloning and Bioinformatics Analysis of ScDF
2.4. Plasmid Construction
2.5. Overexpression and Viral Challenge Assay
2.6. Recombinant Protein Incubation Assay
2.7. Statistical Analysis
3. Results
3.1. Molecular Characterization of ScDF Reveals a Unique Glycosylation Site
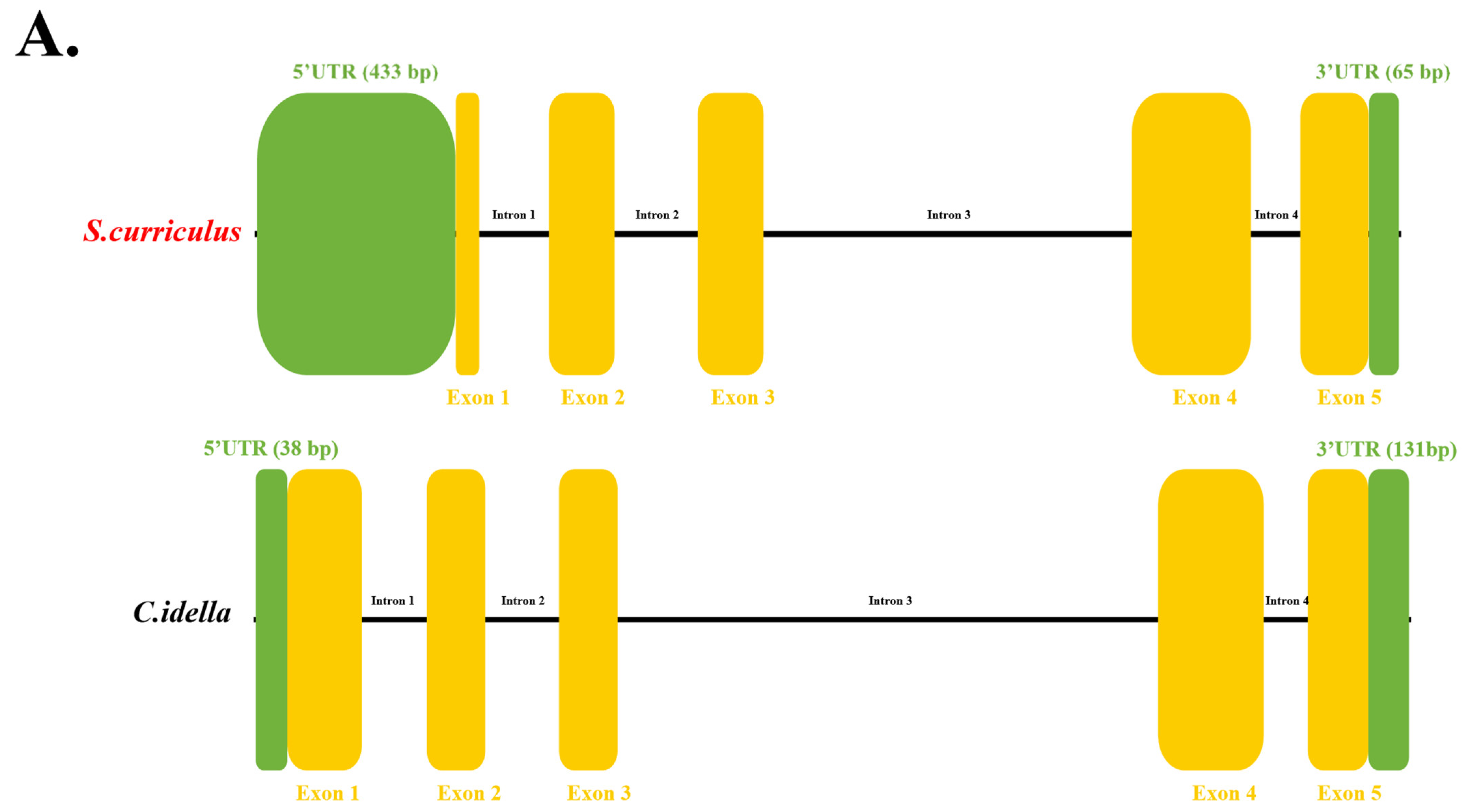
3.2. ScDF Is an Extracellular Protein with Conserved Domain Architecture
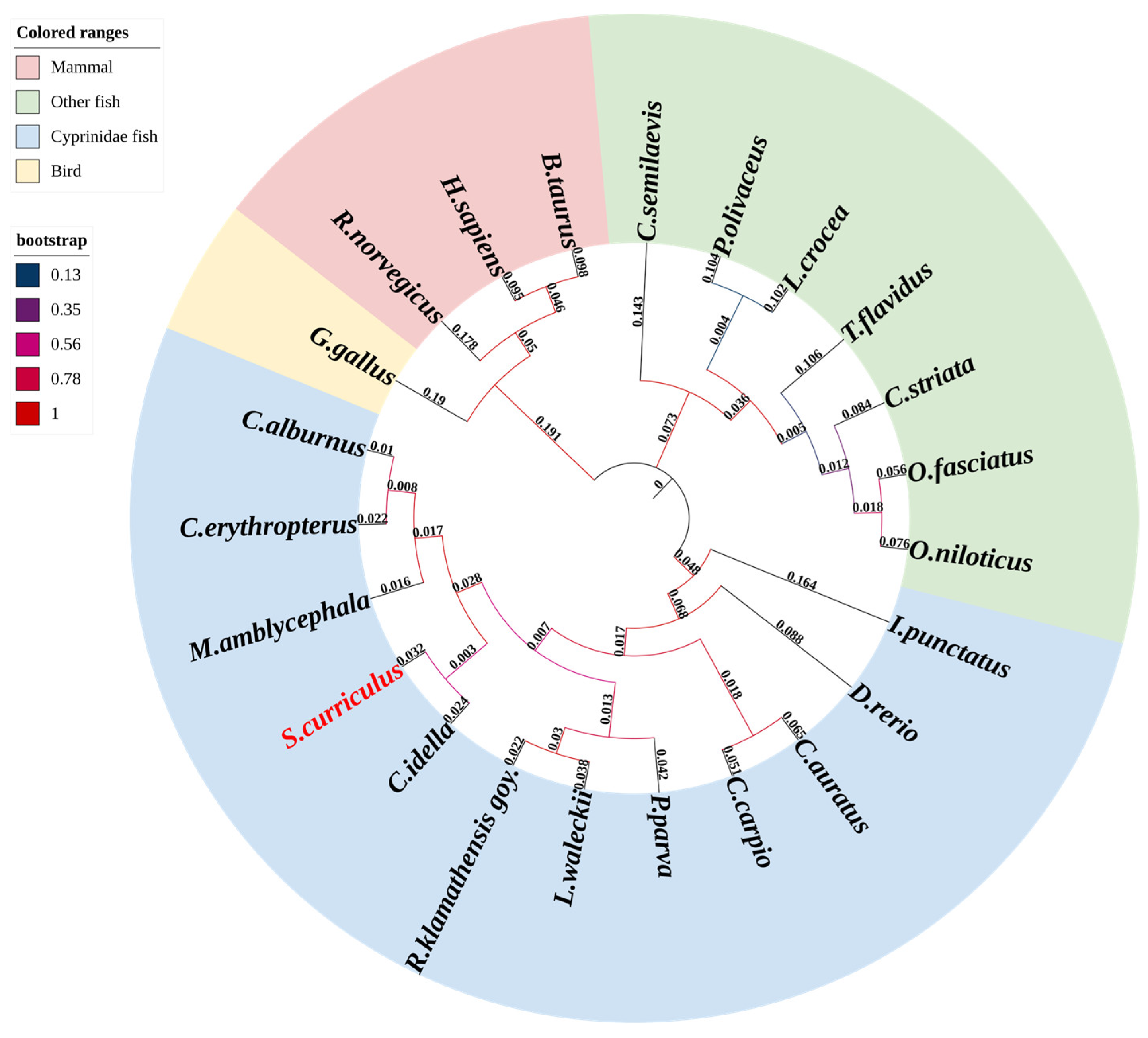
3.3. Evolutionary Conservation of ScDF Within Vertebrate Complement Systems
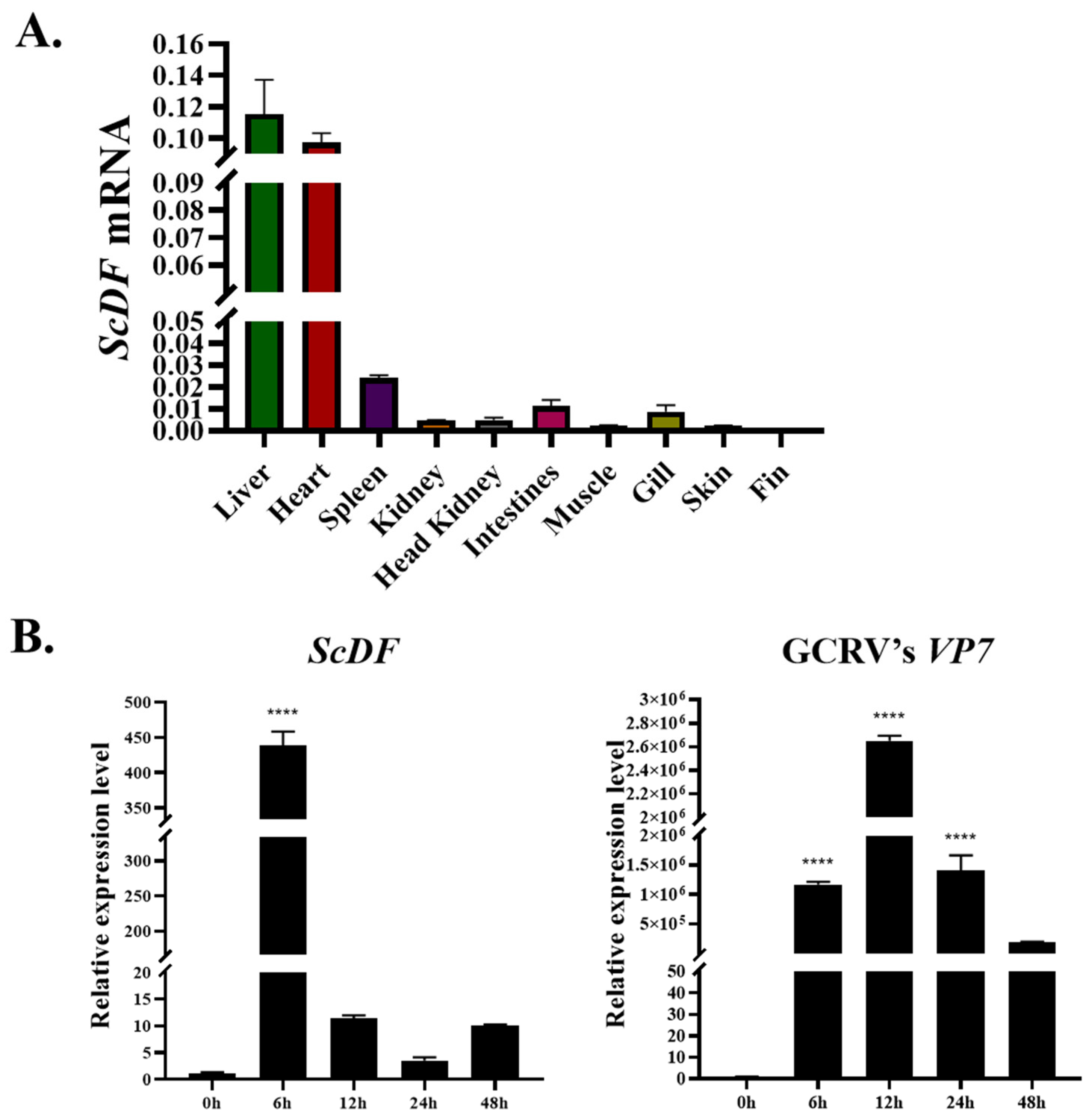
3.4. Hepatic Dominance of ScDF and Its Early Response to GCRV Outpace CiDF
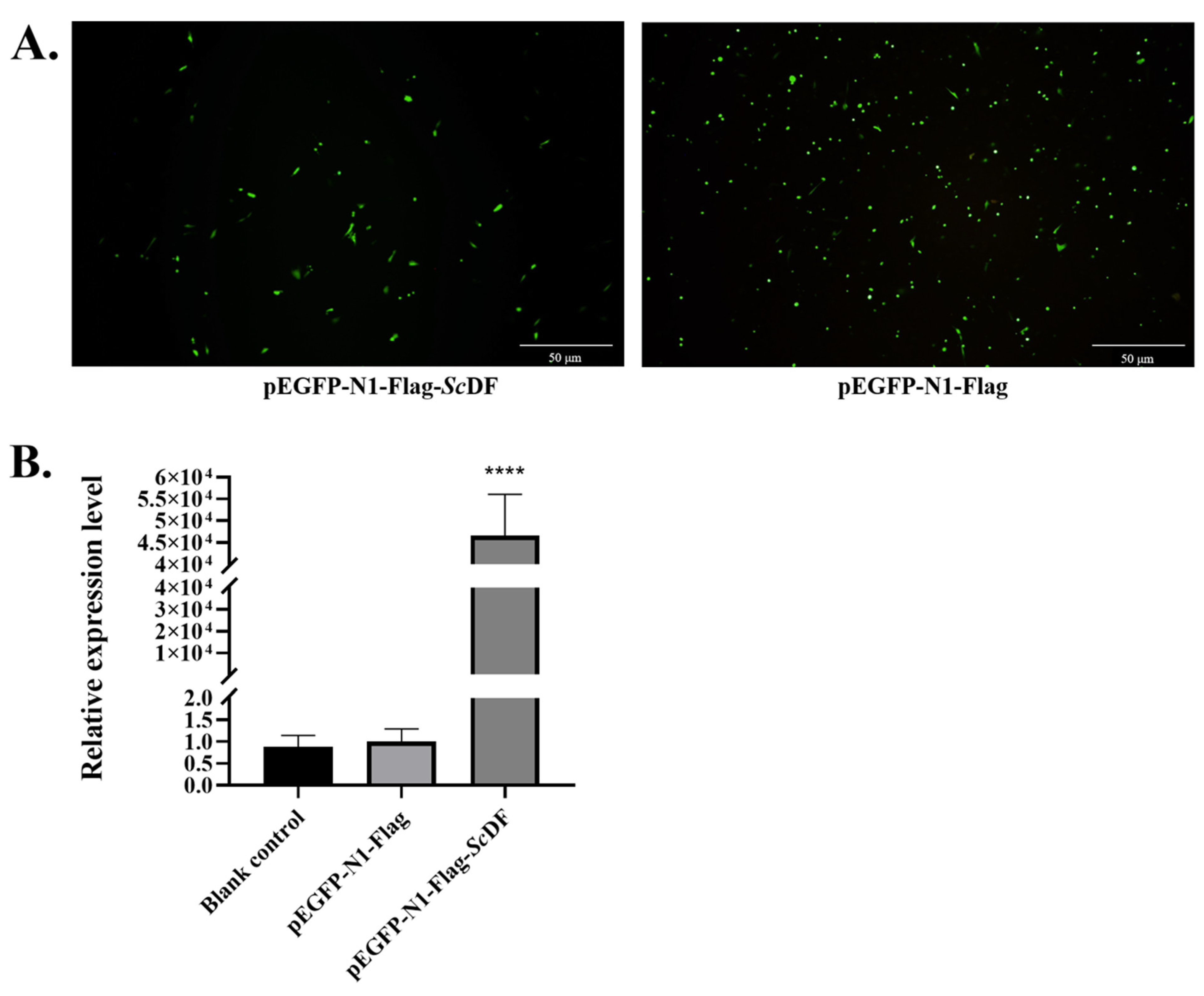
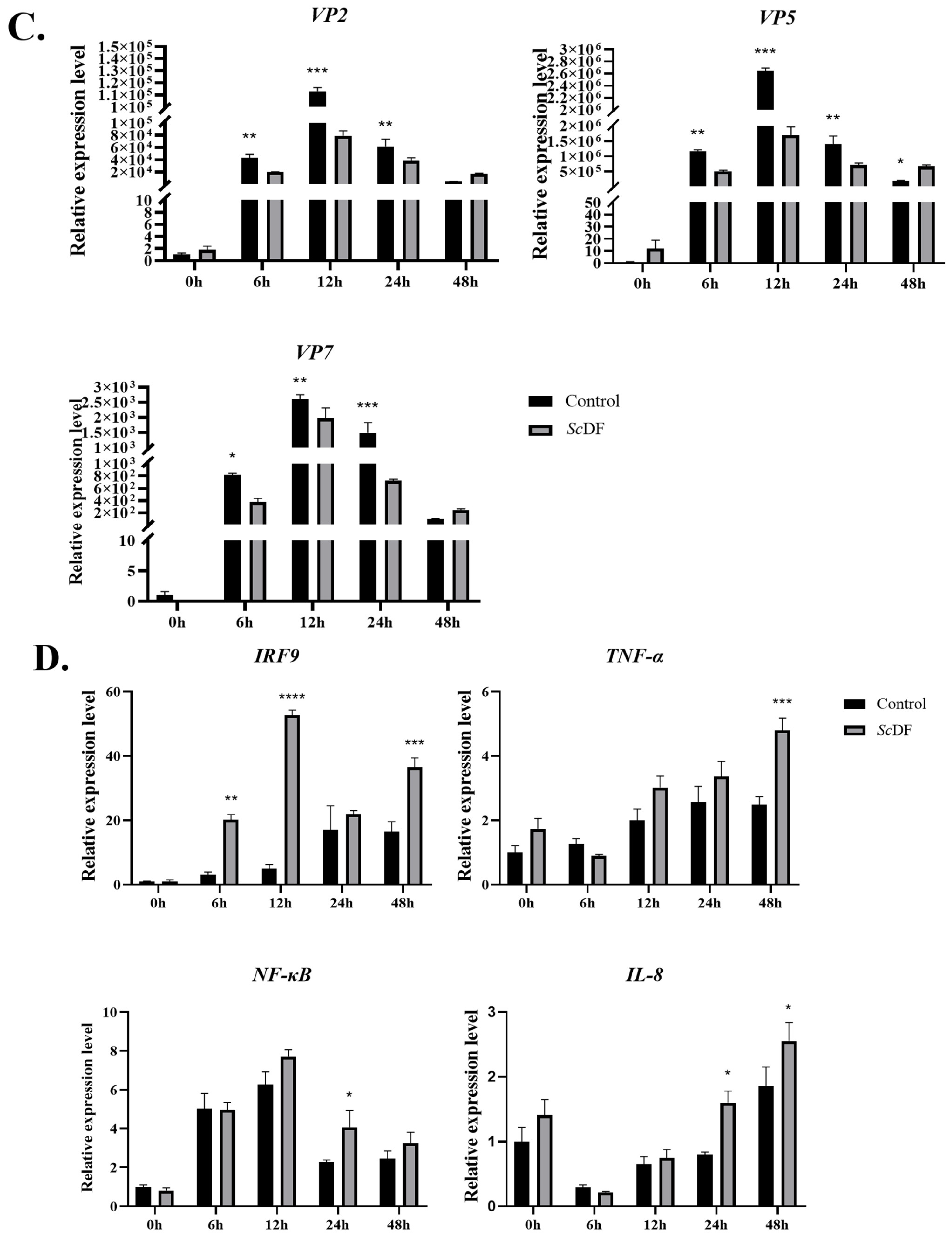
3.5. ScDF Overexpression Suppresses GCRV Replication
3.6. Recombinant ScDF Protein Synergistically Activates Interferon and Complement Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.; Zeng, W.; Liu, C.; Zhang, C.; Wang, Y.; Shi, C.; Wu, S. Complete genome sequence of a reovirus isolated from grass carp, indicating different genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhang, M.; Xu, Y.; Qin, B.; Yang, H.; Wei, R.; Xiao, T. Structural and functional characteristics of TLR19 in barbel chub compared to TLR19 in grass carp. Int. J. Mol. Sci. 2025, 26, 3103. [Google Scholar] [CrossRef]
- Yang, H.; Deng, Y.; Ding, C.H.; Xu, B.H.; Lv, Z.; Xiao, T. Serum proteomic profiling of grass carp survived from GCRV infection. Acta Hydrobiol. Sin. 2024, 48, 304–314. [Google Scholar] [CrossRef]
- Caputo, M.B.; Elias, J.; Cesar, G.; Alvarez, M.G.; Laucella, S.A.; Albareda, M.C. Role of the complement system in the modulation of T-cell responses in chronic chagas disease. Front. Cell Infect. Microbiol. 2022, 12, 910854. [Google Scholar] [CrossRef]
- Dobó, J.; Kocsis, A.; Farkas, B.; Demeter, F.; Cervenak, L.; Gál, P. The lectin pathway of the complement system-activation, regulation, disease connections and interplay with other (proteolytic) systems. Int. J. Mol. Sci. 2024, 25, 1566. [Google Scholar] [CrossRef]
- Mortensen, S.A.; Sander, B.; Jensen, R.K.; Pedersen, J.S.; Golas, M.M.; Jensenius, J.C.; Hansen, A.G.; Thiel, S.; Andersen, G.R. Structure and activation of C1, the complex initiating the classical pathway of the complement cascade. Proc. Natl. Acad. Sci. USA 2017, 114, 986–991. [Google Scholar] [CrossRef]
- Yadav, M.K.; Maharana, J.; Yadav, R.; Saha, S.; Sarma, P.; Soni, C.; Singh, V.; Saha, S.; Ganguly, M.; Li, X.X.; et al. Molecular basis of anaphylatoxin binding, activation, and signaling bias at complement receptors. Cell 2023, 186, 4956–4973. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, M.; Smith, S.L. Complement system of bony and cartilaginous fish. Fish Shellfish Immunol. 2000, 10, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Bavia, L.; Santiesteban-Lores, L.E.; Carneiro, M.C.; Prodocimo, M.M. Advances in the complement system of a teleost fish, Oreochromis niloticus. Fish Shellfish Immunol. 2022, 123, 61–74. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J.O. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Dang, Y.F.; Shen, Y.B.; Xu, X.Y.; Wang, S.T.; Meng, X.Z.; Li, L.S.; Zhang, M.; Hu, M.Y.; Lv, L.Q.; Wang, R.Q.; et al. Mannan-binding lectin-associated serine protease-1 (MASP-1) mediates immune responses against Aeromonas hydrophila in vitro and in vivo in grass carp. Fish Shellfish Immunol. 2017, 66, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Mutsuro, J.; Tanaka, N.; Kato, Y.; Dodds, A.W.; Yano, T.; Nakao, M. Two divergent isotypes of the fourth complement component from a bony fish, the common carp (Cyprinus carpio). J. Immunol. 2005, 175, 4508–4517. [Google Scholar] [CrossRef]
- Dang, Y.; Shen, Y.; Xu, X.; Wang, S.; Meng, X.; Zhang, M.; Lv, L.; Wang, R.; Li, J. Complement component Bf/C2b gene mediates immune responses against Aeromonas hydrophila in grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2018, 74, 509–516. [Google Scholar] [CrossRef]
- Risitano, A.M.; Frieri, C.; Urciuoli, E.; Marano, L. The complement alternative pathway in paroxysmal nocturnal hemoglobinuria: From a pathogenic mechanism to a therapeutic target. Immunol. Rev. 2023, 313, 262–278. [Google Scholar] [CrossRef]
- Goshima, M.; Sekiguchi, R.; Matsushita, M.; Nonaka, M. The complement system of elasmobranches revealed by liver transcriptome analysis of a hammerhead shark, Sphyrna zygaena. Dev. Comp. Immunol. 2016, 61, 13–24. [Google Scholar] [CrossRef]
- Biesma, D.H.; Hannema, A.J.; van Velzen-Blad, H.; Mulder, L.; van Zwieten, R.; Kluijt, I.; Roos, D. A family with complement factor D deficiency. J. Clin. Investig. 2001, 108, 233–240. [Google Scholar] [CrossRef]
- Qi, H.; Wei, J.; Gao, Y.; Yang, Y.; Li, Y.; Zhu, H.; Su, L.; Su, X.; Zhang, Y.; Yang, R. Reg4 and complement factor D prevent the overgrowth of E. coli in the mouse gut. Commun. Biol. 2020, 3, 483. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Fan, J.; Wang, W.; Wang, H.; Liu, H. Molecular characterization, expression and antimicrobial activity of complement factor D in Megalobrama amblycephala. Fish Shellfish Immunol. 2019, 89, 43–51. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, H.; Liu, S.; Sun, F.; Peatman, E.; Kucuktas, H.; Kaltenboeck, L.; Feng, T.; Zhang, H.; Niu, D.; et al. Alternative complement pathway of channel catfish (Ictalurus punctatus): Molecular characterization, mapping and expression analysis of factors Bf/C2 and Df. Fish Shellfish Immunol. 2012, 32, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Godahewa, G.I.; Perera, N.C.; Bathige, S.D.; Nam, B.H.; Noh, J.K.; Lee, J. Complement factor D homolog involved in the alternative complement pathway of rock bream (Oplegnathus fasciatus): Molecular and functional characterization and immune responsive mRNA expression analysis. Fish Shellfish Immunol. 2016, 55, 423–433. [Google Scholar] [CrossRef]
- Byon, J.Y.; Ohira, T.; Hirono, I.; Aoki, T. Comparative immune responses in Japanese flounder, Paralichthys olivaceus after vaccination with viral hemorrhagic septicemia virus (VHSV) recombinant glycoprotein and DNA vaccine using a microarray analysis. Vaccine 2006, 24, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Xiao, T.; Qin, B.; Xu, B.; Lv, Z.; Wang, H. Functional identification of complement factor D and analysis of its expression during GCRV infection in grass carp (Ctenopharyngodon idella). Int. J. Mol. Sci. 2021, 22, 12011. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, A.; Pei, Y.; Chu, P.; Li, Y.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Differences in responses of grass carp to different types of grass carp reovirus (GCRV) and the mechanism of hemorrhage revealed by transcriptome sequencing. BMC Genom. 2017, 18, 452. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhong, X.; Chen, M.; Ding, M.; Zhong, M.; Li, B.; Wang, Y.; Fu, S.; Yin, X.; Guo, Z.; Ye, J. C1r and C1s from Nile tilapia (Oreochromis niloticus): Molecular characterization, transcriptional profiling upon bacterial and IFN-gamma inductions and potential role in response to bacterial infection. Fish Shellfish Immunol. 2017, 70, 240–251. [Google Scholar] [CrossRef]
- Liao, Z.; Wan, Q.; Yuan, G.; Su, J. The systematic identification and mRNA expression profiles post viral or bacterial challenge of complement system in grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2019, 86, 107–115. [Google Scholar] [CrossRef]
- Chen, G.; Xiong, L.; Wang, Y.; He, L.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Different responses in one-year-old and three-year-old grass carp reveal the mechanism of age restriction of GCRV infection. Fish Shellfish Immunol. 2019, 86, 702–712. [Google Scholar] [CrossRef]
- Sasawatari, S.; Okamoto, Y.; Kumanogoh, A.; Toyofuku, T. Blockade of N-glycosylation promotes antitumor immune response of T cells. J. Immunol. 2020, 204, 1373–1385. [Google Scholar] [CrossRef]
- Blouin, C.M.; Hamon, Y.; Gonnord, P.; Boularan, C.; Kagan, J.; Viaris de Lesegno, C.; Ruez, R.; Mailfert, S.; Bertaux, N.; Loew, D.; et al. Glycosylation-dependent IFN-gammaR partitioning in lipid and actin nanodomains is critical for JAK activation. Cell 2016, 166, 920–934. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.; Meng, B.; Gao, Y.; Xue, Z.; He, M.; Jiang, Y.; Dai, X.; Yan, D.; Fang, X. Identification of dysregulated complement activation pathways driven by N-glycosylation alterations in T2D patients. Front. Chem. 2021, 9, 677621. [Google Scholar] [CrossRef]
- Pascoal, C.; Francisco, R.; Mexia, P.; Pereira, B.L.; Granjo, P.; Coelho, H.; Barbosa, M.; Dos Reis Ferreira, V.; Videira, P.A. Revisiting the immunopathology of congenital disorders of glycosylation: An updated review. Front. Immunol. 2024, 15, 1350101. [Google Scholar] [CrossRef]
- Šoić, D.; Keser, T.; Štambuk, J.; Kifer, D.; Pociot, F.; Lauc, G.; Morahan, G.; Novokmet, M.; Gornik, O. High-throughput human complement C3 N-glycoprofiling identifies markers of early onset type 1 diabetes mellitus in children. Mol. Cell Proteom. 2022, 21, 100407. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, N.; Naruse, K.; Shima, A.; Nonaka, M.; Sasaki, M. Molecular cloning and linkage analysis of complement C3 and C4 genes of the Japanese medaka fish. Immunogenetics 2000, 51, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, R.; Li, Y.; Xiao, T. Molecular cloning and functional characterization of interferon regulatory factor 7 of the barbel chub, Squaliobarbus curriculus. Fish Shellfish Immunol. 2017, 69, 185–194. [Google Scholar] [CrossRef]
- Ungrová, L.; Geryk, J.; Kohn, M.; Kučerová, D.; Krchlíková, V.; Hron, T.; Pečenka, V.; Pajer, P.; Gáliková, E.; Pecnová, Ľ.; et al. Avian interferon regulatory factor (IRF) family reunion: IRF3 and IRF9 found. BMC Biol. 2025, 23, 180. [Google Scholar] [CrossRef]
- Guo, C.; Yang, X.; Wu, Y.; Yang, L.; Mi, S.; Liu, Z.; Jia, K.; Huang, Y.; Weng, S.; Yu, X.; et al. Involvement of caveolin-1 in the JAK–STAT signaling pathway and infectious spleen and kidney necrosis virus infection in mandarin fish (Siniperca chuatsi). Mol. Immunol. 2011, 48, 992–1000. [Google Scholar] [CrossRef]
- Wang, W.L.; Liu, W.; Gong, H.Y.; Hong, J.R.; Lin, C.C.; Wu, J.L. Activation of cytokine expression occurs through the TNFalpha/NF-kappaB-mediated pathway in birnavirus-infected cells. Fish Shellfish Immunol. 2011, 31, 10–21. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, Y.; Gao, Y.; Wang, X.; Zhou, H.; Zhang, A. Insights into the functional role of grass carp IL-8 in head kidney leukocytes: Pro-inflammatory effects and signalling mechanisms. J. Fish Biol. 2022, 100, 192–202. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Applications | Sequences (5′-3′) | Primer Names |
|---|---|---|
| qPCR | CTGATATTTGCCTCTTTGC | ScDF-qF |
| qPCR | ATTCCACTGAAGTGAAGCCATGTAC | ScDF-qR |
| qPCR | GGCTGTGCTGTCCCTGTATG | β-actin-qF |
| qPCR | CTCTGGGCACCTGAACCTCT | β-actin-qR |
| qPCR | CGGCATTTACTTCGTCTA | TNF-α-qF |
| qPCR | TCCATAGGAATCGGAATAG | TNF-α-qR |
| qPCR | TCAAGACGAGAACGGAGAC | NF-κB-qF |
| qPCR | CTGCTAGGTGCAAAGGAGT | NF-κB-qR |
| qPCR | ATCAAGGATCCCATTCCGCCTTCA | VP2-qF |
| qPCR | TTAGAGGATCGTGCCATTGAGGGT | VP2-qR |
| qPCR | AGACGACATACAAGGGCGAAGT | VP5-qF |
| qPCR | GGCGGAAAGCATACCAGATT | VP5-qR |
| qPCR | ACCACCAACTTTGATCACGCTGAG | VP7-qF |
| qPCR | AGCGTGGGAGTCTTGAATGGTCTT | VP7-qR |
| qPCR | TGAGTCTTAGAGGTCTGGGTG | IL-8-qF |
| qPCR | TTACAGTGAGGGCTGGGAG | IL-8-qR |
| qPCR | CCTCCATCTCCACCGTCTC | IRF9-qF |
| qPCR | AAGGGCAGCAATTCGTTAA | IRF9-qR |
| qPCR | CTCAGTCTACTATGGTCGTG | C3-qF |
| qPCR | AAAGGTCCAGCCTATGTC | C3-qR |
| qPCR | TGAGACGCTGGTCAAGAAC | IFN1-qF |
| qPCR | ACTTTCCACCCATTTCCCT | IFN1-qR |
| qPCR | TCCCGAAATCTGCACTGCAA | IFN2-qF |
| qPCR | CAGCTGGTTCCAGGACTCTG | IFN2-qR |
| qPCR | GTTGGCGAACTCAAGAGGCT | ISG15-qF |
| qPCR | CTGAGTCCGAGTTCAGTCCG | ISG15-qR |
| cDNA cloning | ATGAATAGACTGATATTTGCCTCTTTGCTG | ScDF-F |
| cDNA cloning | TTACTGGGTGGTTGTGCTGTCAAT | ScDF-R |
| Overexpression vector construction | GGACTCAGATCTCGAGATGAATAGACTGATATTTGCCTCTTTGCTG | ScDF-F1 |
| Overexpression vector construction | TGGTGGCGGCGGATCCCTGGGTGGTTGTGCTGTCAAT | ScDF-R1 |
| Prokaryotic vector construction | TGGTATCGAAGGTAGGCATATGATTACAGGAGGAAGTGAGGCTGAT | ScDF-F2 |
| Prokaryotic vector construction | AGCAGAGATTACCTATCTAGATTACTGGGTGGTTGTGCTGTCA | ScDF-R2 |
| Species | GenBank Accession Numbers |
|---|---|
| Ctenopharyngodon idella | AHB81535.1 |
| Megalobrama amblycephala | XP_048060119.1 |
| Rhinichthys klamathensis goyatoka | XP_056103980.1 |
| Culter alburnus | KAK9964805.1 |
| Chanodichthys erythropterus | XP_067241195.1 |
| Pseudorasbora parva | XP_067277182.1 |
| Cyprinus carpio | XP_042624654.1 |
| Leuciscus waleckii | XDV50082.1 |
| Carassius auratus | XP_026079875.1 |
| Danio rerio | NP_001018368.1 |
| Ictalurus punctatus | AEW10547.1 |
| Oreochromis niloticus | XP_003447819.1 |
| Larimichthys crocea | XP_010739031.3 |
| Oplegnathus fasciatus | AIZ96981.1 |
| Cynoglossus semilaevis | XP_008314450.1 |
| Paralichthys olivaceus | ACV89350.1 |
| Takifugu flavidus | TWW72489.1 |
| Channa striata | SSC14279.1 |
| Gallus gallus | XP_040548688.1 |
| Bos taurus | NP_001029427.1 |
| Rattus norvegicus | NP_001071110.1 |
| Homo sapiens | NP_001919.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Lv, Z.; Wei, Y.; Zhang, M.; Yang, H.; Huang, C.; Xiao, T.; Li, Y. Molecular Characterization and Antiviral Function Against GCRV of Complement Factor D in Barbel Chub (Squaliobarbus curriculus). Fishes 2025, 10, 370. https://doi.org/10.3390/fishes10080370
Xiao Y, Lv Z, Wei Y, Zhang M, Yang H, Huang C, Xiao T, Li Y. Molecular Characterization and Antiviral Function Against GCRV of Complement Factor D in Barbel Chub (Squaliobarbus curriculus). Fishes. 2025; 10(8):370. https://doi.org/10.3390/fishes10080370
Chicago/Turabian StyleXiao, Yu, Zhao Lv, Yuling Wei, Mengyuan Zhang, Hong Yang, Chao Huang, Tiaoyi Xiao, and Yilin Li. 2025. "Molecular Characterization and Antiviral Function Against GCRV of Complement Factor D in Barbel Chub (Squaliobarbus curriculus)" Fishes 10, no. 8: 370. https://doi.org/10.3390/fishes10080370
APA StyleXiao, Y., Lv, Z., Wei, Y., Zhang, M., Yang, H., Huang, C., Xiao, T., & Li, Y. (2025). Molecular Characterization and Antiviral Function Against GCRV of Complement Factor D in Barbel Chub (Squaliobarbus curriculus). Fishes, 10(8), 370. https://doi.org/10.3390/fishes10080370