
Asparagopsis taxiformis Feed Supplementation as a Tool to Improve the Resilience of Farmed Diplodus sargus to Marine Heatwave Events—A Metabolomics Approach

 , ,
, ,  , , ,
, , ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Seaweed Collection and Experimental Diets
2.3. Organisms and Acclimation Period
2.4. Experimental Design and Fish Rearing Conditions
2.5. Fish Sampling
2.6. Extraction of Primary Metabolites from D. sargus Liver Tissues
2.7. GC-TOF-MS Primary Metabolite Profiling Analysis
2.8. Metabolite Data Processing and Statistical Analysis
3. Results
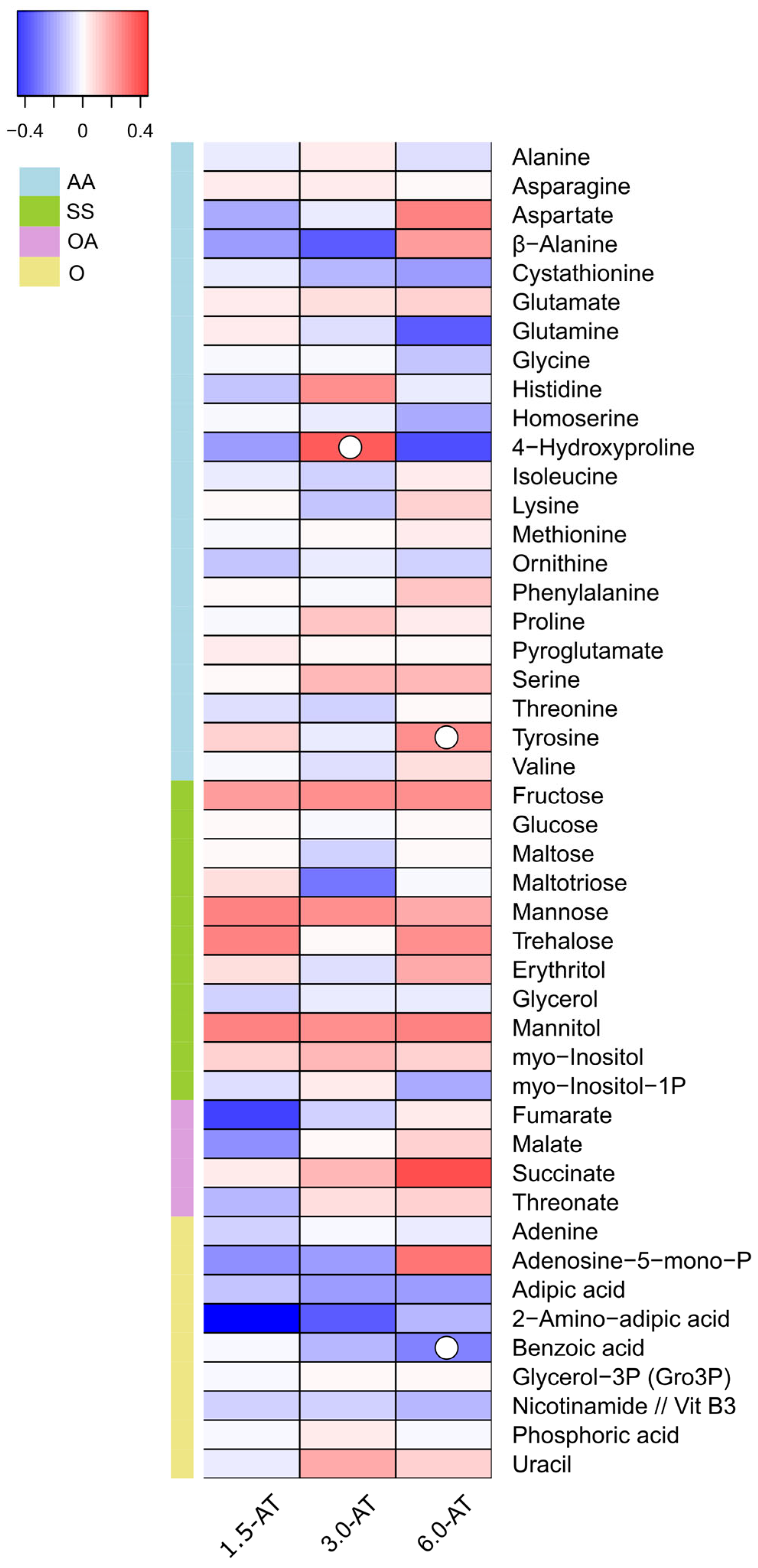
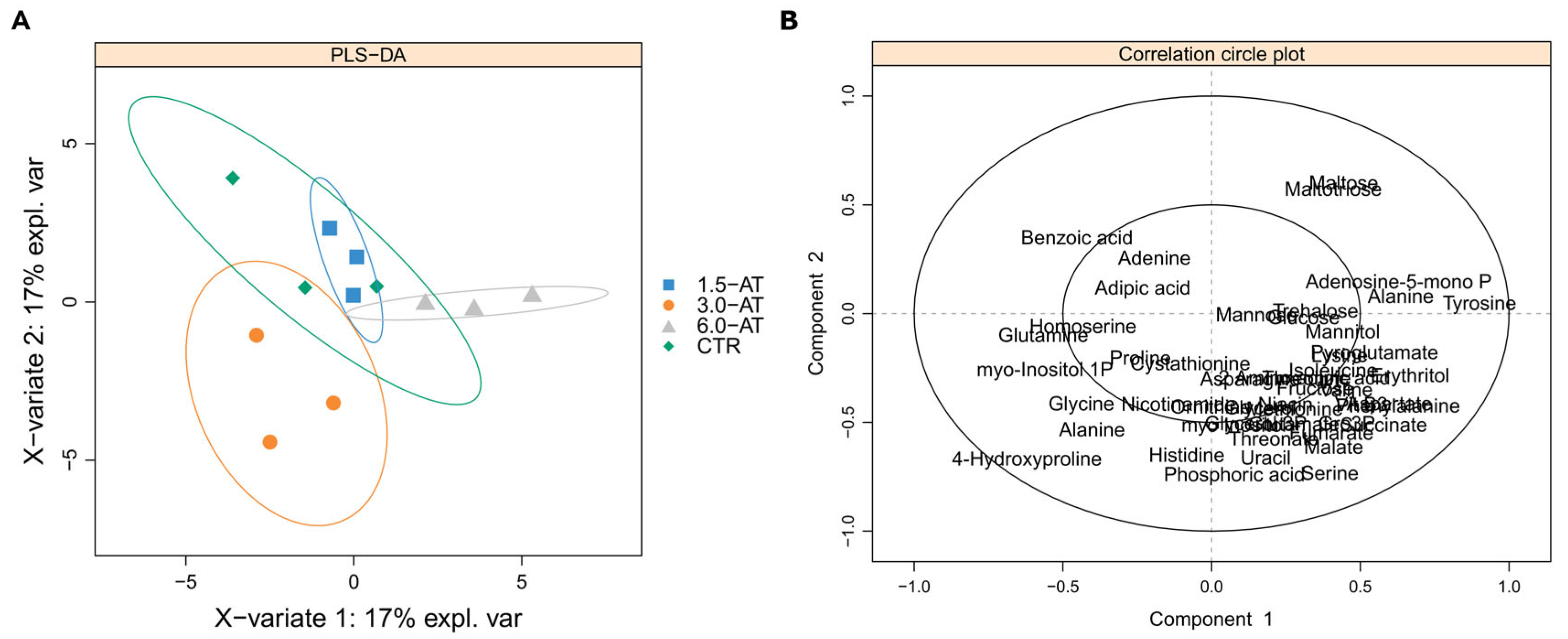
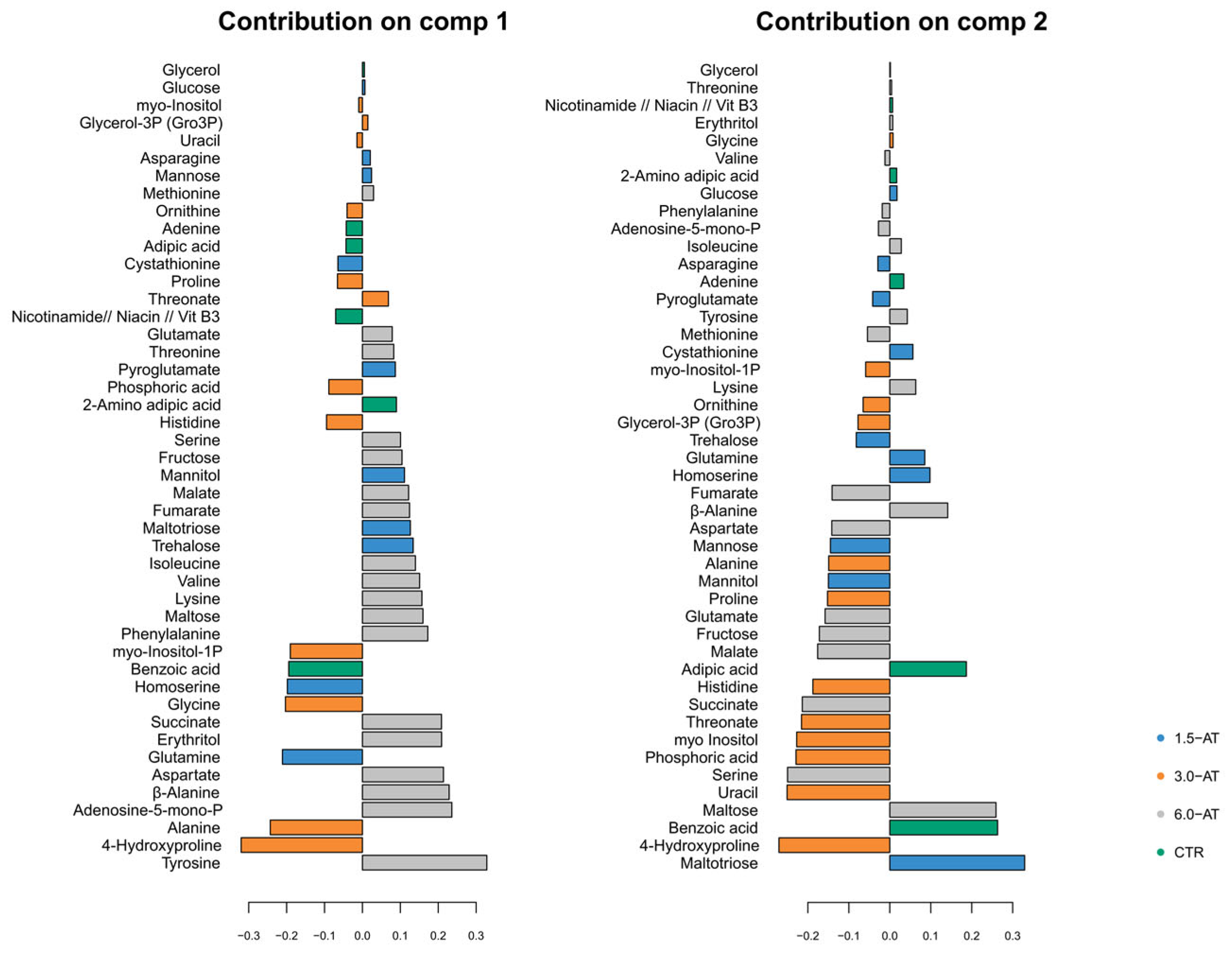
3.1. GC-TOF-MS Primary Metabolite Profiling of Diplodus Sargus Liver Tissues When Supplemented with A. taxiformis
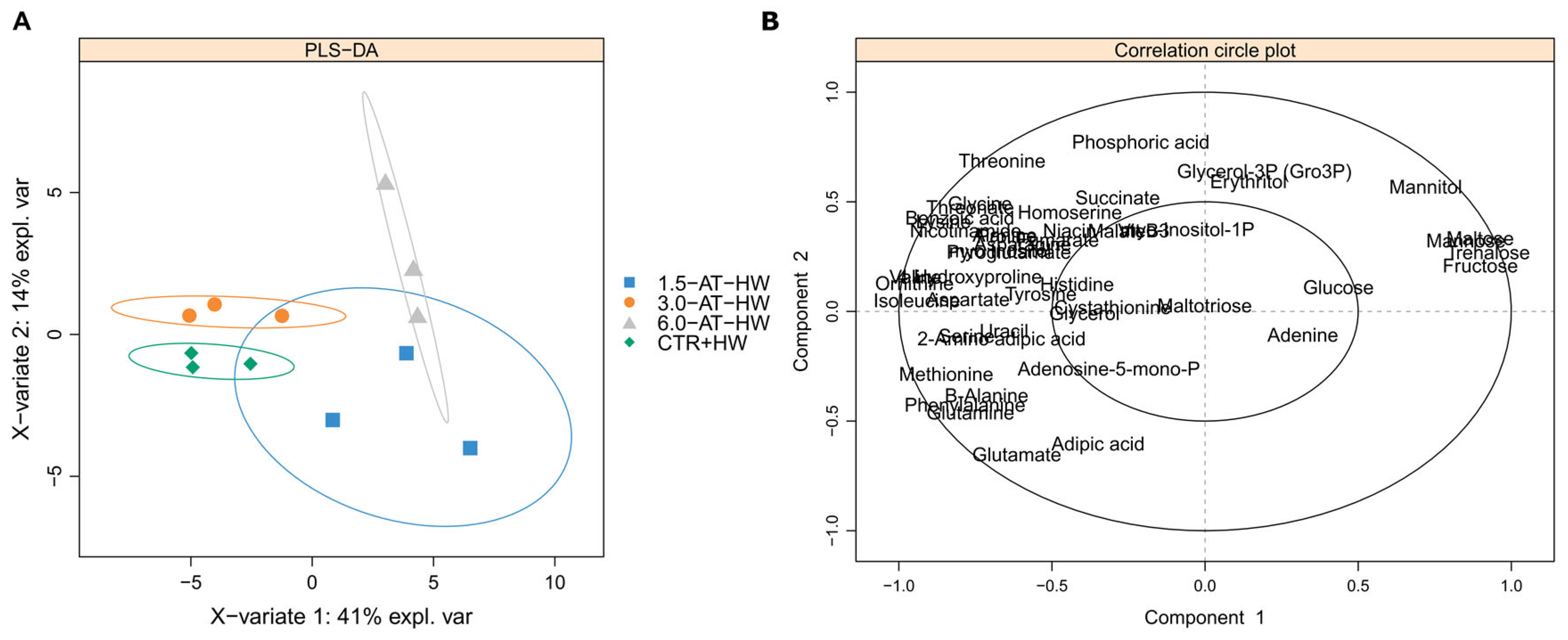
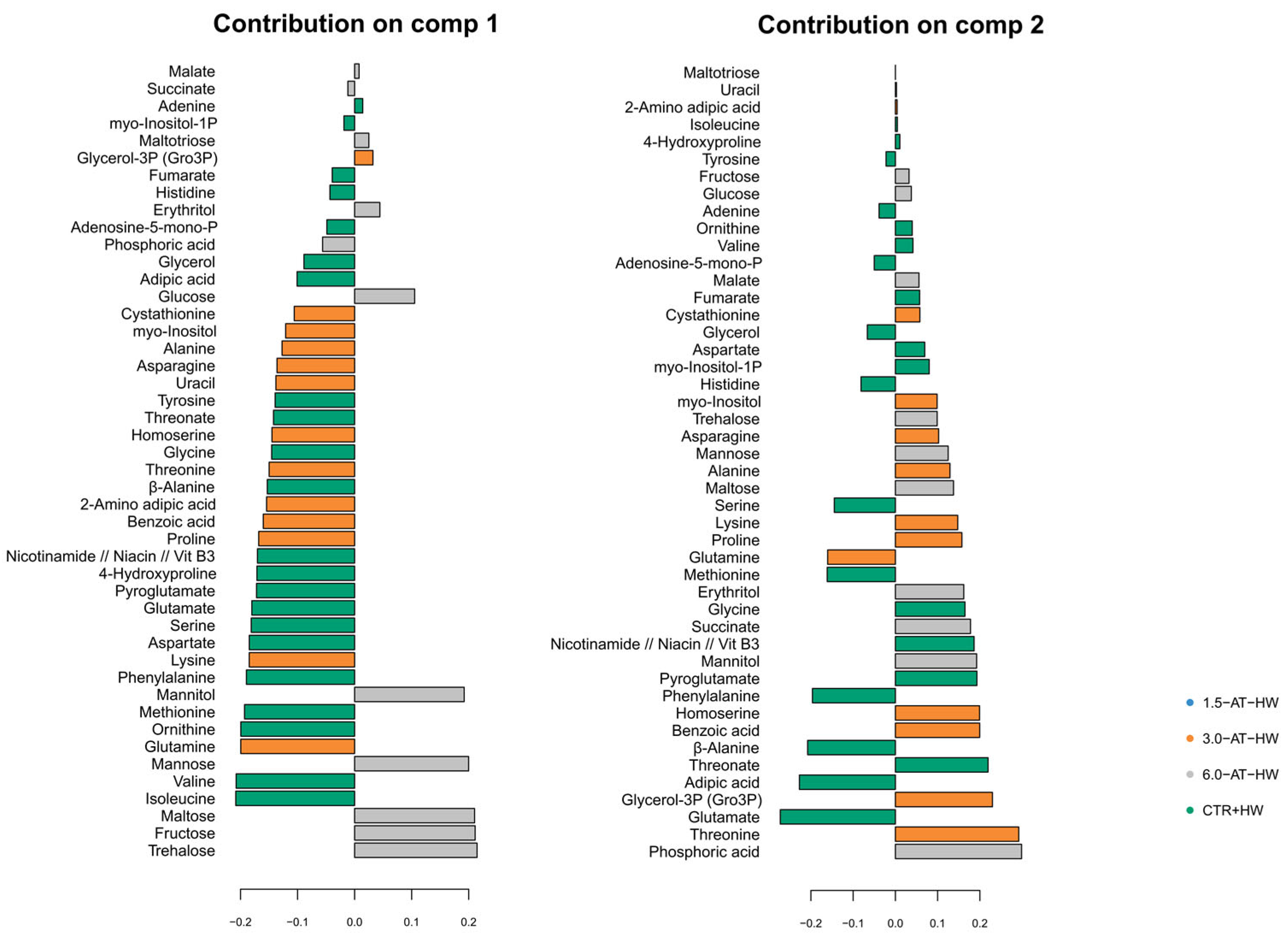
3.2. GC-TOF-MS Primary Metabolite Profiling of Diplodus Sargus Liver Tissues When Exposed to the MHW
4. Discussion
4.1. Effects of A. taxiformis Supplementation on Fish Liver Primary Metabolome Under an Optimal Temperature Regime (CTR Versus 1.5-AT, 3.0-AT, and 6.0-AT)
4.2. Effects of MHW on Fish Liver Primary Metabolome (CTR Versus CTR+HW)
4.3. Influence of A. taxiformis Supplementation on Fish Metabolic Responses to MHW (CTR+HW Versus 1.5-AT-HW, 3.0-AT-HW, and 6.0-AT-HW)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; Academic Press: London, UK; San Diego, CA, USA, 2022. [Google Scholar]
- Gephart, J.A.; Golden, C.D.; Asche, F.; Belton, B.; Brugere, C.; Froehlich, H.E.; Fry, J.P.; Halpern, B.S.; Hicks, C.C.; Jones, R.C.; et al. Scenarios for Global Aquaculture and Its Role in Human Nutrition. Rev. Fish. Sci. Aquac. 2021, 29, 122–138. [Google Scholar] [CrossRef]
- Der Poel, A.F.B.V.; Abdollahi, M.R.; Cheng, H.; Colovic, R.; Den Hartog, L.A.; Miladinovic, D.; Page, G.; Sijssens, K.; Smillie, J.F.; Thomas, M.; et al. Future Directions of Animal Feed Technology Research to Meet the Challenges of a Changing World. Anim. Feed Sci. Technol. 2020, 270, 114692. [Google Scholar] [CrossRef]
- Hardy, R.W.; Kaushik, S.J. (Eds.) Fish Nutrition, 4th ed.; Academic Press: London, UK; San Diego, CA, USA; Cambridge, MA, USA; Kidlington, UK, 2022; ISBN 978-0-12-819587-1. [Google Scholar]
- Jimenez-Lopez, C.; Pereira, A.G.; Lourenço-Lopes, C.; Garcia-Oliveira, P.; Cassani, L.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Main Bioactive Phenolic Compounds in Marine Algae and Their Mechanisms of Action Supporting Potential Health Benefits. Food Chem. 2021, 341, 128262. [Google Scholar] [CrossRef]
- Naiel, M.A.E.; Alagawany, M.; Patra, A.K.; El-Kholy, A.I.; Amer, M.S.; Abd El-Hack, M.E. Beneficial Impacts and Health Benefits of Macroalgae Phenolic Molecules on Fish Production. Aquaculture 2021, 534, 736186. [Google Scholar] [CrossRef]
- Stadtlander, T.; Khalil, W.K.B.; Focken, U.; Becker, K. Effects of Low and Medium Levels of Red Alga Nori (Porphyra yezoensis Ueda) in the Diets on Growth, Feed Utilization and Metabolism in Intensively Fed Nile Tilapia, Oreochromis niloticus (L.). Aquacult. Nutr. 2013, 19, 64–73. [Google Scholar] [CrossRef]
- Cian, R.E.; Bacchetta, C.; Rossi, A.; Cazenave, J.; Drago, S.R. Red Seaweed Pyropia columbina as Antioxidant Supplement in Feed for Cultured Juvenile Pacú (Piaractus mesopotamicus). J. Appl. Phycol. 2019, 31, 1455–1465. [Google Scholar] [CrossRef]
- Xuan, X.; Li, W.; Zhu, W.; Wang, S. Effects of Different Levels of Macroalga Gracilaria lemaneiformis on Growth Performance and Feed Utilization on the Red Sea Bream, Pagrosomus major. J. Appl. Phycol. 2019, 31, 3213–3222. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Svendsen, J.C.; Malte, H.; Pereira, L.F.; Carvalho, P.; Pereira, R.; Gonçalves, J.F.M.; Ozório, R.O.A. Diets Supplemented with Seaweed Affect Metabolic Rate, Innate Immune, and Antioxidant Responses, but Not Individual Growth Rate in European Seabass (Dicentrarchus labrax). J. Appl. Phycol. 2016, 28, 2061–2071. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Salas-Leitón, E.; Brito, F.; Pereira, L.F.; Svendsen, J.C.; Baptista, T.; Pereira, R.; Abreu, H.; Reis, P.A.; Gonçalves, J.F.M.; et al. Effects of Dietary Gracilaria sp. and Alaria sp. Supplementation on Growth Performance, Metabolic Rates and Health in Meagre (Argyrosomus regius) Subjected to Pathogen Infection. J. Appl. Phycol. 2017, 29, 433–447. [Google Scholar] [CrossRef]
- Marmelo, I.; Dias, M.; Grade, A.; Pousão-Ferreira, P.; Diniz, M.S.; Marques, A.; Maulvault, A.L. Immunomodulatory and Antioxidant Effects of Functional Aquafeeds Biofortified with Whole Laminaria digitata in Juvenile Gilthead Seabream (Sparus aurata). Front. Mar. Sci. 2024, 11, 1325244. [Google Scholar] [CrossRef]
- Pereira, V.; Marques, A.; Gaivão, I.; Rego, A.; Abreu, H.; Pereira, R.; Santos, M.A.; Guilherme, S.; Pacheco, M. Marine Macroalgae as a Dietary Source of Genoprotection in Gilthead Seabream (Sparus aurata) against Endogenous and Exogenous Challenges. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 219, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Bansemir, A.; Blume, M.; Schröder, S.; Lindequist, U. Screening of Cultivated Seaweeds for Antibacterial Activity against Fish Pathogenic Bacteria. Aquaculture 2006, 252, 79–84. [Google Scholar] [CrossRef]
- Vatsos, I.N.; Rebours, C. Seaweed Extracts as Antimicrobial Agents in Aquaculture. J. Appl. Phycol. 2015, 27, 2017–2035. [Google Scholar] [CrossRef]
- Lozano, I.; Wacyk, J.M.; Carrasco, J.; Cortez-San Martín, M.A. Red Macroalgae Pyropia columbina and Gracilaria chilensis: Sustainable Feed Additive in the Salmo salar Diet and the Evaluation of Potential Antiviral Activity against Infectious Salmon Anemia Virus. J. Appl. Phycol. 2016, 28, 1343–1351. [Google Scholar] [CrossRef]
- Gora, A.H.; Sahu, N.P.; Sahoo, S.; Rehman, S.; Dar, S.A.; Ahmad, I.; Agarwal, D. Effect of Dietary Sargassum wightii and Its Fucoidan-Rich Extract on Growth, Immunity, Disease Resistance and Antimicrobial Peptide Gene Expression in Labeo rohita. Int. Aquat. Res. 2018, 10, 115–131. [Google Scholar] [CrossRef]
- Shi, Q.; Rong, H.; Hao, M.; Zhu, D.; Aweya, J.J.; Li, S.; Wen, X. Effects of Dietary Sargassum horneri on Growth Performance, Serum Biochemical Parameters, Hepatic Antioxidant Status, and Immune Responses of Juvenile Black Sea Bream Acanthopagrus schlegelii. J. Appl. Phycol. 2019, 31, 2103–2113. [Google Scholar] [CrossRef]
- Xuan, X.; Wen, X.; Li, S.; Zhu, D.; Li, Y. Potential Use of Macro-Algae Gracilaria lemaneiformis in Diets for the Black Sea Bream, Acanthopagrus schlegelii, Juvenile. Aquaculture 2013, 412–413, 167–172. [Google Scholar] [CrossRef]
- Kamunde, C.; Sappal, R.; Melegy, T.M. Brown Seaweed (AquaArom) Supplementation Increases Food Intake and Improves Growth, Antioxidant Status and Resistance to Temperature Stress in Atlantic Salmon, Salmo salar. PLoS ONE 2019, 14, e0219792. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A Hierarchical Approach to Defining Marine Heatwaves. Prog. Oceanogr. 2016, 141, 227–238. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Medrano, A.; Cerrano, C.; Ponti, M.; Schlegel, R.; Bensoussan, N.; Turicchia, E.; Sini, M.; Gerovasileiou, V.; et al. Marine Heatwaves Drive Recurrent Mass Mortalities in the Mediterranean Sea. Glob. Change Biol. 2022, 28, 5708–5725. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.; Leonelli, F.E.; García-Ladona, E.; Garrabou, J.; Kersting, D.K.; Bensoussan, N.; Pisano, A. Evolution of Marine Heatwaves in Warming Seas: The Mediterranean Sea Case Study. Front. Mar. Sci. 2023, 10, 1193164. [Google Scholar] [CrossRef]
- Islam, M.J.; Kunzmann, A.; Slater, M.J. Responses of Aquaculture Fish to Climate Change-induced Extreme Temperatures: A Review. J. World Aquac. Soc. 2022, 53, 314–366. [Google Scholar] [CrossRef]
- Barbosa, V.; Maulvault, A.L.; Anacleto, P.; Santos, M.; Mai, M.; Oliveira, H.; Delgado, I.; Coelho, I.; Barata, M.; Araújo-Luna, R.; et al. Enriched Feeds with Iodine and Selenium from Natural and Sustainable Sources to Modulate Farmed Gilthead Seabream (Sparus aurata) and Common Carp (Cyprinus carpio) Fillets Elemental Nutritional Value. Food Chem. Toxicol. 2020, 140, 111330. [Google Scholar] [CrossRef]
- Silva-Brito, F.; Timóteo, F.; Esteves, Â.; Peixoto, M.J.; Ozorio, R.; Magnoni, L. Impact of the Replacement of Dietary Fish Oil by Animal Fats and Environmental Salinity on the Metabolic Response of European Seabass (Dicentrarchus labrax). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 233, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Oliva-Teles, A. Nutrition and Health of Aquaculture Fish. J. Fish Dis. 2012, 35, 83–108. [Google Scholar] [CrossRef]
- Hixson, S.H. Fish Nutrition and Current Issues in Aquaculture: The Balance in Providing Safe and Nutritious Seafood, in an Environmentally Sustainable Manner. J. Aquac. Res. Development 2014, 3, 234. [Google Scholar] [CrossRef]
- Jobling, M. Fish Nutrition Research: Past, Present and Future. Aquacult. Int. 2016, 24, 767–786. [Google Scholar] [CrossRef]
- Mahanty, A.; Yadav, R.P.; Purohit, G.K.; Mohanty, S.; Mohanty, B.P. Metabolomic Response to High Temperature Stress in Murrel Channa striatus and Insights for Designer Feeds. In Outlook of Climate Change and Fish Nutrition; Sinha, A., Kumar, S., Kumari, K., Eds.; Springer Nature: Singapore, 2022; pp. 197–205. ISBN 978-981-19-5499-3. [Google Scholar]
- Viant, M.R.; Rosenblum, E.S.; Tjeerdema, R.S. NMR-Based Metabolomics: A Powerful Approach for Characterizing the Effects of Environmental Stressors on Organism Health. Environ. Sci. Technol. 2003, 37, 4982–4989. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Young, T. Showcasing Metabolomic Applications in Aquaculture: A Review. Rev. Aquac. 2018, 10, 135–152. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The Link between Genotypes and Phenotypes. In Functional Genomics; Town, C., Ed.; Springer: Dordrecht, The Netherlands, 2002; pp. 155–171. ISBN 978-94-010-3903-1. [Google Scholar]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite Profiling: From Diagnostics to Systems Biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef]
- Elolimy, A.; Alharthi, A.; Zeineldin, M.; Parys, C.; Helmbrecht, A.; Loor, J.J. Supply of Methionine During Late-Pregnancy Alters Fecal Microbiota and Metabolome in Neonatal Dairy Calves Without Changes in Daily Feed Intake. Front. Microbiol. 2019, 10, 2159. [Google Scholar] [CrossRef] [PubMed]
- Novais, F.J.; Pires, P.R.L.; Alexandre, P.A.; Dromms, R.A.; Iglesias, A.H.; Ferraz, J.B.S.; Styczynski, M.P.-W.; Fukumasu, H. Identification of a Metabolomic Signature Associated with Feed Efficiency in Beef Cattle. BMC Genomics 2019, 20, 8. [Google Scholar] [CrossRef]
- Carmelo, V.A.O.; Banerjee, P.; Da Silva Diniz, W.J.; Kadarmideen, H.N. Metabolomic Networks and Pathways Associated with Feed Efficiency and Related-Traits in Duroc and Landrace Pigs. Sci. Rep. 2020, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, B.A.; Powers, J.B.; Campagna, S.R.; Seay, T.B.; Embree, M.M.; Myer, P.R. Rumen Fluid Metabolomics of Beef Steers Differing in Feed Efficiency. Metabolomics 2020, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Chen, J.; Zhang, L.; Wang, X.; Yang, Y.; Ren, X. LC/MS-Based Metabolomics to Evaluate the Milk Composition of Human, Horse, Goat and Cow from China. Eur. Food Res. Technol. 2021, 247, 663–675. [Google Scholar] [CrossRef]
- Ye, X.; Zhou, L.; Zhang, Y.; Xue, S.; Gan, Q.F.; Fang, S. Effect of Host Breeds on Gut Microbiome and Serum Metabolome in Meat Rabbits. BMC Vet. Res. 2021, 17, 24. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography–Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.4.1–30.4.32. [Google Scholar] [CrossRef]
- Marmelo, I.; Lourenço-Marques, C.; Silva, I.A.L.; Soares, F.; Pousão-Ferreira, P.; Mata, L.; Marques, A.; Diniz, M.S.; Maulvault, A.L. Eco-Innovative Aquafeeds Biofortified with Asparagopsis taxiformis to Improve the Resilience of Farmed White Seabream (Diplodus sargus) to Marine Heatwave Events. Heliyon 2024, 10, e35135. [Google Scholar] [CrossRef]
- Belghit, I.; Rasinger, J.D.; Heesch, S.; Biancarosa, I.; Liland, N.; Torstensen, B.; Waagbø, R.; Lock, E.-J.; Bruckner, C.G. In-Depth Metabolic Profiling of Marine Macroalgae Confirms Strong Biochemical Differences between Brown, Red and Green Algae. Algal Res. 2017, 26, 240–249. [Google Scholar] [CrossRef]
- Mancuso, F.P.; D’Agostaro, R.; Milazzo, M.; Badalamenti, F.; Musco, L.; Mikac, B.; Lo Brutto, S.; Chemello, R. The Invasive Seaweed Asparagopsis taxiformis Erodes the Habitat Structure and Biodiversity of Native Algal Forests in the Mediterranean Sea. Mar. Environ. Res. 2022, 173, 105515. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, C.; Dias, M.; Narciso, L.; Mendonça, V. Reliance of Coastal Intertidal Food Webs on River Input—Current and Future Perspectives. Ecol. Indic. 2019, 101, 632–639. [Google Scholar] [CrossRef]
- Gamito, S.; Quental-Ferreira, H.; Parejo, A.; Aubin, J.; Christensen, V.; Cunha, M. Integrated Multi-Trophic Aquaculture Systems: Energy Transfers and Food Web Organization in Coastal Earthen Ponds. Aquacult. Environ. Interact. 2020, 12, 457–470. [Google Scholar] [CrossRef]
- Morales-Nin, B.; Moranta, J.; García, C.; Tugores, M.P.; Grau, A.M.; Riera, F.; Cerdà, M. The Recreational Fishery off Majorca Island (Western Mediterranean): Some Implications for Coastal Resource Management. ICES J. Mar. Sci. 2005, 62, 727–739. [Google Scholar] [CrossRef]
- Veiga, P.; Ribeiro, J.; Gonçalves, J.M.S.; Erzini, K. Quantifying Recreational Shore Angling Catch and Harvest in Southern Portugal (North-east Atlantic Ocean): Implications for Conservation and Integrated Fisheries Management. J. Fish Biol. 2010, 76, 2216–2237. [Google Scholar] [CrossRef]
- FAO. Fisheries and Aquaculture Department, FAO FishFinder Aquatic Species—Fact Sheets, Diplodus sargus; FAO: Rome, Italy, 2020. [Google Scholar]
- Santos, C.; Soares, F.; Candeias-Mendes, A.; Pousão-Ferreira, P.; Dinis, M.T.; Cortes Valente Oliveira, C. Characterization of Spawning Rhythms of a Sparidae Aquaculture Species, the White Seabream (Diplodus sargus), in the South of Portugal. Aquac. Res. 2022, 53, 1424–1434. [Google Scholar] [CrossRef]
- Pereira, A.; Marmelo, I.; Dias, M.; Silva, A.C.; Grade, A.C.; Barata, M.; Pousão-Ferreira, P.; Dias, J.; Anacleto, P.; Marques, A.; et al. Asparagopsis taxiformis as a Novel Antioxidant Ingredient for Climate-Smart Aquaculture: Antioxidant, Metabolic and Digestive Modulation in Juvenile White Seabream (Diplodus sargus) Exposed to a Marine Heatwave. Antioxidants 2024, 13, 949. [Google Scholar] [CrossRef]
- Hobday, A.J.; Oliver, E.C.; Gupta, A.S.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Holbrook, N.J.; Moore, P.J.; Thomsen, M.S.; Wernberg, T.; et al. Categorizing and Naming Marine Heatwaves. Oceanography 2018, 31, 162–173. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas Chromatography Mass Spectrometry–Based Metabolite Profiling in Plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Luedemann, A.; Strassburg, K.; Erban, A.; Kopka, J. TagFinder for the Quantitative Analysis of Gas Chromatography—Mass Spectrometry (GC-MS)-Based Metabolite Profiling Experiments. Bioinformatics 2008, 24, 732–737. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmuller, E.; Dormann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: The Golm Metabolome Database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef]
- Schauer, N.; Steinhauser, D.; Strelkov, S.; Schomburg, D.; Allison, G.; Moritz, T.; Lundgren, K.; Roessner-Tunali, U.; Forbes, M.G.; Willmitzer, L.; et al. GC–MS Libraries for the Rapid Identification of Metabolites in Complex Biological Samples. FEBS Letters 2005, 579, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio: Integrated Development for R. RStudio, PBC, Boston, MA, USA. 2020. Available online: http://www.rstudio.com/ (accessed on 8 July 2024).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, Version 1.3-3; R Package: Madison, WI, USA, 2020. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 2.12.1. 2020. Available online: http://CRAN.R-project.org/package=gplots (accessed on 13 March 2025).
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Machado, M.; Machado, S.; Pimentel, F.B.; Freitas, V.; Alves, R.C.; Oliveira, M.B.P.P. Amino Acid Profile and Protein Quality Assessment of Macroalgae Produced in an Integrated Multi-Trophic Aquaculture System. Foods 2020, 9, 1382. [Google Scholar] [CrossRef] [PubMed]
- Félix, R.; Dias, P.; Félix, C.; Cerqueira, T.; Andrade, P.B.; Valentão, P.; Lemos, M.F.L. The Biotechnological Potential of Asparagopsis armata: What Is Known of Its Chemical Composition, Bioactivities and Current Market? Algal Res. 2021, 60, 102534. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Hayes, M. In Vitro Protein Digestibility of Selected Seaweeds. Foods 2022, 11, 289. [Google Scholar] [CrossRef]
- Zhou, A.Y.; Robertson, J.; Hamid, N.; Ma, Q.; Lu, J. Changes in Total Nitrogen and Amino Acid Composition of New Zealand Undaria pinnatifida with Growth, Location and Plant Parts. Food Chem. 2015, 186, 319–325. [Google Scholar] [CrossRef]
- Aksnes, A.; Mundheim, H.; Toppe, J.; Albrektsen, S. The Effect of Dietary Hydroxyproline Supplementation on Salmon (Salmo salar L.) Fed High Plant Protein Diets. Aquaculture 2008, 275, 242–249. [Google Scholar] [CrossRef]
- Liu, X.; Mai, K.; Liufu, Z.; Ai, Q. Effects of Dietary Protein and Lipid Levels on Growth, Nutrient Utilization, and the Whole-body Composition of Turbot, Scophthalmus maximus, Linnaeus 1758, at Different Growth Stages. J. World Aquac. Soc. 2014, 45, 355–366. [Google Scholar] [CrossRef]
- Rong, H.; Zhang, Y.; Hao, M.; Lin, F.; Zou, W.; Zhang, H.; Yu, C.; Yu, J.; Shi, Q.; Aweya, J.J.; et al. Effect of Hydroxyproline Supplementation on Growth Performance, Body Composition, Amino Acid Profiles, Blood-biochemistry and Collagen Synthesis of Juvenile Chu’s Croaker (Nibea coibor). Aquac. Res. 2020, 51, 1264–1275. [Google Scholar] [CrossRef]
- Salamanca, N.; Giráldez, I.; Morales, E.; De La Rosa, I.; Herrera, M. Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre. Animals 2020, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hou, Y.; Dai, Z.; Hu, C.-A.A.; Wu, G. Metabolism, Nutrition, and Redox Signaling of Hydroxyproline. Antioxid. Redox Signal. 2019, 30, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhao, L.; Ma, R.; Wang, J.; Du, L. FoxO Signaling and Mitochondria-Related Apoptosis Pathways Mediate Tsinling Lenok Trout (Brachymystax lenok tsinlingensis) Liver Injury under High Temperature Stress. Int. J. Biol. Macromol. 2023, 251, 126404. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New Developments in Fish Amino Acid Nutrition: Towards Functional and Environmentally Oriented Aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Jasour, M.S.; Wagner, L.; Sundekilde, U.K.; Larsen, B.K.; Greco, I.; Orlien, V.; Olsen, K.; Rasmussen, H.T.; Hjermitslev, N.H.; Hammershøj, M.; et al. A Comprehensive Approach to Assess Feathermeal as an Alternative Protein Source in Aquafeed. J. Agric. Food Chem. 2017, 65, 10673–10684. [Google Scholar] [CrossRef] [PubMed]
- Phang, J.M.; Donald, S.P.; Pandhare, J.; Liu, Y. The Metabolism of Proline, a Stress Substrate, Modulates Carcinogenic Pathways. Amino Acids 2008, 35, 681–690. [Google Scholar] [CrossRef]
- Hu, S.; He, W.; Wu, G. Hydroxyproline in Animal Metabolism, Nutrition, and Cell Signaling. Amino Acids 2022, 54, 513–528. [Google Scholar] [CrossRef]
- Nisar, N.; Mustafa, F.; Tahir, A.; Qadri, R.; Yang, Y.; Khan, M.I.; Wang, F. Proximate Composition, Functional Properties and Quantitative Analysis of Benzoyl Peroxide and Benzoic Acid in Wheat Flour Samples: Effect on Wheat Flour Quality. PeerJ 2020, 8, e8788. [Google Scholar] [CrossRef]
- Libanori, M.C.M.; Santos, G.G.; Pereira, S.A.; Lopes, G.R.; Owatari, M.S.; Soligo, T.A.; Yamashita, E.; Pereira, U.P.; Martins, M.L.; Mouriño, J.L.P. Dietary Supplementation with Benzoic Organic Acid Improves the Growth Performance and Survival of Nile Tilapia (Oreochromis niloticus) after Challenge with Streptococcus agalactiae (Group B). Aquaculture 2021, 545, 737204. [Google Scholar] [CrossRef]
- Hussein, E.E.; Habiba, M.M.; Ashry, A.M.; Al-Zayat, A.M.; Teiba, I.I.; Shehata, A.I.; Shahin, S.A.; El-Ratel, I.T.; Mzengereza, K.; Tembo, M.; et al. Effects of Dietary Supplementation with Organic Acids Mixture on Growth, Feed Efficiency, Hematobiochemical Parameters, Immunity, and Intestinal Microbiota of Gilthead Seabream (Sparus aurata) Juveniles. Aquac. Rep. 2023, 33, 101846. [Google Scholar] [CrossRef]
- Santos, G.G.; Libanori, M.C.M.; Pereira, S.A.; Ferrarezi, J.V.S.; Ferreira, M.B.; Soligo, T.A.; Yamashita, E.; Martins, M.L.; Mouriño, J.L.P. Probiotic Mix of Bacillus spp. and Benzoic Organic Acid as Growth Promoter against Streptococcus agalactiae in Nile Tilapia. Aquaculture 2023, 566, 739212. [Google Scholar] [CrossRef]
- Hu, Z.; Zhu, Y.; Chen, J.; Chen, J.; Li, C.; Gao, Z.; Li, J.; Liu, L. Discovery of Novel Bactericides from Aspergillus alabamensis and Their Antibacterial Activity against Fish Pathogens. J. Agric. Food Chem. 2023, 71, 4298–4305. [Google Scholar] [CrossRef]
- Long, Z.; Qin, H.; Huang, Z.; Xu, A.; Ye, Y.; Li, Z. Effects of Heat Stress on Physiological Parameters, Biochemical Parameters and Expression of Heat Stress Protein Gene in Lateolabrax maculatus. J. Therm. Biol. 2023, 115, 103606. [Google Scholar] [CrossRef]
- Hao, R.; Li, H.; Tian, Y.; Ru, X.; Deng, Q.; Zhu, K.; Yang, T.; Huang, Y.; Zhu, C. The Effect of Heat Stress on Energy Metabolism, Immune Function, and Oxidative Stress of Juvenile Greater Amberjack Seriola dumerili. Aquac. Res. 2024, 2024, 1–9. [Google Scholar] [CrossRef]
- Yan, H.; Du, J.; Li, S.; Lei, C.; Zhu, T.; Han, L.; Song, H. Chronic Heat Stress Is Capable of Reducing the Growth Performance, Causing Damage to the Liver Structure, and Altering the Liver Glucose Metabolism and Lipid Metabolism in Largemouth Bass (Micropterus salmoides L.). Fish Physiol. Biochem. 2025, 51, 24. [Google Scholar] [CrossRef]
- Currie, S.; Bagatto, B.; DeMille, M.; Learner, A.; LeBlanc, D.; Marks, C.; Ong, K.; Parker, J.; Templeman, N.; Tufts, B.L.; et al. Metabolism, Nitrogen Excretion, and Heat Shock Proteins in the Central Mudminnow (Umbra limi), a Facultative Air-Breathing Fish Living in a Variable Environment. Can. J. Zool. 2010, 88, 43–58. [Google Scholar] [CrossRef]
- Song, M.; Zhao, J.; Wen, H.-S.; Li, Y.; Li, J.-F.; Li, L.-M.; Tao, Y.-X. The Impact of Acute Thermal Stress on the Metabolome of the Black Rockfish (Sebastes schlegelii). PLoS ONE 2019, 14, e0217133. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Ye, S.; Bureau, D.P.; Liu, H.; Yin, J.; Mou, Z.; Lin, H.; Hao, F. Global Metabolic Responses of the Lenok (Brachymystax lenok) to Thermal Stress. Comp. Biochem. Physiol. Part D Genomics Proteomics 2019, 29, 308–319. [Google Scholar] [CrossRef]
- Hillyer, K.E.; Beale, D.J.; Shima, J.S. Artificial Light at Night Interacts with Predatory Threat to Alter Reef Fish Metabolite Profiles. Sci. Total Environ. 2021, 769, 144482. [Google Scholar] [CrossRef]
- Clark, T.C.; Tinsley, J.; Macqueen, D.J.; Martin, S.A.M. Rainbow Trout (Oncorhynchus mykiss) Urea Cycle and Polyamine Synthesis Gene Families Show Dynamic Expression Responses to Inflammation. Fish Shellfish Immunol. 2019, 89, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Engelking, L.R. Chapter 10: Urea Cycle (Krebs-Henseleit Ornithine Cycle). In Textbook of Veterinary Physiological Chemistry; Academic Press: Cambridge, MA, USA, 2015; pp. 58–64. ISBN 978-0-12-391909-0. [Google Scholar]
- Jiang, Y.; Cheng, X.; Lu, J.; Xu, G.; Liu, Q.; Sun, J. Thermal Stress Induces Metabolic Responses in Juvenile Qingtian Paddy Field Carp Cyprinus carpio var qingtianensis. Animals 2022, 12, 3395. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Huang, F.; Zhang, Z.; Habumugisha, T.; Yan, C.; Shaheen, U.; Zhang, X. Nanoplastic Contamination: Impact on Zebrafish Liver Metabolism and Implications for Aquatic Environmental Health. Environ. Int. 2024, 187, 108713. [Google Scholar] [CrossRef]
- Gamarra, Y.; Santiago, F.C.; Molina-López, J.; Castaño, J.; Herrera-Quintana, L.; Domínguez, Á.; Planells, E. Pyroglutamic Acidosis by Glutathione Regeneration Blockage in Critical Patients with Septic Shock. Crit. Care 2019, 23, 162. [Google Scholar] [CrossRef]
- Fang, Y.-Z.; Yang, S.; Wu, G. Free Radicals, Antioxidants, and Nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Aguilar, A.; Mattos, H.; Carnicero, B.; Sanhueza, N.; Muñoz, D.; Teles, M.; Tort, L.; Boltaña, S. Metabolomic Profiling Reveals Changes in Amino Acid and Energy Metabolism Pathways in Liver, Intestine and Brain of Zebrafish Exposed to Different Thermal Conditions. Front. Mar. Sci. 2022, 9, 835379. [Google Scholar] [CrossRef]
- Yang, D.-X.; Yang, M.-J.; Yin, Y.; Kou, T.-S.; Peng, L.-T.; Chen, Z.-G.; Zheng, J.; Peng, B. Serine Metabolism Tunes Immune Responses To Promote Oreochromis niloticus Survival upon Edwardsiella tarda Infection. mSystems 2021, 6, e00426-21. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, A.; Ma, S.; Lee, W.; Lee, S.; Yoon, D.; Kim, D.-H.; Kim, S. Glutathione Injection Alleviates the Fluctuation of Metabolic Response under Thermal Stress in Olive Flounder, Paralichthys olivaceus. Metabolites 2019, 10, 3. [Google Scholar] [CrossRef]
- Adam, A.-C.; Lie, K.K.; Moren, M.; Skjærven, K.H. High Dietary Arachidonic Acid Levels Induce Changes in Complex Lipids and Immune-Related Eicosanoids and Increase Levels of Oxidised Metabolites in Zebrafish (Danio rerio). Br. J. Nutr. 2017, 117, 1075–1085. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, W.; Jiang, W.; Zhang, G.; He, J.; Dong, S.; Zhou, Y.; Yang, W.; Tang, Q.; Yu, Y.; et al. Non-Targeted Metabolomics Provides Insights into the Distinct Amino Acid and Lipid Metabolism in Liver Tissues of Rainbow Trout (Oncorhynchus mykiss) Cultured in Seawater at Different Temperatures. Aquaculture 2024, 579, 740188. [Google Scholar] [CrossRef]
- Falco, F.; Stincone, P.; Cammarata, M.; Brandelli, A. Amino Acids as the Main Energy Source in Fish Tissues. Aquac. Fish Stud. 2020, 3, 1–11. [Google Scholar] [CrossRef]
- Zhao, H.; Ke, H.; Zhang, L.; Zhao, Z.; Lai, J.; Zhou, J.; Huang, Z.; Li, H.; Du, J.; Li, Q. Integrated Analysis about the Effects of Heat Stress on Physiological Responses and Energy Metabolism in Gymnocypris chilianensis. Sci. Total Environ. 2022, 806, 151252. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Feng, B.; Zhu, W.; Liang, Z.; Xu, W.; Fu, J.; Miao, L.; Dong, Z. Proteomics and Metabolomics Analysis of American Shad (Alosa sapidissima) Liver Responses to Heat Stress. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2024, 296, 111686. [Google Scholar] [CrossRef]
- Kooijman, S.A.L.M.; Baas, J.; Bontje, D.; Broerse, M.; Van Gestel, C.A.M.; Jager, T. Ecotoxicological Applications of Dynamic Energy Budget Theory. In Ecotoxicology Modeling; Devillers, J., Ed.; Emerging Topics in Ecotoxicology; Springer: Boston, MA, USA, 2009; Volume 2, pp. 237–259. ISBN 978-1-4419-0196-5. [Google Scholar]
- Yan, L.; Qiu-Zhou, X. Dietary Glutamine Supplementation Improves Structure and Function of Intestine of Juvenile Jian Carp (Cyprinus carpio var. Jian). Aquaculture 2006, 256, 389–394. [Google Scholar] [CrossRef]
- Cheng, Z.; Buentello, A.; Gatlin, D.M. Effects of Dietary Arginine and Glutamine on Growth Performance, Immune Responses and Intestinal Structure of Red Drum, Sciaenops ocellatus. Aquaculture 2011, 319, 247–252. [Google Scholar] [CrossRef]
- Larsen, T.; Wang, Y.V.; Wan, A.H.L. Tracing the Trophic Fate of Aquafeed Macronutrients With Carbon Isotope Ratios of Amino Acids. Front. Mar. Sci. 2022, 9, 813961. [Google Scholar] [CrossRef]
- Zhou, X. Use of Synthetic Lysine in Fish Feeds: A Review on Research and Application. Feed Ind. 2005, 27, 1–7. [Google Scholar]
- Ramos-Pinto, L.; Machado, M.; Calduch-Giner, J.; Pérez-Sánchez, J.; Dias, J.; Conceição, L.E.C.; Silva, T.S.; Costas, B. Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals 2021, 11, 1193. [Google Scholar] [CrossRef]
- Wu, G. Functional Amino Acids in Nutrition and Health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef]
- Li, X.; Zheng, S.; Wu, G. Nutrition and Metabolism of Glutamate and Glutamine in Fish. Amino Acids 2020, 52, 671–691. [Google Scholar] [CrossRef]
- Gillis, T.E.; Ballantyne, J.S. The Effects of Starvation on Plasma Free Amino Acid and Glucose Concentrations in Lake Sturgeon. J. Fish Biol. 1996, 49, 1306–1316. [Google Scholar] [CrossRef]
- Hassel, B. Pyruvate Carboxylation in Neurons. J. Neurosci. Res. 2001, 66, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, C.; Wang, B.; Wang, C.; Sagada, G.; Yan, Y. Methionine in Fish Health and Nutrition: Potential Mechanisms, Affecting Factors, and Future Perspectives. Aquaculture 2023, 568, 739310. [Google Scholar] [CrossRef]
- Tulli, F.; Messina, M.; Calligaris, M.; Tibaldi, E. Response of European Sea Bass (Dicentrarchus labrax) to Graded Levels of Methionine (Total Sulfur Amino Acids) in Soya Protein-Based Semi-Purified Diets. Br. J. Nutr. 2010, 104, 664–673. [Google Scholar] [CrossRef]
- He, J.-Y.; Tian, L.-X.; Lemme, A.; Gao, W.; Yang, H.-J.; Niu, J.; Liang, G.-Y.; Chen, P.-F.; Liu, Y.-J. Methionine and Lysine Requirements for Maintenance and Efficiency of Utilization for Growth of Two Sizes of Tilapia (Oreochromis niloticus). Aquacult. Nutr. 2013, 19, 629–640. [Google Scholar] [CrossRef]
- Rolland, M.; Dalsgaard, J.; Holm, J.; Gómez-Requeni, P.; Skov, P.V. Dietary Methionine Level Affects Growth Performance and Hepatic Gene Expression of GH–IGF System and Protein Turnover Regulators in Rainbow Trout (Oncorhynchus mykiss) Fed Plant Protein-Based Diets. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2015, 181, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Belghit, I.; Skiba-Cassy, S.; Geurden, I.; Dias, K.; Surget, A.; Kaushik, S.; Panserat, S.; Seiliez, I. Dietary Methionine Availability Affects the Main Factors Involved in Muscle Protein Turnover in Rainbow Trout (Oncorhynchus mykiss). Br. J. Nutr. 2014, 112, 493–503. [Google Scholar] [CrossRef]
- Kuang, S.-Y.; Xiao, W.-W.; Feng, L.; Liu, Y.; Jiang, J.; Jiang, W.-D.; Hu, K.; Li, S.-H.; Tang, L.; Zhou, X.-Q. Effects of Graded Levels of Dietary Methionine Hydroxy Analogue on Immune Response and Antioxidant Status of Immune Organs in Juvenile Jian Carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2012, 32, 629–636. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary Tryptophan and Methionine as Modulators of European Seabass (Dicentrarchus labrax) Immune Status and Inflammatory Response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef]
- Pan, F.-Y.; Feng, L.; Jiang, W.-D.; Jiang, J.; Wu, P.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Zhou, X.-Q.; et al. Methionine Hydroxy Analogue Enhanced Fish Immunity via Modulation of NF-κB, TOR, MLCK, MAPKs and Nrf2 Signaling in Young Grass Carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2016, 56, 208–228. [Google Scholar] [CrossRef]
- Elmada, C.Z.; Huang, W.; Jin, M.; Liang, X.; Mai, K.; Zhou, Q. The Effect of Dietary Methionine on Growth, Antioxidant Capacity, Innate Immune Response and Disease Resistance of Juvenile Yellow Catfish (Pelteobagrus fulvidraco). Aquacult. Nutr. 2016, 22, 1163–1173. [Google Scholar] [CrossRef]
- Noor, Z.; Noor, M.; Khan, S.A.; Younas, W.; Ualiyeva, D.; Hassan, Z.; Yousafzai, A.M. Dietary Supplementations of Methionine Improve Growth Performances, Innate Immunity, Digestive Enzymes, and Antioxidant Activities of Rohu (Labeo rohita). Fish Physiol. Biochem. 2021, 47, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Yang, L.-F.; Zheng, J.; Chen, Z.-G.; Peng, B. Maltose Promotes Crucian Carp Survival against Aeromonas sobrial Infection at High Temperature. Virulence 2020, 11, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Kou, T.; Peng, L.; Munang’andu, H.M.; Peng, B. Fructose Promotes Crucian Carp Survival Against Aeromonas hydrophila Infection. Front. Immunol. 2022, 13, 865560. [Google Scholar] [CrossRef] [PubMed]
- Kou, T.; Wu, J.; Chen, X.; Peng, B. Functional Proteomics Identify Mannitol Metabolism in Serum Resistance and Therapeutic Implications in Vibrio alginolyticus. Front. Immunol. 2022, 13, 1010526. [Google Scholar] [CrossRef]
- Li, R.-X.; Chen, L.-Y.; Yao, B.; Rahimnejad, S.; Ren, J.; Luo, Y.; Qiao, F.; Zhang, M.-L.; Du, Z.-Y. Trehalose Alleviated Hepatic Cholesterol Accumulation via Inhibiting Transformation from Glucose-Derived Acyl-CoA to Cholesterol Synthesis in Nile Tilapia. Aquaculture 2022, 560, 738600. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | CTR | 1.5-AT | 3.0-AT | 6.0-AT |
|---|---|---|---|---|
| Fishmeal super prime 1 | 25.0 | 25.0 | 25.0 | 25.0 |
| Fish protein concentrate 2 | 2.0 | 2.0 | 2.0 | 2.0 |
| Soy protein concentrate 3 | 10.0 | 10.0 | 10.0 | 10.0 |
| Pea protein concentrate 4 | 3.0 | 3.0 | 3.0 | 3.0 |
| Wheat gluten 5 | 6.5 | 6.5 | 6.5 | 6.5 |
| Corn gluten meal 6 | 10.0 | 10.0 | 10.0 | 10.0 |
| Soybean meal 44 7 | 6.0 | 6.0 | 6.0 | 6.0 |
| Rapeseed meal 8 | 6.0 | 6.0 | 6.0 | 6.0 |
| Wheat meal 9 | 10.8 | 9.3 | 7.8 | 4.8 |
| Faba beans (low tannins) 10 | 6.0 | 6.0 | 6.0 | 6.0 |
| Vitamin and mineral premix 11 | 1.0 | 1.0 | 1.0 | 1.0 |
| Choline chloride 50% 12 | 0.2 | 0.2 | 0.2 | 0.2 |
| Monoammonium phosphate 13 | 1.2 | 1.2 | 1.2 | 1.2 |
| Fish oil 14 | 5.0 | 5.0 | 5.0 | 5.0 |
| Soybean oil 15 | 7.3 | 7.3 | 7.3 | 7.3 |
| Macroalga Asparagopsis taxiformis 16 | 0 | 1.5 | 3.0 | 6.0 |
| Dry matter, DM (%) | 94.2 | 94.0 | 93.9 | 94.1 |
| Crude protein, %DM | 46.0 | 46.0 | 45.9 | 45.7 |
| Crude fat, %DM | 16.0 | 16.0 | 16.1 | 16.1 |
| Fiber, %DM | 1.8 | 1.9 | 2.0 | 2.1 |
| Starch, %DM | 13.7 | 12.8 | 11.8 | 9.9 |
| Ash, %DM | 6.8 | 7.1 | 7.4 | 8.0 |
| Gross energy, MJ kg−1 | 21.0 | 21.0 | 20.9 | 20.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, M.; Marmelo, I.; António, C.; Rodrigues, A.M.; Marques, A.; Diniz, M.S.; Maulvault, A.L. Asparagopsis taxiformis Feed Supplementation as a Tool to Improve the Resilience of Farmed Diplodus sargus to Marine Heatwave Events—A Metabolomics Approach. Fishes 2025, 10, 350. https://doi.org/10.3390/fishes10070350
Dias M, Marmelo I, António C, Rodrigues AM, Marques A, Diniz MS, Maulvault AL. Asparagopsis taxiformis Feed Supplementation as a Tool to Improve the Resilience of Farmed Diplodus sargus to Marine Heatwave Events—A Metabolomics Approach. Fishes. 2025; 10(7):350. https://doi.org/10.3390/fishes10070350
Chicago/Turabian StyleDias, Marta, Isa Marmelo, Carla António, Ana M. Rodrigues, António Marques, Mário S. Diniz, and Ana Luísa Maulvault. 2025. "Asparagopsis taxiformis Feed Supplementation as a Tool to Improve the Resilience of Farmed Diplodus sargus to Marine Heatwave Events—A Metabolomics Approach" Fishes 10, no. 7: 350. https://doi.org/10.3390/fishes10070350
APA StyleDias, M., Marmelo, I., António, C., Rodrigues, A. M., Marques, A., Diniz, M. S., & Maulvault, A. L. (2025). Asparagopsis taxiformis Feed Supplementation as a Tool to Improve the Resilience of Farmed Diplodus sargus to Marine Heatwave Events—A Metabolomics Approach. Fishes, 10(7), 350. https://doi.org/10.3390/fishes10070350