
Limit Reference Points and Equilibrium Stock Dynamics in the Presence of Recruitment Depensation


Abstract
1. Introduction
2. Materials and Methods
2.1. Data
2.2. Stock–Recruitment Models
2.3. Traditional Reference Points
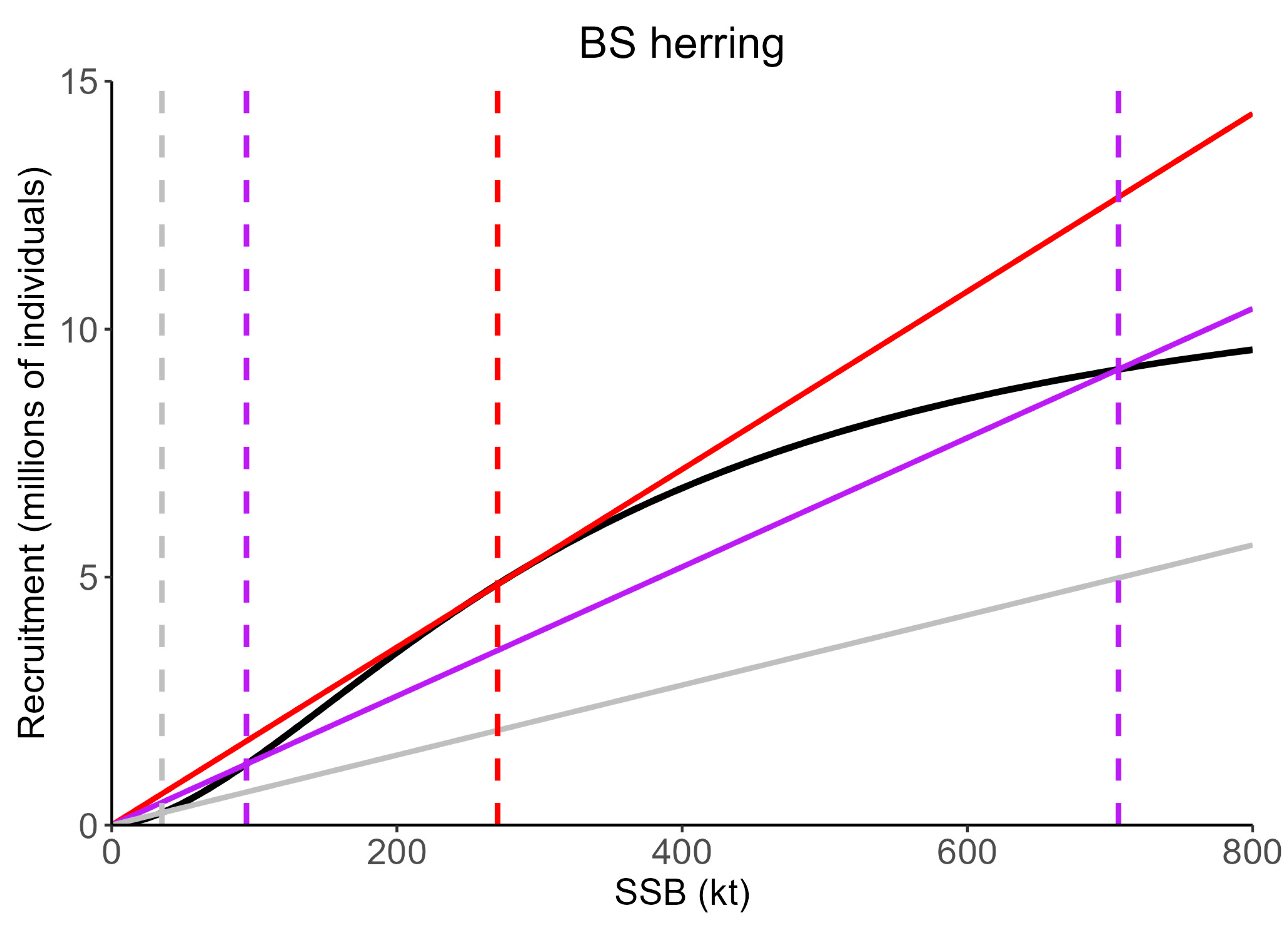
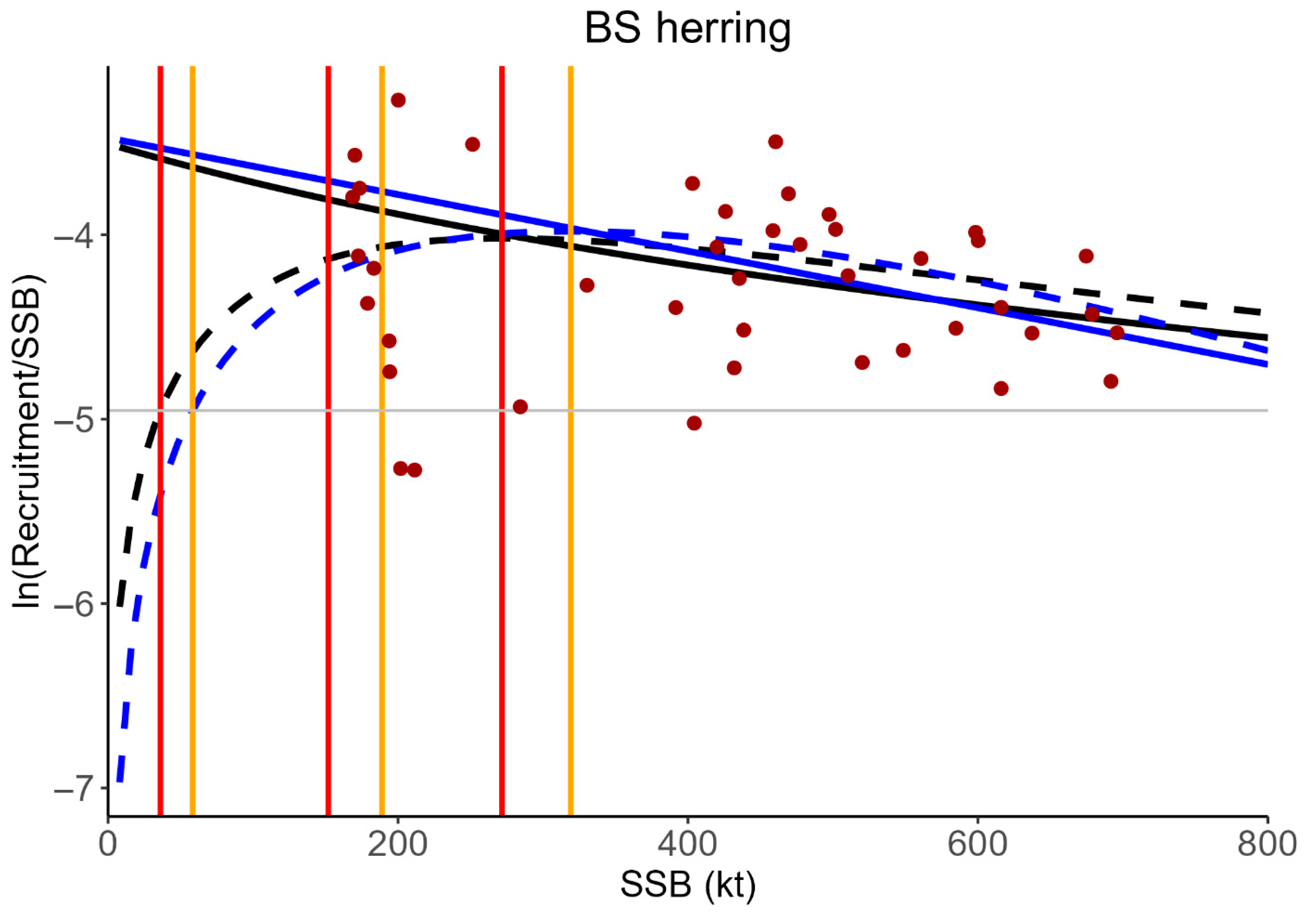
2.4. Allee Effect Reference Points
3. Results
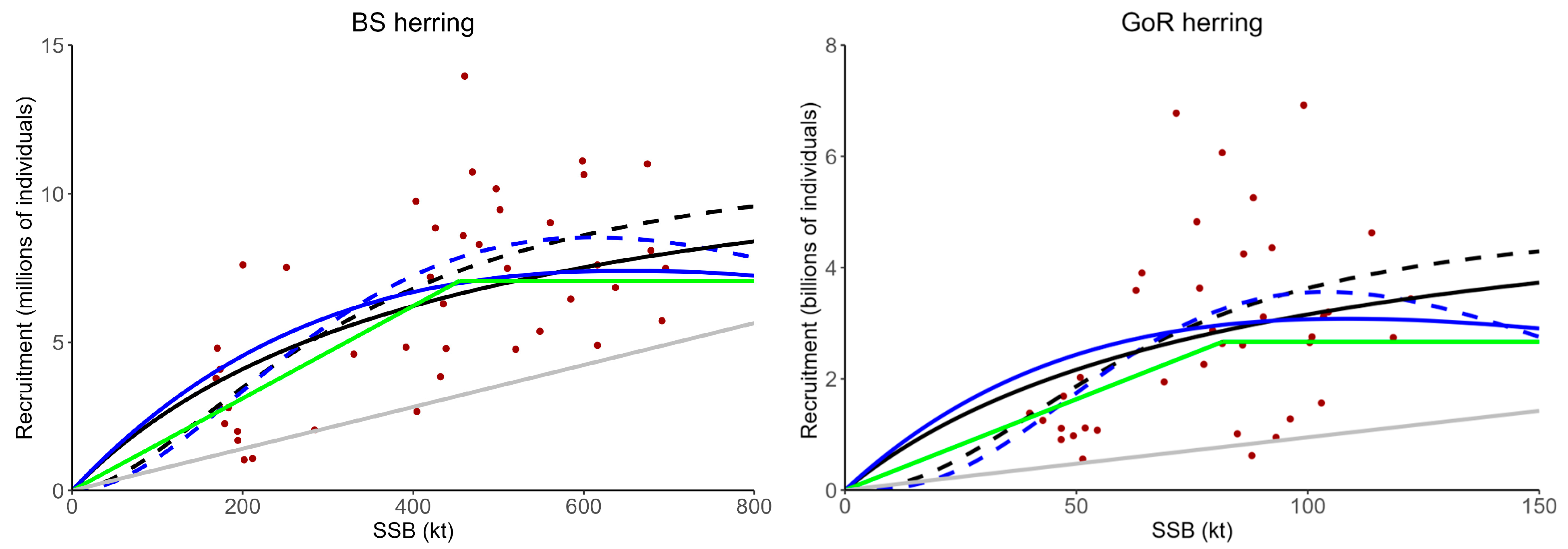
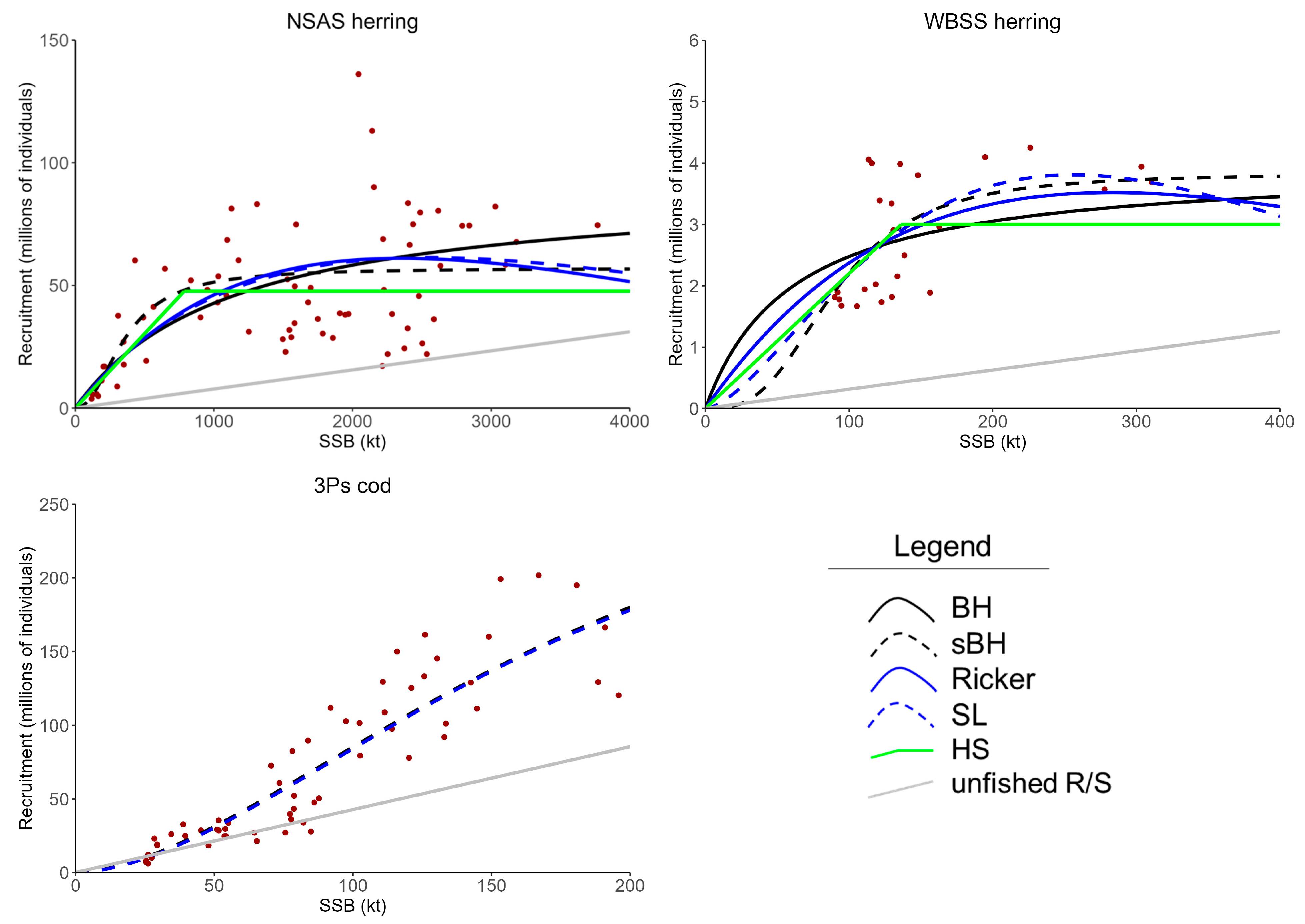
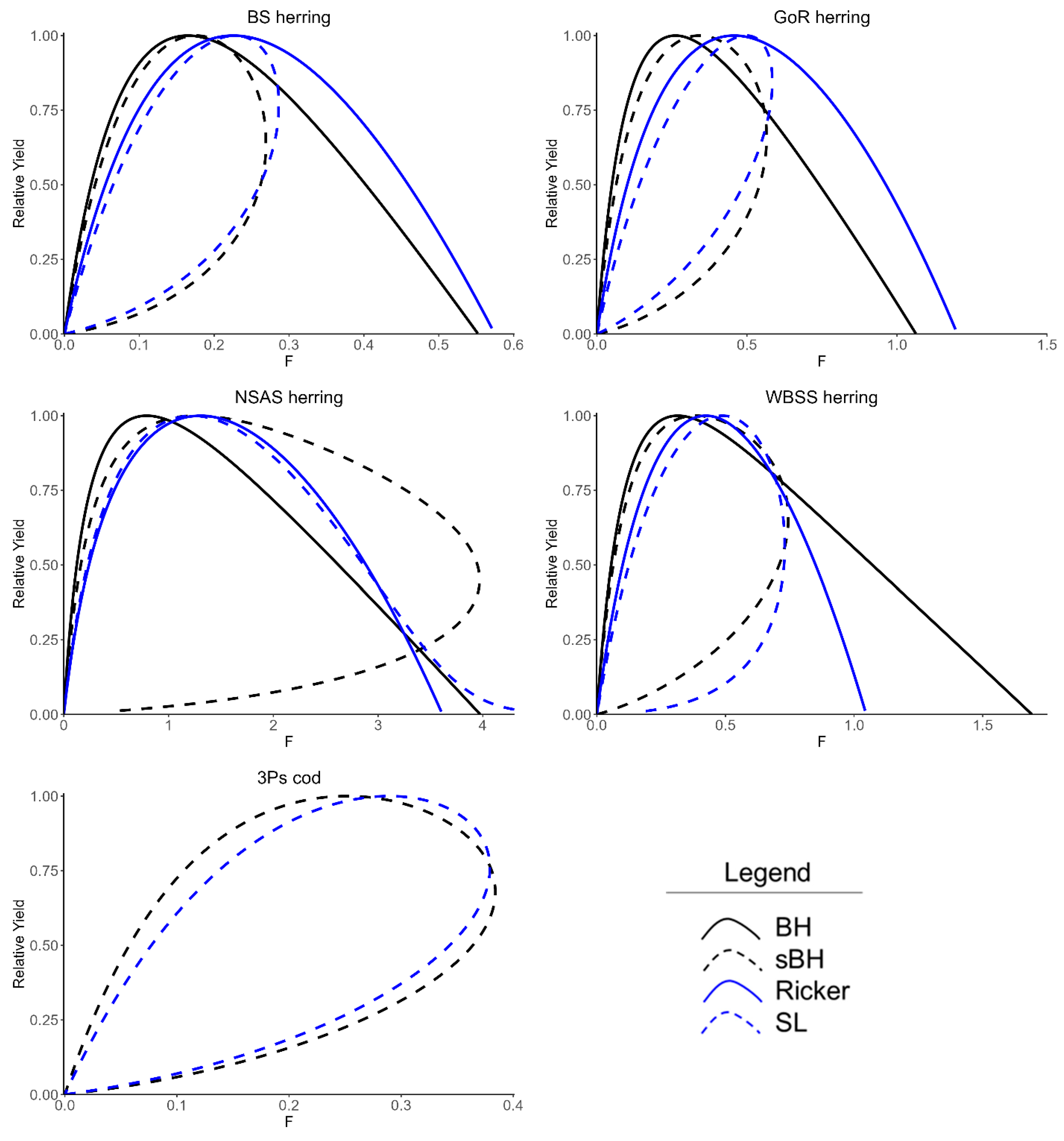
3.1. Model Fits and Comparison of Candidate LRPs
3.2. Stock Dynamics with Multiple Equilibrium States and F Reference Points
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Maximum age group (plus group) | |
| Age at recruitment | |
| AIC | Akaike information criterion |
| B | Biomass |
| B0 | Equilibrium unfished biomass |
| B50 | Model predicted biomass at 50% of asymptotic recruitment |
| BCP | Biomass at the change point of a hockey-stick stock–recruitment relationship |
| Bk | Biomass at maximum predicted recruitment |
| Binflection | Biomass at the inflection point of a stock–recruitment relationship |
| Blim | Biomass limit below which a stock is considered to have reduced reproductive capacity |
| BMSY | Equilibrium biomass at maximum sustainable yield |
| BH | Beverton–Holt |
| BS | Bothnian Sea |
| c | Depensation parameter |
| DFO | Fisheries and Oceans Canada |
| F | Fishing mortality rate |
| Fcrash | Long-term fishing mortality rate that would lead the stock to crash |
| FMSY | Fishing mortality rate at maximum sustainable yield |
| GoR | Gulf of Riga |
| HS | Hockey stick |
| ICES | International Council for the Exploration of the Sea |
| k | Maximum predicted recruitment |
| survivorship-at-age | |
| LOOIC | Leave-one-out cross-validation information criterion |
| LRP | Limit reference point |
| M | Natural mortality rate |
| Maturity-at-age | |
| Natural mortality rate-at-age | |
| NAFO | Northwest Atlantic Fisheries Organization |
| NSAS | North Sea autumn spawners |
| Spawning stock biomass-per-recruit | |
| R/S | Recruits-per-spawner |
| Asymptotic recruitment | |
| RAM | Ransom A. Myers |
| sBH | Sigmoid Beverton–Holt |
| SL | Saila–Lorda |
| SRR | Stock–recruitment relationship |
| SSB | Spawning stock biomass |
| USA | United States of America |
| Selectivity-at-age | |
| Weight-at-age | |
| WBSS | Western Baltic spawning spawners |
| YPR | Yield-per-recruit |
References
- DFO (Fisheries and Oceans Canada). A Fishery Decision-Making Framework Incorporating the Precautionary Approach. Last Updated 2009-03-23. 2009. Available online: https://www.dfo-mpo.gc.ca/reports-rapports/regs/sff-cpd/precaution-eng.htm (accessed on 20 May 2025).
- ICES (International Council for the Exploration of the Sea). Technical Guidelines. In Report of the ICES Advisory Committee, 2021. ICES Advice 2021, Section 16.4.3.1. 2021. Available online: https://doi.org/10.17895/ices.advice.7891 (accessed on 20 May 2025).
- eCFR (Electronic Code of Federal Regulations). Title 50: Wildlife and Fisheries, Part 600: Magnuson-Stevens Act Provisions, Subpart D: National Standards. 2021. Available online: https://www.ecfr.gov/cgi-bin/text-idx?SID=71b8c6026001cb90e4b0925328dce685&mc=true&node=se50.12.600_1310&rgn=div8 (accessed on 20 May 2025).
- DAWR (Department of Agriculture and Water Resources, Australian Government). Guidelines for the Implementation of the Commonwealth Fisheries Harvest Strategy Policy. 2nd Edition. 2018. Available online: https://www.agriculture.gov.au/sites/default/files/sitecollectiondocuments/fisheries/domestic/hsp.pdf (accessed on 20 May 2025).
- Sainsbury, K. Best Practice Reference Points for Australian Fisheries. Australian Fisheries Management Authority Report R2001/0999. 2008. Available online: https://www.agriculture.gov.au/sites/default/files/sitecollectiondocuments/fisheries/environment/bycatch/best-practice-references-keith-sainsbury.pdf (accessed on 20 May 2025).
- DFO. Guidelines for Implementing the Fish Stocks Provisions in the Fisheries Act. Last Updated 2022-08-23. 2022. Available online: https://www.dfo-mpo.gc.ca/reports-rapports/regs/sff-cpd/guidelines-lignes-directrices-eng.htm (accessed on 20 May 2025).
- MF (New Zealand Government, Ministry of Fisheries). Harvest Strategy Standard for New Zealand Fisheries. 2008. Available online: https://www.mpi.govt.nz/dmsdocument/728-Harvest-Strategy-Standard-for-New-Zealand-Fisheries (accessed on 20 May 2025).
- Restrepo, V.R.; Thompson, G.G.; Mace, P.M.; Gabriel, W.L.; Low, L.L.; MacCall, A.D.; Methot, R.D.; Powers, J.E.; Taylor, B.L.; Wade, P.R.; et al. Technical Guidance on the Use of Precautionary Approaches to Implementing National Standard 1 of the Magnuson-Stevens Fishery Conservation and Management Act. NOAA Technical Memorandum NMFS–F/SPO–31. 1998. Available online: https://www.st.nmfs.noaa.gov/Assets/stock/documents/Tech-Guidelines.pdf (accessed on 20 May 2025).
- Myers, R.A.; Rosenberg, A.A.; Mace, P.M.; Barrowman, N.; Restrepo, V.R. In search of thresholds for recruitment overfishing. ICES J. Mar. Sci. 1994, 51, 191–205. [Google Scholar] [CrossRef]
- Hilborn, R.; Walters, C.J. Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty; Chapman and Hall: New York, NY, USA, 1992; 570p. [Google Scholar]
- Hutchings, J.A. Renaissance of a caveat: Allee effects in marine fish. ICES J. Mar. Sci. 2014, 71, 2152–2157. [Google Scholar] [CrossRef]
- Allee, W.H. Animal Aggregations, A Study in General Sociology; University Chicago Press: Chicago, IL, USA, 1931. [Google Scholar]
- Hutchings, J.A. Thresholds for impaired species recovery. Proc. R. Soc. Ser. B Biol. Sci. 2015, 282, 20150654. [Google Scholar] [CrossRef] [PubMed]
- Liermann, M.; Hilborn, R. Depensation: Evidence, models and implications. Fish Fish. 2001, 2, 33–58. [Google Scholar] [CrossRef]
- Kronlund, A.R.; Forrest, R.E.; Cleary, J.S.; Grinnell, M.H. The selection and role of limit reference points for Pacific herring (Clupea pallasii) in British Columbia, Canada. DFO Canadian Science Advisory Secretariat Research Document. 2018/009. 2018. ix + 125 p. Available online: https://waves-vagues.dfo-mpo.gc.ca/library-bibliotheque/40685147.pdf (accessed on 20 May 2025).
- Myers, R.A.; Barrowman, N.J.; Hutchings, J.A.; Rosenberg, A.A. Population dynamics of exploited fish stocks at low population levels. Science 1995, 269, 1106–1108. [Google Scholar] [CrossRef]
- Liermann, M.; Hilborn, R. Depensation in fish stocks: A hierarchic Bayesian meta-analysis. Can. J. Fish. Aquat. Sci. 1997, 54, 1976–1984. [Google Scholar] [CrossRef]
- Hilborn, R.; Hively, D.J.; Jensen, O.P.; Branch, T.A. The dynamics of fish populations at low abundance and prospects for rebuilding and recovery. ICES J. Mar. Sci. 2014, 71, 2141–2151. [Google Scholar] [CrossRef]
- Ricard, D.; Minto, D.; Jensen, O.P.; Baum, J.K. Examining the knowledge base and status of commercially exploited marine species with the RAM Legacy Stock Assessment Database. Fish Fish. 2012, 13, 380–398. [Google Scholar] [CrossRef]
- Keith, D.M.; Hutchings, J.A. Population dynamics of marine fishes at low abundance. Can. J. Fish. Aquat. Sci. 2012, 69, 1150–1163. [Google Scholar] [CrossRef]
- Perälä, T.; Kuparinen, A. Detection of Allee effects in marine fishes: Analytical biases generated by data availability and model selection. Proc. R. Soc. Ser. B Biol. Sci. 2017, 284, 20171284. [Google Scholar] [CrossRef]
- Perälä, T.; Hutchings, J.A.; Kuparinen, A. Allee effects and the Allee-effect zone in northwest Atlantic cod. Biol. Lett. 2022, 18, 20210439. [Google Scholar] [CrossRef] [PubMed]
- Marentette, J.R.; Kronlund, A.R.; Cogliati, K.M. Specification of precautionary approach reference points and harvest control rules in domestically managed and assessed key harvested stocks in Canada. DFO Canadian Science Advisory Secretariat Research Document. 2021/057. 2021. vii + 98 p. Available online: https://publications.gc.ca/collections/collection_2021/mpo-dfo/fs70-5/Fs70-5-2021-057-eng.pdf (accessed on 20 May 2025).
- DFO. Stock Assessment of NAFO subdivision 3Ps cod. DFO Canadian Science Advisory Secretariat Science Advisory Report. 2020/018. Available online: https://publications.gc.ca/collections/collection_2020/mpo-dfo/fs70-6/Fs70-6-2020-018-eng.pdf (accessed on 20 May 2025).
- ICES. Report of the Herring Assessment Working Group for the Area South of 62ºN (HAWG), 10–19 March 2015; ICES CM 2015/ACOM:06; ICES: Copenhagen, Denmark, 2015. [Google Scholar]
- ICES. Report of the Baltic Fisheries Assessment Working Group (WGBFAS), 14–21 April 2015; ICES CM 2015/ACOM:10; ICES: Copenhagen, Denmark, 2015. [Google Scholar]
- Varkey, D.A.; Babyn, J.; Regular, P.; Ings, D.W.; Kumar, R.; Rogers, B.; Champagnat, J.; Morgan, M.J. A state-space model for stock assessment of cod (Gadus morhua) stock in NAFO subdivision 3Ps. DFO Canadian Science Advisory Secretariat Research Document. 2022/022. 2022. v + 78 p. Available online: https://waves-vagues.dfo-mpo.gc.ca/library-bibliotheque/4105829x.pdf (accessed on 20 May 2025).
- Neilson, A.; Berg, C.W. Estimation of time-varying selectivity in stock assessments using state-space models. Fish. Res. 2014, 158, 96–101. [Google Scholar] [CrossRef]
- Shepherd, J.G. Extended survivors analysis: An improved method for the analysis of catch-at-age data and abundance indices. ICES J. Mar. Sci. 1999, 56, 584–591. [Google Scholar] [CrossRef]
- Link, W.A. Modeling pattern in collections of parameters. J. Wildl. Manag. 1999, 63, 1017–1027. [Google Scholar] [CrossRef]
- Brooks, E.N.; Deroba, J.J. When “data” are not data: The pitfalls of post hoc analyses that use stock assessment model output. Can. J. Fish. Aquat. Sci. 2015, 72, 634–641. [Google Scholar] [CrossRef]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Ministry of Agriculture, Fisheries and Food. Fishery Investigations: London, UK, 1957; p. 533. [Google Scholar]
- Ricker, W.E. Stock and recruitment. J. Fish. Res. Board Can. 1954, 11, 559–623. [Google Scholar] [CrossRef]
- Saila, S.; Recksiek, C.W.; Prager, M.H. Basic Fishery Science Programs; Elsevier: Amsterdam, The Netherlands, 1988; 230p. [Google Scholar]
- Iles, T.C. A review of stock-recruitment relationships with reference to flatfish populations. Neth. J. Sea Res. 1994, 32, 399–420. [Google Scholar] [CrossRef]
- Punt, A.E.; Cope, J.M. Extending integrated stock assessment models to use non-depensatory three-parameter stock-recruitment relationships. Fish. Res. 2019, 217, 46–57. [Google Scholar] [CrossRef]
- Miller, T.J.; Brooks, E.N. Steepness is a slippery slope. Fish Fish. 2020, 22, 634–645. [Google Scholar] [CrossRef]
- Guo, J.; Gabry, J.; Goodrich, B.; Johnson, A.; Weber, S.; Badr, H.S.; Lee, D.; Sakrejda, K.; Martin, M.; Trustees of Columbia University; et al. R Interface to Stan. Package ‘rstan’, Version 2.32.7. 2025. Available online: https://cran.r-project.org/web/packages/rstan/rstan.pdf (accessed on 20 May 2025).
- Gabry, J.; Veen, D.; Stan Development Team; Andreae, M.; Betancourt, M.; Carpenter, B.; Gao, Y.; Gelman, A.; Goodrich, B.; Lee, D.; et al. Interactive Visual and Numerical Diagnostics and Posterior Analysis for Bayesian Models. Package ‘shinystan’. Version 2.6.0. 2022. Available online: https://cran.r-project.org/web/packages/shinystan/shinystan.pdf (accessed on 20 May 2025).
- Stan Development Team. Stan Reference Manual. 2025. Available online: https://mc-stan.org/ (accessed on 20 May 2025).
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef]
- Vehtari, A.; Gabry, J.; Magnusson, M.; Yao, Y.; Bürkner, P.-C.; Paananen, T.; Gelman, A.; Goodrich, B.; Piironen, J.; Nicenboim, B.; et al. Efficient Leave-One-out Cross-Validation and WAIC for Bayesian Models. Package ‘loo’. Version 2.8.0. 2024. Available online: https://cran.r-project.org/web/packages/loo/loo.pdf (accessed on 20 May 2025).
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Maunder, M.N. Stock-recruitment models from the viewpoint of density-dependent survival and the onset of strong density-dependence when a carrying capacity limit is reached. Fish. Res. 2022, 249, 106249. [Google Scholar] [CrossRef]
- Soetaert, K.; Hindmarch, A.C.; Eisenstat, S.C.; Moler, C.; Dongarra, J.; Saad, Y. Nonlinear Root Finding, Equilibrium and Steady-State Analysis of Ordinary Differential Equations. Package ‘rootSolve’. Version 1.8.2.3. 2022. Available online: https://cran.r-project.org/web/packages/rootSolve/rootSolve.pdf (accessed on 20 May 2025).
- Brooks, E.N.; Powers, J.E. Generalized compensation in stock-recruit functions: Properties and implications for management. ICES J. Mar. Sci. 2007, 64, 413–424. [Google Scholar] [CrossRef]
- Barrowman, N.J.; Myers, R.A. Still more spawner-recruitment curves: The hockey stick and its generalizations. Can. J. Fish. Aquat. Sci. 2000, 57, 665–676. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2025. Available online: https://www.R-project.org/ (accessed on 20 May 2025).
- Neubauer, P.; Jensen, O.P.; Hutchings, J.A.; Baum, J.K. Resilience and recovery of overexploited marine populations. Science 2013, 340, 347–349. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J.; Freckleton, R.P. What is the Allee effect? Oikos 1999, 87, 185–190. [Google Scholar] [CrossRef]
- Holt, C.A.; Michielsens, C.G.J. Impact of time-varying productivity on estimated stock–recruitment parameters and biological reference points. Can. J. Fish. Aquat. Sci. 2020, 77, 836–847. [Google Scholar] [CrossRef]
- Szuwalski, C.S.; Britten, G.L.; Licandeo, R.; Amoroso, R.O.; Hilborn, R.; Walters, C. Global forage fish recruitment dynamics: A comparison of methods, time-variation, and reverse causality. Fish. Res. 2019, 214, 56–64. [Google Scholar] [CrossRef]
- Punt, A.E. Refocusing stock assessment in support of policy evaluation. In Fisheries for Global Welfare and Environment, Proceedings of the 5th World Fisheries Congress 2008, Yokohama, Japan, 20–24 October 2008; Tsukamoto, K., Kawamura, T., Takeuchi, T., Beard, T.D., Kaiser, M.J., Eds.; TERRAPUB: Tokyo, Japan, 2008; pp. 139–152. [Google Scholar]
- Butterworth, D.S.; Punt, A.E. Experiences in the evaluation and implementation of management procedures. ICES J. Mar. Sci. 1999, 56, 985–998. [Google Scholar] [CrossRef]
- Punt, A.E.; Butterworth, D.S.; de Moor, C.L.; Oliveira, J.A.A.; Haddon, M. Management strategy evaluation: Best practices. Fish Fish. 2016, 17, 303–334. [Google Scholar] [CrossRef]
- van Deurs, M.; Brooks, M.E.; Lindengren, M.; Henriksen, O.; Rindorf, A. Biomass limit reference points are sensitive to estimation method, time-series length and stock development. Fish Fish. 2021, 22, 18–30. [Google Scholar] [CrossRef]
- ICES. Workshop on ICES Reference Points (WKREF1). ICES Scientific Reports. 2022, Volume 4, Issue 2, 70p. Available online: http://doi.org/10.17895/ices.pub.9822 (accessed on 20 May 2025).
- Haltuch, M.A.; Punt, A.E.; Dorn, M.W. Evaluating alternative estimators of fishery management reference points. Fish. Res. 2008, 94, 290–303. [Google Scholar] [CrossRef]
- Punt, A.E.; Dorn, M.W.; Haltuch, M.A. Evaluation of threshold management strategies for groundfish off the U.S. West Coast. Fish. Res. 2008, 94, 251–266. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stock | SRR | FMSY | Fcrash | 0.2B0 (kt) | 0.4BMSY (kt) | BCP (kt) | BAT (kt) | Binflection (kt) | BAET (kt) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BS herring | BH | 0.165 | 0.553 | 279 | 207 | - | - | - | - | - | - | - | - |
| sBH | 0.175 | 0.269 | 309 | 249 | - | 35.2 | 151 | 271 | 0.023 | 0.17 | 0.43 | 0.59 | |
| Ricker | 0.225 | 0.570 | 193 | 170 | - | - | - | - | - | - | - | - | |
| SL | 0.227 | 0.286 | 190 | 204 | - | 58.7 | 189 | 320 | 0.062 | 0.34 | 0.63 | 0.70 | |
| HS | - | - | - | - | 455 | - | - | - | - | - | - | - | |
| GoR herring | BH | 0.262 | 1.06 | 106 | 67.3 | - | - | - | - | - | - | - | - |
| sBH | 0.342 | 0.565 | 104 | 64.8 | - | 10.5 | 40.1 | 68.2 | 0.020 | 0.13 | 0.42 | 0.84 | |
| Ricker | 0.458 | 1.20 | 45.6 | 36.2 | - | - | - | - | - | - | - | - | |
| SL | 0.497 | 0.584 | 37.2 | 39.7 | - | 18.6 | 47.0 | 72.7 | 0.10 | 0.39 | 0.73 | 0.89 | |
| HS | - | - | - | - | 81.6 | - | - | - | - | - | - | - | |
| NSAS herring | BH | 0.791 | 3.97 | 2134 | 1426 | - | - | - | - | - | - | - | - |
| sBH | 1.26 | 3.97 | 1464 | 852 | - | 0 | 226 | 385 | 0 | 0.053 | 0.18 | 0.49 | |
| Ricker | 1.30 | 3.60 | 1038 | 912 | - | - | - | - | - | - | - | - | |
| SL | 1.23 | 4.48 | 1101 | 956 | - | - 1 | - 1 | - 1 | - 1 | - 1 | - 1 | - 1 | |
| HS | - | - | - | - | 786 | - | - | - | - | - | - | - | |
| WBSS herring | BH | 0.315 | 1.69 | 242 | 136 | - | - | - | - | - | - | - | - |
| sBH | 0.385 | 0.744 | 244 | 128 | - | 25.2 | 71.9 | 114 | 0.021 | 0.093 | 0.36 | 0.84 | |
| Ricker | 0.424 | 1.043 | 134 | 110 | - | - | - | - | - | - | - | - | |
| SL | 0.489 | 0.73 | 116 | 105 | - | 0 | 57.1 | 102 | 0 | 0.17 | 0.39 | 0.75 | |
| HS | - | - | - | - | 136 | - | - | - | - | - | - | - | |
| 3Ps cod | sBH | 0.252 | 0.384 | 149 | 130 | - | 27.1 | 89.0 | 159 | 0.036 | 0.21 | 0.49 | 0.81 |
| SL | 0.289 | 0.379 | 94.4 | 104 | - | 29.5 | 92.1 | 160 | 0.063 | 0.34 | 0.62 | 0.81 | |
| None * | - | - | - | - | 196 | - | - | - | - | - | - | - |
| Stock | SRR | 0.2B0 (kt) | 0.4BMSY (kt) | BAET * (kt) | ||
|---|---|---|---|---|---|---|
| BS herring | BH | 279 | 207 | 271 | 0.19 | 0.52 |
| Ricker | 193 | 170 | 320 | 0.33 | 0.75 | |
| GoR herring | BH | 106 | 67.3 | 68.2 | 0.13 | 0.41 |
| Ricker | 45.6 | 36.2 | 72.7 | 0.32 | 0.80 | |
| NSAS herring | BH | 2134 | 1426 | 385 | 0.036 | 0.11 |
| Ricker | 1038 | 912 | - | - | - | |
| WBSS herring | BH | 242 | 136 | 114 | 0.094 | 0.33 |
| Ricker | 134 | 110 | 102 | 0.15 | 0.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrett, T.J.; Huynh, Q.C. Limit Reference Points and Equilibrium Stock Dynamics in the Presence of Recruitment Depensation. Fishes 2025, 10, 342. https://doi.org/10.3390/fishes10070342
Barrett TJ, Huynh QC. Limit Reference Points and Equilibrium Stock Dynamics in the Presence of Recruitment Depensation. Fishes. 2025; 10(7):342. https://doi.org/10.3390/fishes10070342
Chicago/Turabian StyleBarrett, Timothy J., and Quang C. Huynh. 2025. "Limit Reference Points and Equilibrium Stock Dynamics in the Presence of Recruitment Depensation" Fishes 10, no. 7: 342. https://doi.org/10.3390/fishes10070342
APA StyleBarrett, T. J., & Huynh, Q. C. (2025). Limit Reference Points and Equilibrium Stock Dynamics in the Presence of Recruitment Depensation. Fishes, 10(7), 342. https://doi.org/10.3390/fishes10070342