
Impacts of Low-Order Stream Connectivity Restoration Projects on Aquatic Habitat and Fish Diversity

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
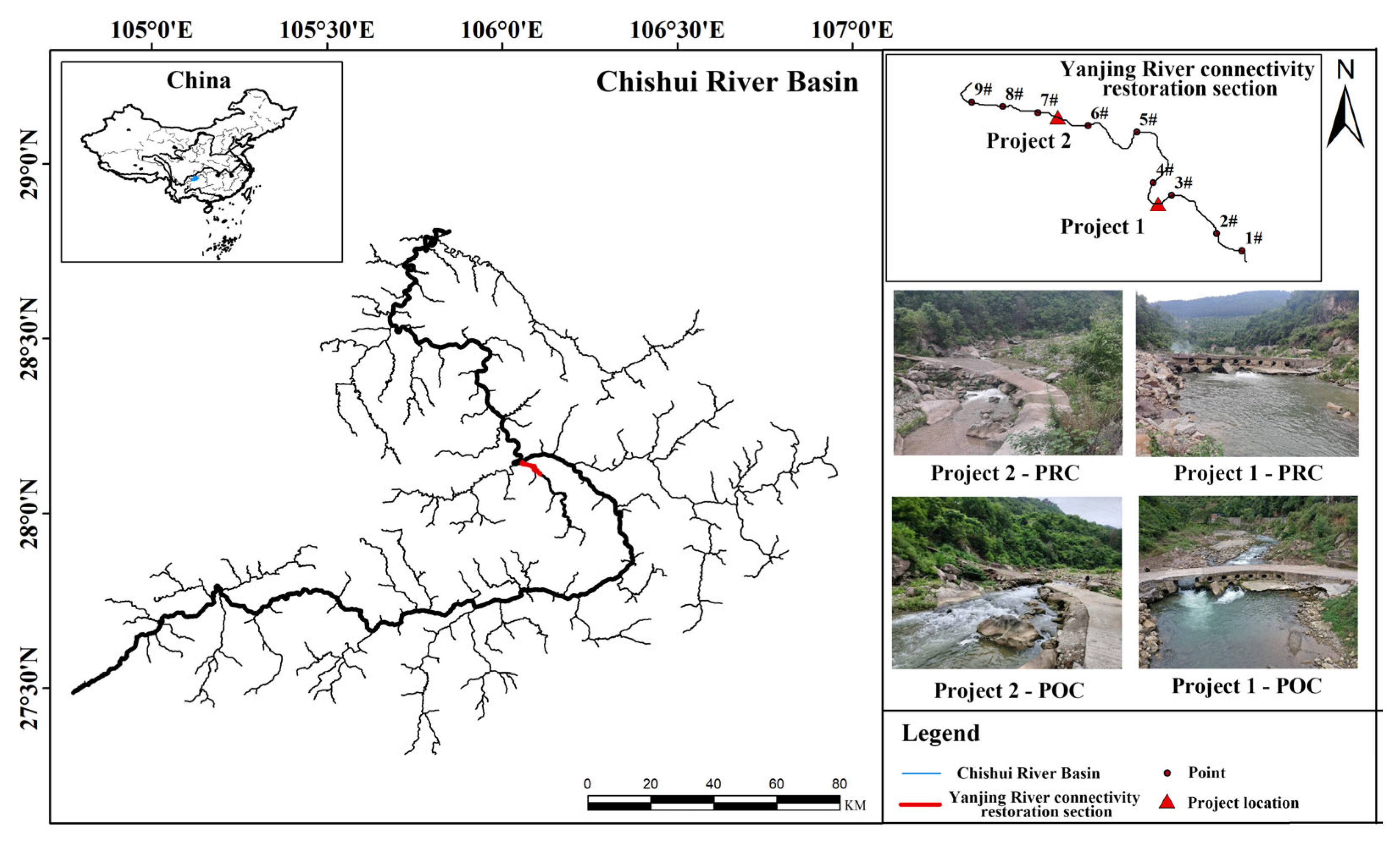
2.1. Connectivity Project Implementation Area
2.2. Connectivity Restoration Measures
2.3. Sampling Site Setup and Sampling Timing
2.4. Measurement of Aquatic Environmental Factors
2.5. Fish Sampling and Data Collection
2.6. Selection of Functional Traits
2.7. Fish Diversity
2.7.1. Selection of Species Diversity Indices
2.7.2. Selection of Functional Diversity Indices
2.8. Data Analysis Methods and Software
2.8.1. Differential Analysis
2.8.2. Principal Component Analysis (PCA)
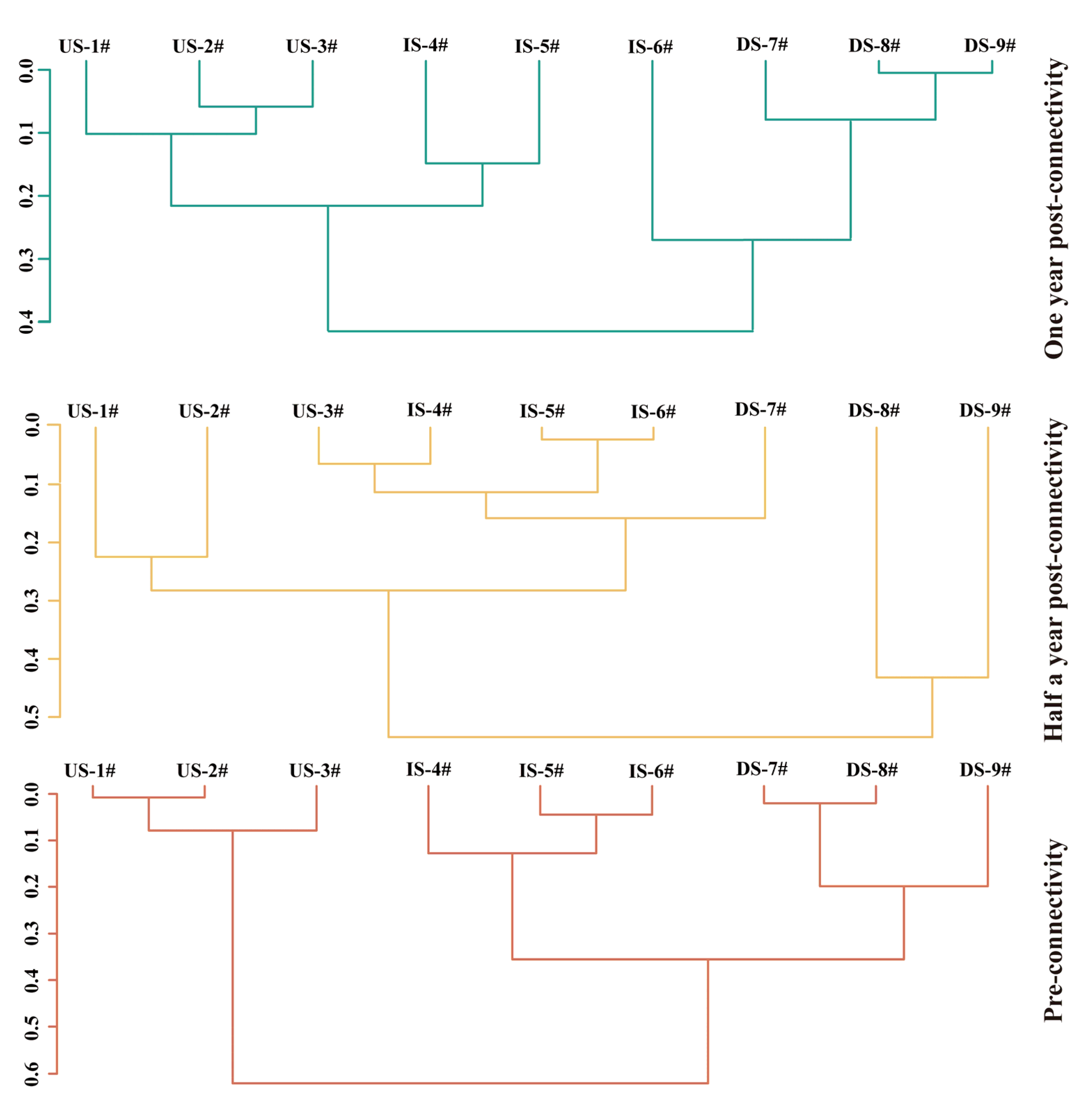
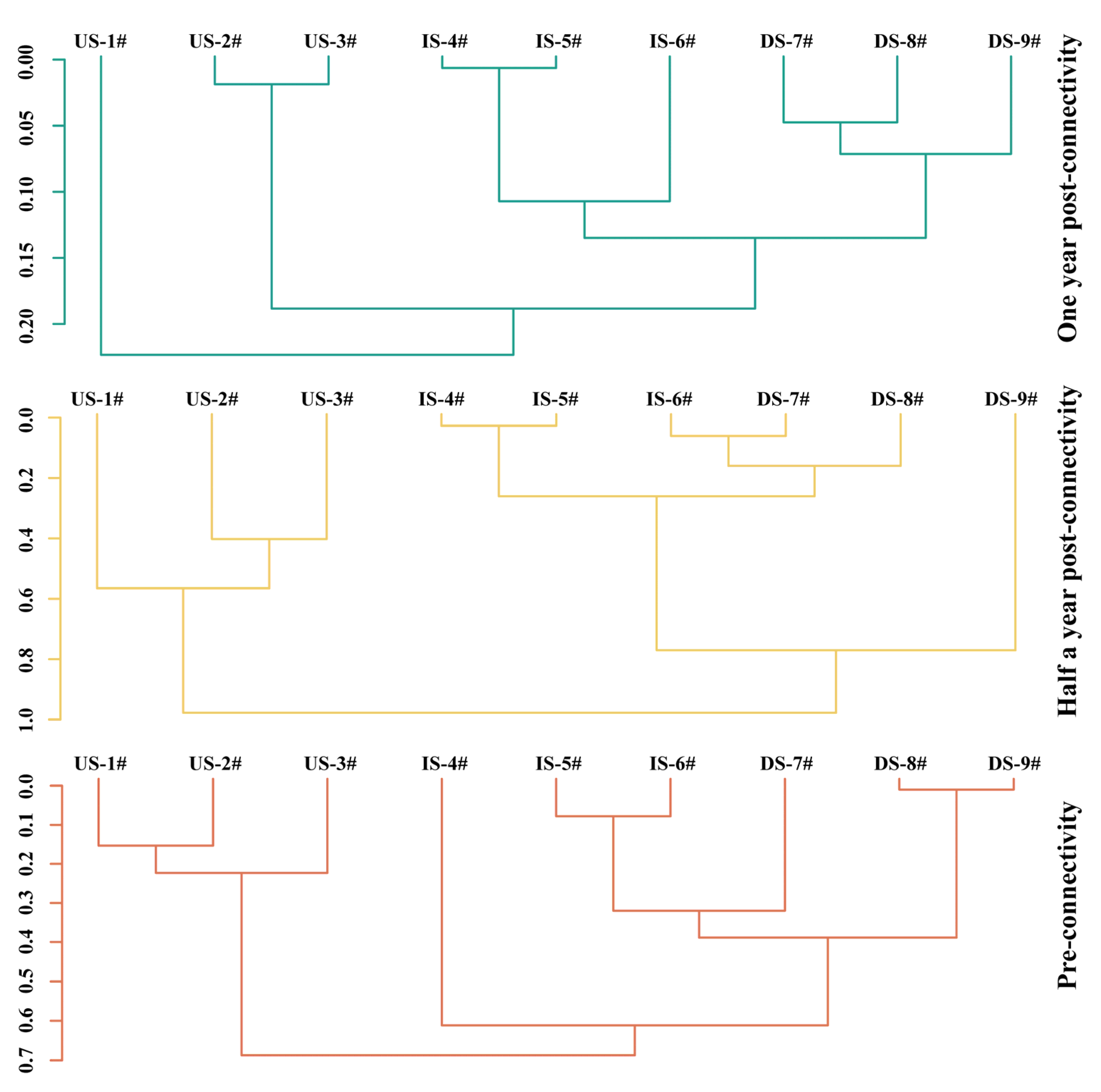
2.8.3. Constrained Sequential Clustering (CONISS)
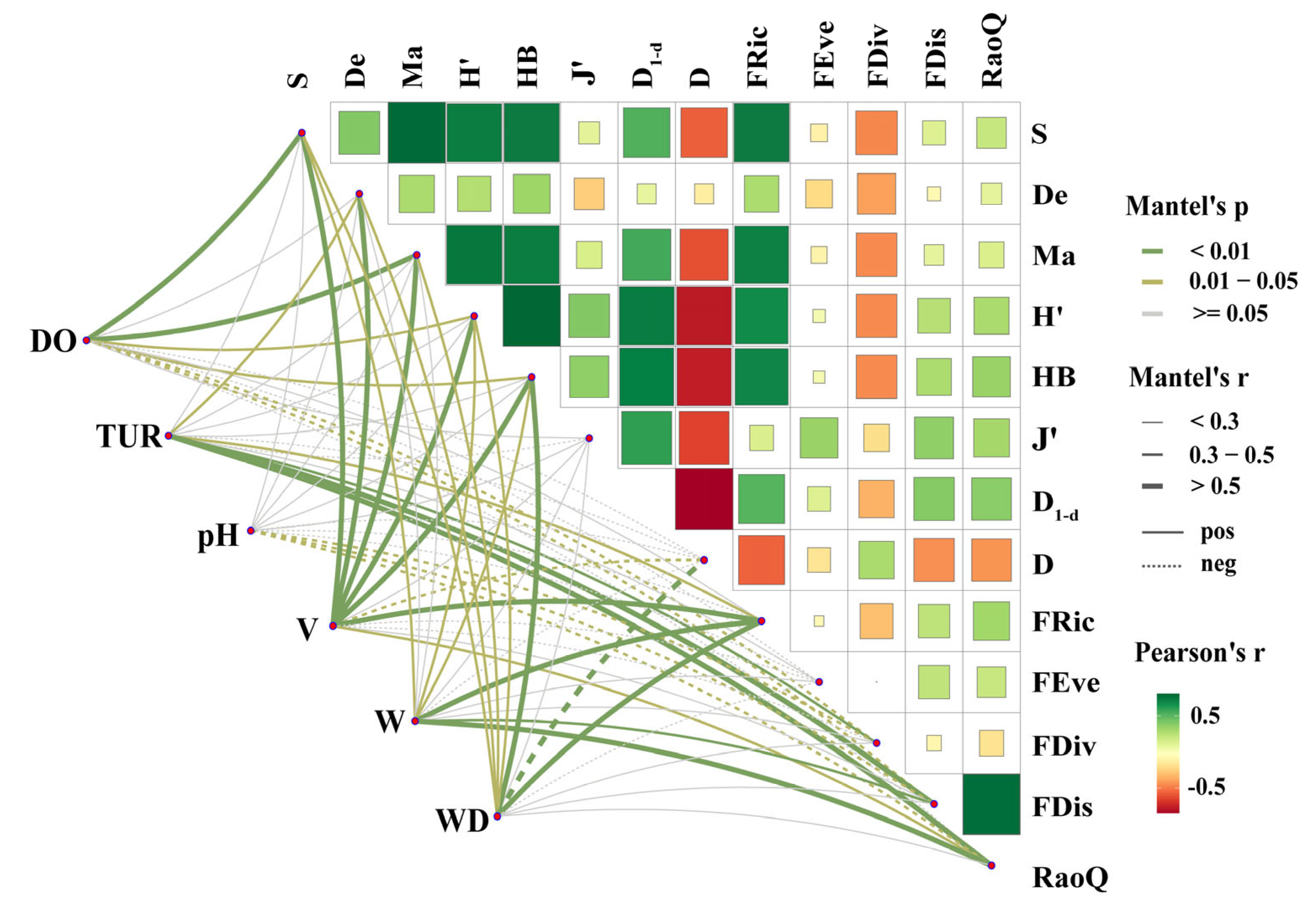
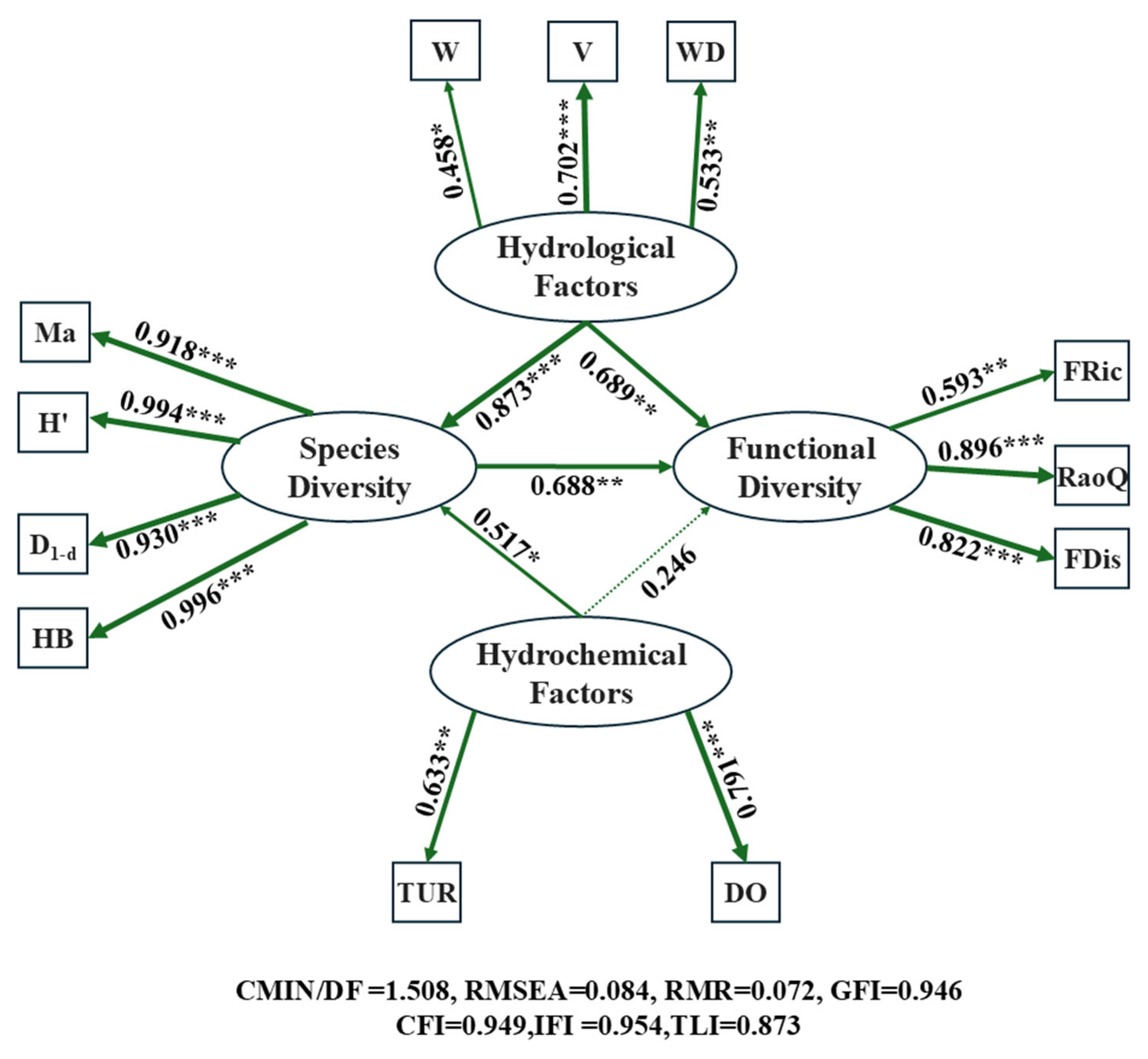
2.8.4. Mantel Test and Structural Equation Modeling Construction
3. Results
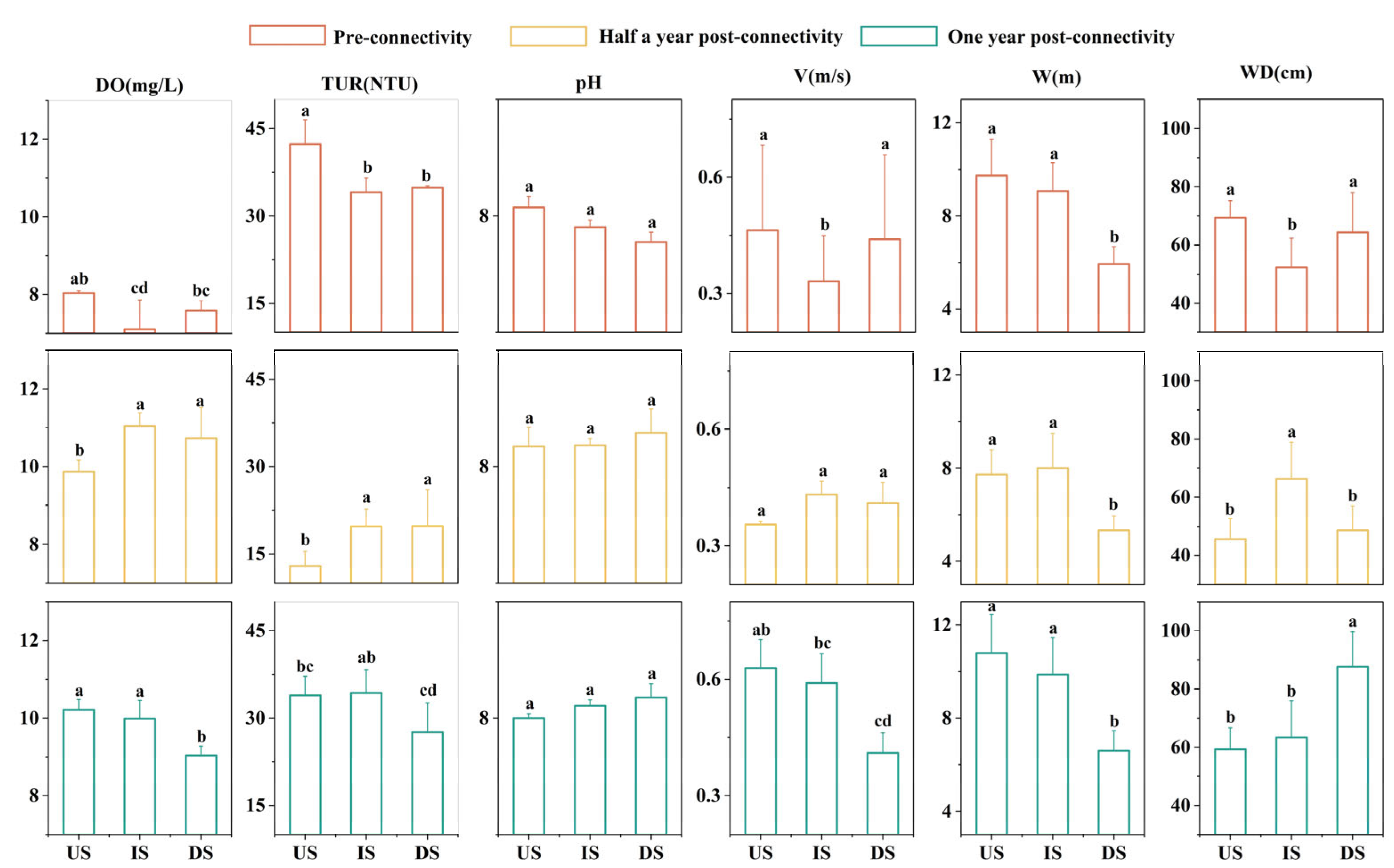
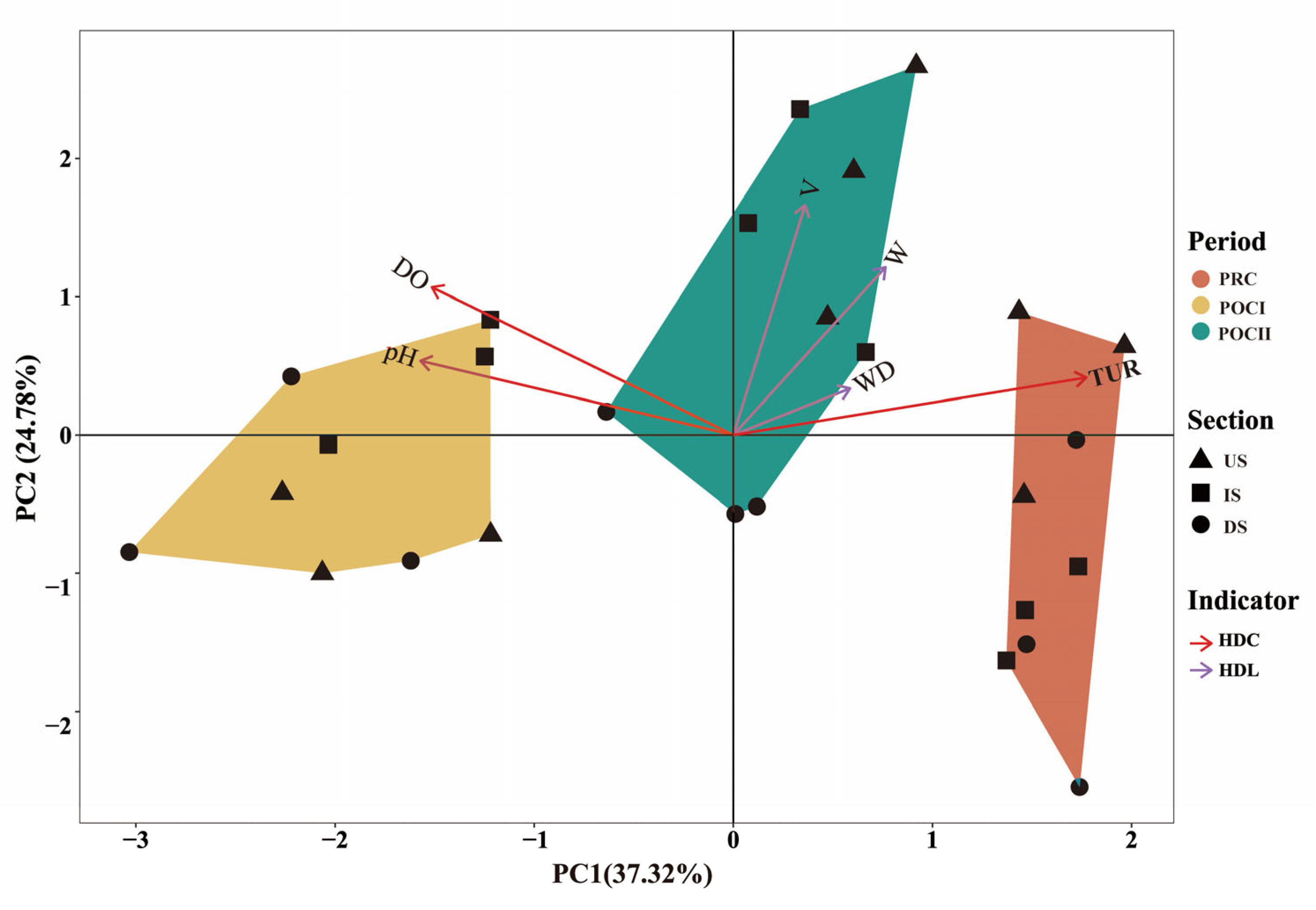
3.1. Variation Characteristics of Aquatic Environmental Factors
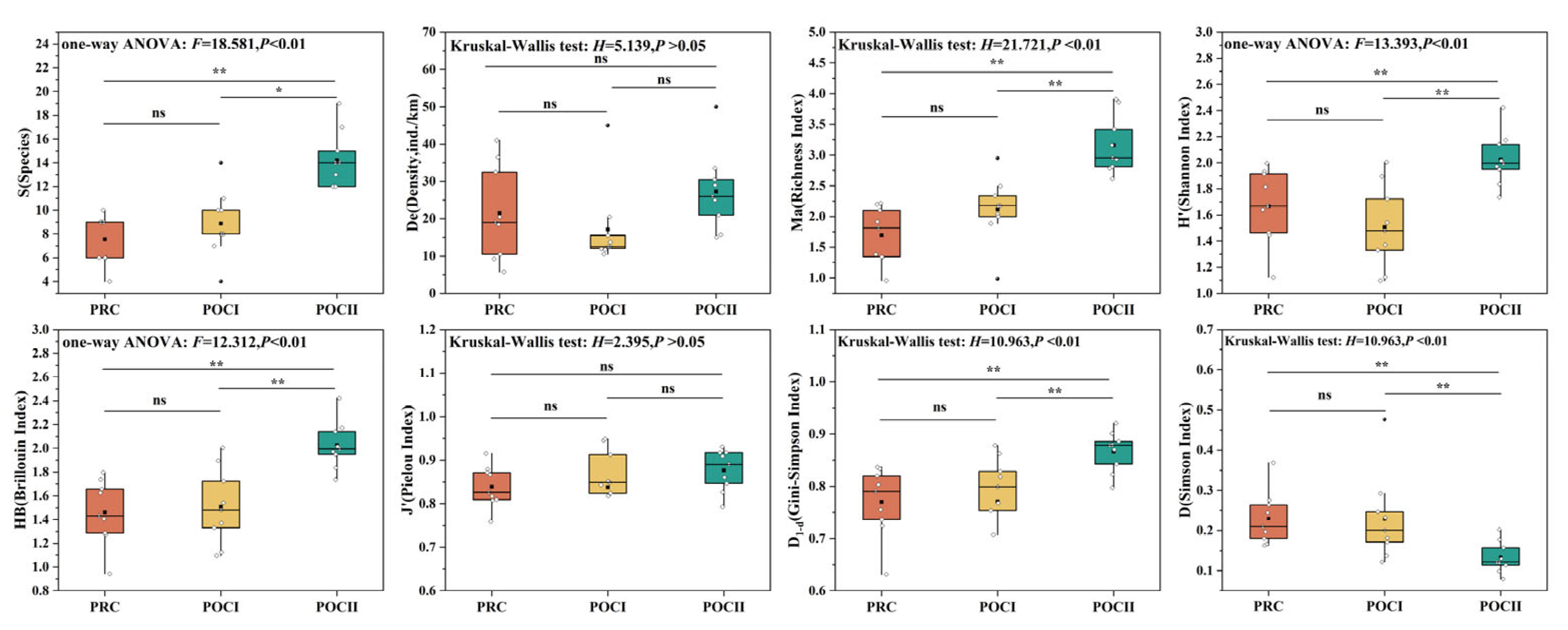
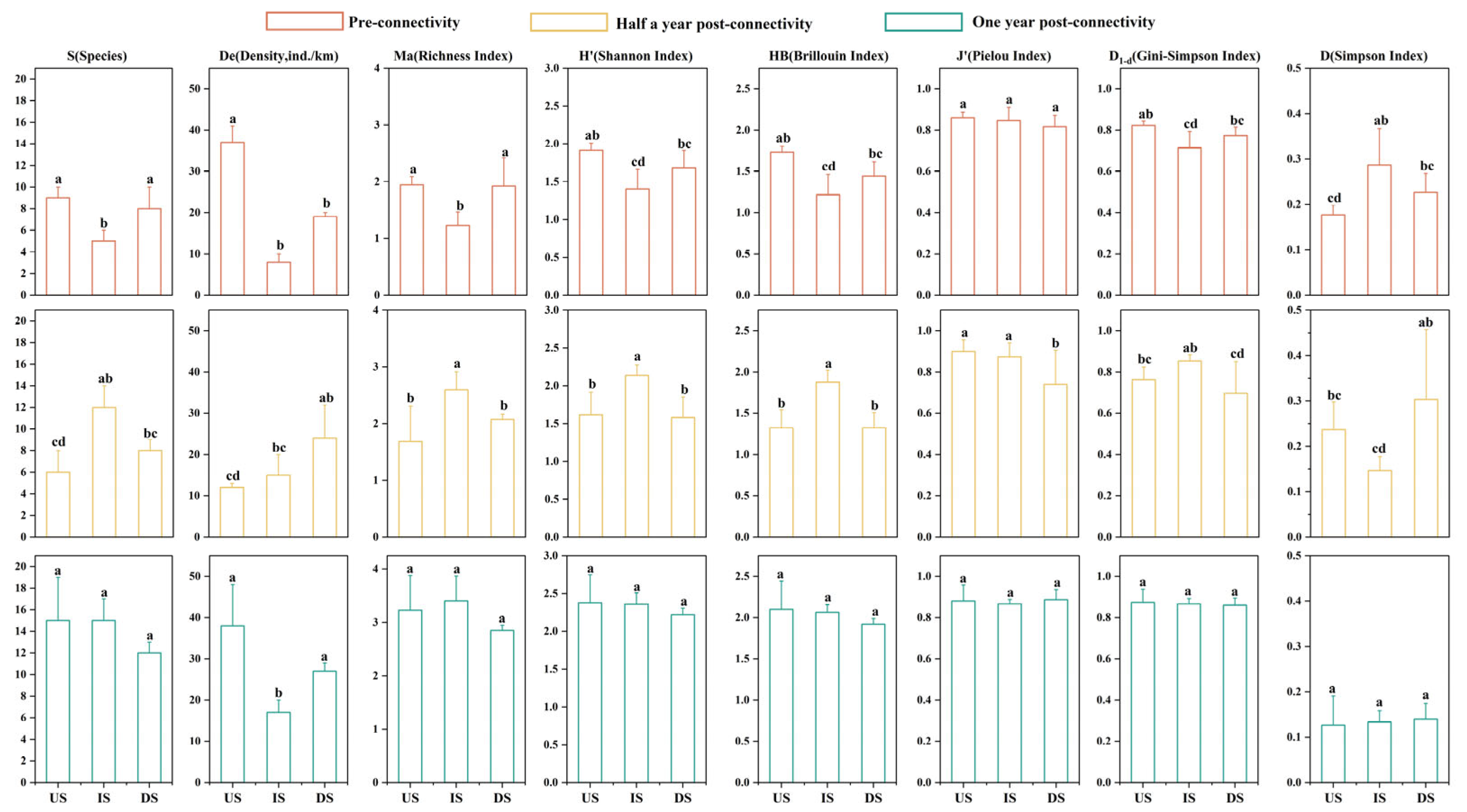
3.2. Variation Characteristics of Fish Species Diversity
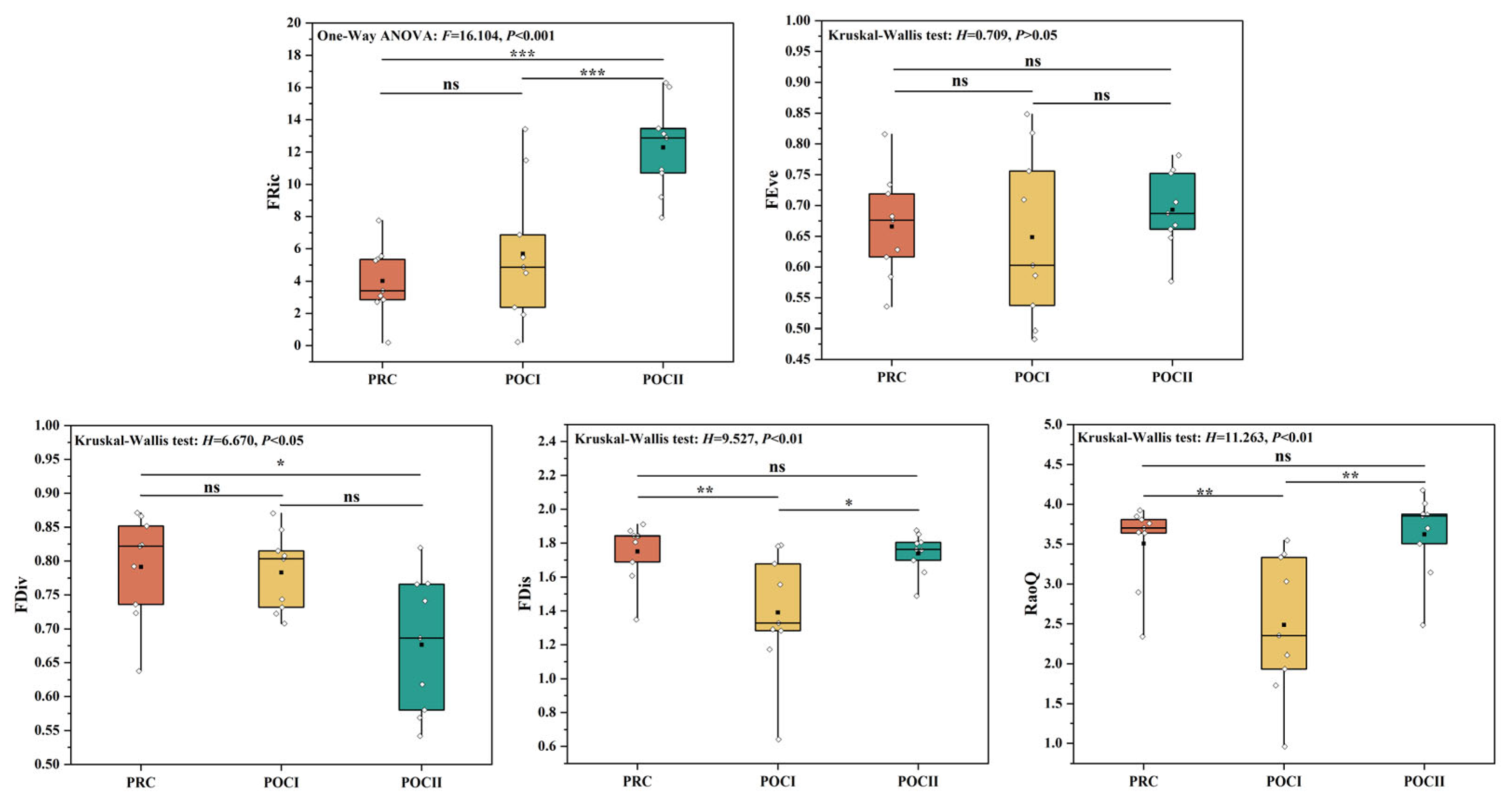
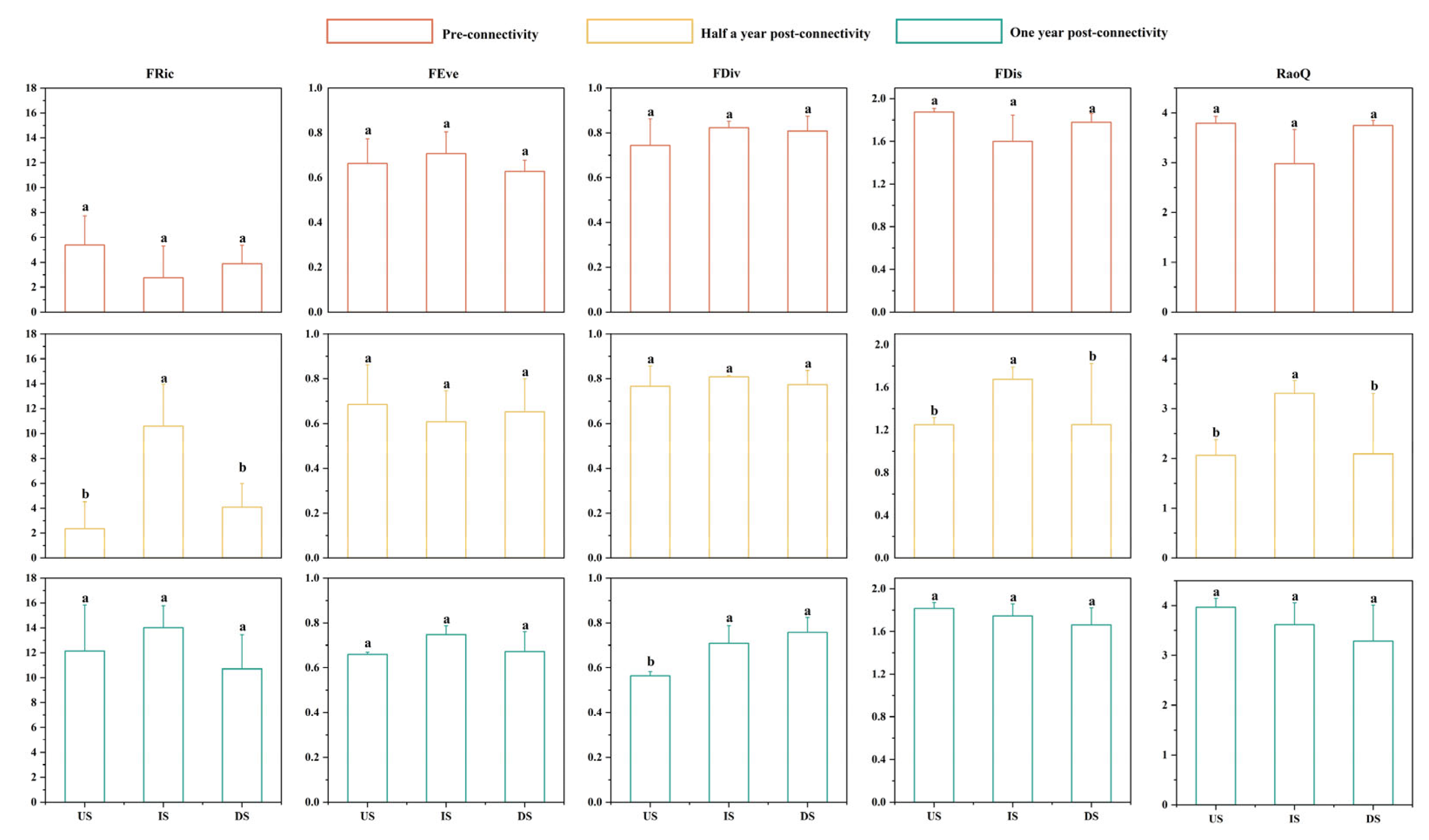
3.3. Variation Characteristics of Fish Functional Diversity
3.4. Analysis of the Intrinsic Relationship Between Aquatic Habitat and Fish Diversity
4. Discussion
4.1. Changes in Aquatic Habitat and Fish Diversity Before and After River Connectivity Restoration
4.2. Analysis of Heterogeneity Between River Sections Formed by River Blockages After River Connectivity
4.3. The Internal Relationships Between Aquatic Habitat and Fish Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Garcia De Leaniz, C.; Wantzen, K.M.; Wolter, C.; Tharme, R.E.; Zalewski, M.; Belletti, B. Editorial: Challenges and Benefits of Restoring River Connectivity. Front. Ecol. Evol. 2023, 11, 1110413. [Google Scholar] [CrossRef]
- Kujanová, K.; Matoušková, M. Improvement in Physical River Habitat Quality in Response to River Restoration Measures. Geografie 2016, 121, 54–78. [Google Scholar] [CrossRef]
- Belletti, B.; Garcia De Leaniz, C.; Jones, J.; Bizzi, S.; Börger, L.; Segura, G.; Castelletti, A.; Van De Bund, W.; Aarestrup, K.; Barry, J.; et al. More than One Million Barriers Fragment Europe’s Rivers. Nature 2020, 588, 436–441. [Google Scholar] [CrossRef]
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the World’s Free-Flowing Rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef]
- Poff, N.L.; Hart, D.D. How Dams Vary and Why It Matters for the Emerging Science of Dam Removal. Bioscience 2002, 52, 659–668. [Google Scholar] [CrossRef]
- Ioannidou, C.T.; Neeson, T.M.; O’Hanley, J.R. Boosting Large-Scale River Connectivity Restoration by Planning for the Presence of Unrecorded Barriers. Conserv. Biol. 2023, 37, e14093. [Google Scholar] [CrossRef]
- McKay, S.K.; Martin, E.H.; McIntyre, P.B.; Milt, A.W.; Moody, A.T.; Neeson, T.M. A Comparison of Approaches for Prioritizing Removal and Repair of Barriers to Stream Connectivity. River Res. Appl. 2020, 36, 1754–1761. [Google Scholar] [CrossRef]
- Wohl, E.; Fryirs, K.; Grabowski, R.C.; Morrison, R.R.; Sear, D. Enhancing the Natural Absorbing Capacity of Rivers to Restore Their Resilience. Bioscience 2024, 74, 782–796. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Nielsen, J.; Frandsen, S.B.; Olsen, H.-M.; Aarestrup, K. Catchment-Scale Effects of River Fragmentation: A Case Study on Restoring Connectivity. J. Environ. Manage. 2020, 264, 110408. [Google Scholar] [CrossRef]
- Tummers, J.S.; Hudson, S.; Lucas, M.C. Evaluating the Effectiveness of Restoring Longitudinal Connectivity for Stream Fish Communities: Towards a More Holistic Approach. Sci. Total Environ. 2016, 569–570, 850–860. [Google Scholar] [CrossRef]
- Drouineau, H.; Carter, C.; Rambonilaza, M.; Beaufaron, G.; Bouleau, G.; Gassiat, A.; Lambert, P.; Le Floch, S.; Tétard, S.; De Oliveira, E. River Continuity Restoration and Diadromous Fishes: Much More than an Ecological Issue. Environ. Manag. 2018, 61, 671–686. [Google Scholar] [CrossRef]
- Kunakh, O.M.; Volkova, A.M.; Tutova, G.F.; Zhukov, O.V. Diversity of diversity indices: Which diversity measure is better? Biosyst. Divers. 2023, 31, 131–146. [Google Scholar] [CrossRef]
- Rodeles, A.A.; Miranda, R.; Galicia, D. Barriers to Longitudinal River Connectivity: Review of Impacts, Study Methods and Management for Iberian Fish Conservation. Limnetica 2020, 39, 601–619. [Google Scholar] [CrossRef]
- Wegscheider, B.; Waldock, C.; Calegari, B.B.; Josi, D.; Brodersen, J.; Seehausen, O. Neglecting Biodiversity Baselines in Longitudinal River Connectivity Restoration Impacts Priority Setting. Sci. Total Environ. 2024, 954, 175167. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, C.S.; Barraud, R.; Germaine, M.-A. Dam Removals and River Restoration in International Perspective. Water Altern. 2017, 10, 648–654. [Google Scholar]
- Gessner, J.; Zahn, S.; Jaric, I.; Wolter, C. Estimating the Potential for Habitat Restoration and Connectivity Effects on European Sturgeon (Acipenser Sturio L. 1758) Population Rehabilitation in a Lowland River-the Havel, Germany. J. Appl. Ichthyol. 2014, 30, 1473–1482. [Google Scholar] [CrossRef]
- Besacier-Monbertrand, A.-L.; Paillex, A.; Castella, E. Short-term impacts of lateral hydrological connectivity restoration on aquatic macroinvertebrates. River Res. Appl. 2012, 30, 557–570. [Google Scholar] [CrossRef]
- Pohl, M.M. Bringing down our dams: Trends in american dam removal rationales. J. Am. Water Resour. Assoc. 2002, 38, 1511–1519. [Google Scholar] [CrossRef]
- Bellmore, J.R.; Pess, G.R.; Duda, J.J.; O’Connor, J.E.; East, A.E.; Foley, M.M.; Wilcox, A.C.; Major, J.J.; Shafroth, P.B.; Morley, S.A. Conceptualizing Ecological Responses to Dam Removal: If You Remove It, What’s to Come? Bioscience 2019, 69, 26–39. [Google Scholar] [CrossRef]
- Major, J.J.; East, A.E.; O’Connor, J.E.; Grant, G.E.; Wilcox, A.C.; Magirl, C.S.; Collins, M.J.; Tullos, D.D. Geomorphic Responses to Dam Removal in the United States—A Two-Decade Perspective. In Gravel-Bed Rivers; Tsutsumi, D., Laronne, J.B., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 355–383. [Google Scholar]
- Wood, D.M.; Welsh, A.B.; Todd Petty, J. Genetic Assignment of Brook Trout Reveals Rapid Success of Culvert Restoration in Headwater Streams. North Am. J. Fish. Manag. 2018, 38, 991–1003. [Google Scholar] [CrossRef]
- Qiu, J.; Yuan, S.; Tang, H.; Zhang, Q.; Wolter, C.; Nikora, V. Ecological Connectivity of River-Lake Ecosystem: Evidence from Fish Population Dynamics in a Connecting Channel. Water Resour. Res. 2024, 60, e2024WR037495. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Candee, M.M.; Baktoft, H.; Larsen, M.H.; Koed, A.; Aarestrup, K. River Connectivity Reestablished: Effects and Implications of Six Weir Removals on Brown Trout Smolt Migration. River Res. Appl. 2018, 34, 548–554. [Google Scholar] [CrossRef]
- Branco, P.; Segurado, P.; Santos, J.M.; Ferreira, M.T. Prioritizing Barrier Removal to Improve Functional Connectivity of Rivers. J. Appl. Ecol. 2014, 51, 1197–1206. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The Future of Fish Passage Science, Engineering, and Practice. Fish Fish. 2017, 19, 340–362. [Google Scholar] [CrossRef]
- Downing, J. Global Abundance and Size Distribution of Streams and Rivers. Inland Waters 2012, 2, 229–236. [Google Scholar] [CrossRef]
- Harvey, J.; Gomez-Velez, J.; Schmadel, N.; Scott, D.; Boyer, E.; Alexander, R.; Eng, K.; Golden, H.; Kettner, A.; Konrad, C.; et al. How Hydrologic Connectivity Regulates Water Quality in River Corridors. J. Am. Water Resour. Assoc. 2018, 55, 369–381. [Google Scholar] [CrossRef]
- Aftabuddin, M.; Hassan, M.A.; Das, A.K.; Jha, B.C.; Sharma, A.P. Effect of River Connectivity on Hydrochemistry, Sediment Enzyme Activity and Biotic Communities of Wetlands. Aquat. Ecosyst. Health Manag. 2017, 20, 140–150. [Google Scholar] [CrossRef]
- Lei, Y.; Dong, F.; Liu, X.; Ma, B.; Huang, W. Short-Term Variations and Correlations in Water Quality after Dam Removal in the Chishui River Basin. J. Environ. Manag. 2023, 327, 116917. [Google Scholar] [CrossRef]
- Yu, F.; Liu, F.; Xia, Z.; Lin, P.; Xu, C.; Wang, J.; Hou, M.; Zou, X. Classification and Assessment Methods for Mountain Channel Habitats in the Chishui River Basin, China. Water 2022, 14, 515. [Google Scholar] [CrossRef]
- Yujun, Y.; Yanning, G.; Shanghong, Z. The Impact of Dams on the River Connectivity of the Two Largest River Basins in China. River Res. Appl. 2021, 38, 185–193. [Google Scholar] [CrossRef]
- SL 58-2014; Specification for Geodesic Survey in Hydrology. China Water&Power Press: Beijing, China, 2014.
- Chu, Y.; Sun, Z.; He, X.; Jin, W.; Cheng, Y. Colored Atlas of Fishes in Sichuan, China: Volumes I and II; C.S.P.M.: Beijing, China, 2021. [Google Scholar]
- Guo, X.; Cheng, Y. The Fishes of Yunnan, China: Volumes I and II; C.S.P.M.: Beijing, China, 1990. [Google Scholar]
- Wu, L.; Li, D.; Zhao, Z. The Fishes of Guizhou, China; Guizhou People’s Publishing House: Guizhou, China, 1989. [Google Scholar]
- Villéger, S.; Miranda, J.R.; Hernández, D.F.; Mouillot, D. Contrasting Changes in Taxonomic vs. Functional Diversity of Tropical Fish Communities after Habitat Degradation. Ecol. Appl. 2010, 20, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Villéger, S.; Grenouillet, G.; Brosse, S. Decomposing Functional β-Diversity Reveals That Low Functional β-Diversity Is Driven by Low Functional Turnover in European Fish Assemblages: Decomposing Functional β-Diversity. Glob. Ecol. Biogeogr. 2013, 22, 671–681. [Google Scholar] [CrossRef]
- Bower, L.M.; Marion, C.A.; Scott, M.; Kubach, K.; Gelder, A. Fish Assemblage and Functional Trait Responses to Small-Dam Removal. Freshw. Biol. 2024, 69, 1043–1056. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Wu, Z.; Zhao, M.; Feng, G. Community Structure Characteristics and Changes in Fish Species at Poyang Lake after the Yangtze River Fishing Ban. Fishes 2024, 9, 281. [Google Scholar] [CrossRef]
- Vakal, A.; Govorun, O.; Gulevets, D.; Korchynska, Z.; Kushch, Y. The Impact of the Ecosystem on Biodiversity Restoration in the Natural Ecosystems of Ukraine. Grassroots J. Nat. Resour. 2025, 8, 945–963. [Google Scholar] [CrossRef]
- Happel, A. Increasing Fish Diversity of Chicago’s Waterways. Knowl. Manag. Aquat. Ecosyst. 2022, 423, 6. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.; Villéger, S.; Sun, I.; Thorn, S.; Lin, Y.; Chiang, J.; Sherwin, W.B. An Attribute-diversity Approach to Functional Diversity, Functional Beta Diversity, and Related (Dis)Similarity Measures. Ecol. Monogr. 2019, 89, e01343. [Google Scholar] [CrossRef]
- Zhang, X.; Xue, Y.; Zhang, C.; Ren, Y.; Xu, B.; Chen, Y. Sampling Intensity Influences the Estimation of Functional Diversity Indices of Fish Communities. Ecol. Indic. 2021, 121, 107169. [Google Scholar] [CrossRef]
- Grimm, E.C. CONISS: A FORTRAN 77 Program for Stratigraphically Constrained Cluster Analysis by the Method of Incremental Sum of Squares. Comput. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Shijie, P.; Lei, W.; Yongkun, L.; Ruowen, W.; Tianming, G.; Zongjun, G. A Study on Ecohydrological Mutual Feedback Relationship of the Shangdong River Basin Based on Hydrological Connectivity. Sci. Total Environ. 2024, 927, 171957. [Google Scholar] [CrossRef]
- Deng, X. Correlations between Water Quality and the Structure and Connectivity of the River Network in the Southern Jiangsu Plain, Eastern China. Sci. Total Environ. 2019, 664, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, Q.; Xu, Y.; Lu, M.; Lin, Z.; Gao, B. Dynamic Impacts of Changes in River Structure and Connectivity on Water Quality under Urbanization in the Yangtze River Delta Plain. Ecol. Indic. 2022, 135, 108582. [Google Scholar] [CrossRef]
- Yang, S.; Yang, G.; Li, B.; Wan, R. Water Quality Improves with Increased Spatially Surface Hydrological Connectivity in Plain River Network Areas. J. Environ. Manag. 2025, 377, 124703. [Google Scholar] [CrossRef]
- Beschta, R.L.; Platts, W.S. Morphological features of small streams: Significance and function. J. Am. Water Resour. Assoc. 1986, 22, 369–379. [Google Scholar] [CrossRef]
- Zhang, K.; Hu, M.; Yu, D.; Bao, Y. A Method for Detecting Navigable Areas in Narrow Rivers under Complex Reflection Conditions. Eng. Rep. 2024, 6, e12959. [Google Scholar] [CrossRef]
- Xiongjun, L.; Suping, P.; Lvliu, L.; Ye, Z. Variation Law of Organic Contaminant in Lijiang River. J. China Univ. Min. Technol. 2004, 14, 133–137. [Google Scholar]
- Nabi, M. Computational Modelling of Small-Scale River Morphodynamics. Ph.D. Thesis, Delft University of Technology, Delft, The Netherlands, 2012. [Google Scholar]
- Raffard, A.; Santoul, F.; Cucherousset, J.; Blanchet, S. The Community and Ecosystem Consequences of Intraspecific Diversity: A Meta-analysis. Biol. Rev. 2018, 94, 648–661. [Google Scholar] [CrossRef]
- Song, Y.; Wang, P.; Li, G.; Zhou, D. Relationships between Functional Diversity and Ecosystem Functioning: A Review. Acta Ecol. Sin. 2014, 34, 85–91. [Google Scholar] [CrossRef]
- Stoffers, T.; Buijse, A.D.; Geerling, G.W.; Jans, L.H.; Schoor, M.M.; Poos, J.J.; Verreth, J.A.J.; Nagelkerke, L.A.J. Freshwater Fish Biodiversity Restoration in Floodplain Rivers Requires Connectivity and Habitat Heterogeneity at Multiple Spatial Scales. Sci. Total Environ. 2022, 838, 156509. [Google Scholar] [CrossRef]
- Sun, J.; Tummers, J.S.; Galib, S.M.; Lucas, M.C. Fish Community and Abundance Response to Improved Connectivity and More Natural Hydromorphology in a Post-Industrial Subcatchment. Sci. Total Environ. 2022, 802, 149720. [Google Scholar] [CrossRef]
- Hansen, H.H.; Schneider, M.; Hägele, T. A Habitat Connectivity Reality Check for Fish Physical Habitat Model Results and Decision-making for River Restoration. Ecol. Solut. Evid. 2023, 4, e12291. [Google Scholar] [CrossRef]
- Shaffer, J.A.; Munsch, S.; Juanes, F. Functional Diversity Responses of a Nearshore Fish Community to Restoration Driven by Large-Scale Dam Removal. Estuar. Coast. Shelf Sci. 2018, 213, 245–252. [Google Scholar] [CrossRef]
- Faucheux, N.M.; Miranda, L.E.; Taylor, J.M.; Farris, J. Impact of Dams on Stream Fish Diversity: A Different Result. Diversity 2023, 15, 728. [Google Scholar] [CrossRef]
- Pool, T.K.; Olden, J.D.; Whittier, J.B.; Paukert, C.P. Environmental Drivers of Fish Functional Diversity and Composition in the Lower Colorado River Basin. Can. J. Fish. Aquat. Sci. 2010, 67, 1791–1807. [Google Scholar] [CrossRef]
- Torres-Bejarano, A.M.; Sulliván, S.M.P.; González-Daza, W.; Cáceres, C.; Colorado, Z.G.J. Riparian Vegetation Structure and Seasonality Influence Functional Diversity More than Taxonomic Diversity of Stream Fish Assemblages in the Colombian Amazon. Aquat. Ecol. 2021, 56, 153–172. [Google Scholar] [CrossRef]
- Henriques-Silva, R.; Boivin, F.; Calcagno, V.; Urban, M.C.; Peres-Neto, P.R. On the Evolution of Dispersal via Heterogeneity in Spatial Connectivity. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142879. [Google Scholar] [CrossRef]
- Wolter, C.; Buijse, A.D.; Parasiewicz, P. Temporal and Spatial Patterns of Fish Response to Hydromorphological Processes. River Res. Appl. 2015, 32, 190–201. [Google Scholar] [CrossRef]
- Woolnough, D.A.; Downing, J.A.; Newton, T.J. Fish Movement and Habitat Use Depends on Water Body Size and Shape. Ecol. Freshw. Fish 2009, 18, 83–91. [Google Scholar] [CrossRef]
- Hale, R.; Colton, M.A.; Peng, P.; Swearer, S.E. Do Spatial Scale and Life History Affect Fish–Habitat Relationships? J. Anim. Ecol. 2019, 88, 439–449. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Ballew, N.G.; Kjelvik, M.K. Fish Behavioral Types and Their Ecological Consequences. Can. J. Fish. Aquat. Sci. 2014, 71, 927–944. [Google Scholar] [CrossRef]
- Magilligan, F.J.; Graber, B.E.; Nislow, K.H.; Chipman, J.W.; Sneddon, C.S.; Fox, C.A. River Restoration by Dam Removal: Enhancing Connectivity at Watershed Scales. Elem. Sci. Anth. 2016, 4, 000108. [Google Scholar] [CrossRef]
- Wenwu, M.; Fujiang, H.; Wei, W. Situation of Water Environment and Evaluation of Water Quality in Chishui River Basin. Acta Agric. Jiangxi 2021, 33, 87–91. [Google Scholar]
- Shao, X.; Fang, Y.; Jawitz, J.W.; Yan, J.; Cui, B. River Network Connectivity and Fish Diversity. Sci. Total Environ. 2019, 689, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.R.; Mallik, A.U. Species Diversity and Functional Diversity Relationship Varies with Disturbance Intensity. Ecosphere 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Hooper, D.U.; Solan, M.; Symstad, A.; Áz, S.D.; Gessner, M.O.; Buchmann, N.; Degrange, V.; Grime, P.; Hulot, F.; Mermillod-Blondin, F.; et al. Species Diversity, Functional Diversity, and Ecosystem Functioning. In Biodiversity and Ecosystem Functioning; Loreau, M., Naeem, S., Lnchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 195–208. [Google Scholar]
- Bowden, T.J. Modulation of the Immune System of Fish by Their Environment. Fish Shellfish Immunol. 2008, 25, 373–383. [Google Scholar] [CrossRef]
- Mamun, M.; An, K.-G. Key Factors Determining Water Quality, Fish Community Dynamics, and the Ecological Health in an Asian Temperate Lotic System. Ecol. Inform. 2022, 72, 101890. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Che, X.; Chen, X.; Tian, C.; Zhang, J. Impacts of Low-Order Stream Connectivity Restoration Projects on Aquatic Habitat and Fish Diversity. Fishes 2025, 10, 321. https://doi.org/10.3390/fishes10070321
Li X, Che X, Chen X, Tian C, Zhang J. Impacts of Low-Order Stream Connectivity Restoration Projects on Aquatic Habitat and Fish Diversity. Fishes. 2025; 10(7):321. https://doi.org/10.3390/fishes10070321
Chicago/Turabian StyleLi, Xinfeng, Xuan Che, Xiaolong Chen, Changfeng Tian, and Jiahua Zhang. 2025. "Impacts of Low-Order Stream Connectivity Restoration Projects on Aquatic Habitat and Fish Diversity" Fishes 10, no. 7: 321. https://doi.org/10.3390/fishes10070321
APA StyleLi, X., Che, X., Chen, X., Tian, C., & Zhang, J. (2025). Impacts of Low-Order Stream Connectivity Restoration Projects on Aquatic Habitat and Fish Diversity. Fishes, 10(7), 321. https://doi.org/10.3390/fishes10070321