Utilizing Environmental DNA for Early Monitoring of Non-Indigenous Fish Species in Maritime Ballast Water



Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Filtration
2.2. DNA Extraction and PCR Amplification
2.3. Sequencing and Sequence Analysis
2.4. Species Composition Analysis
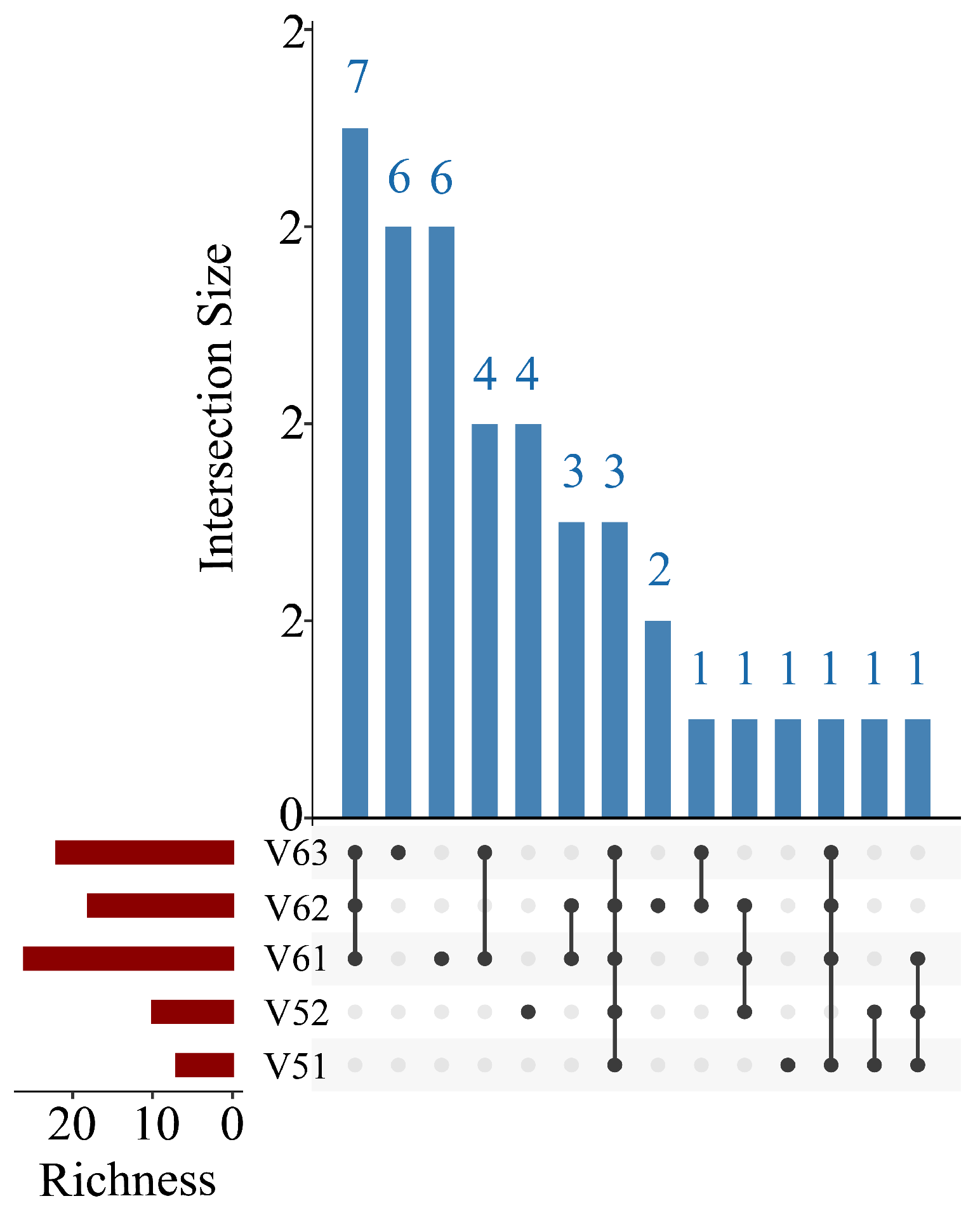
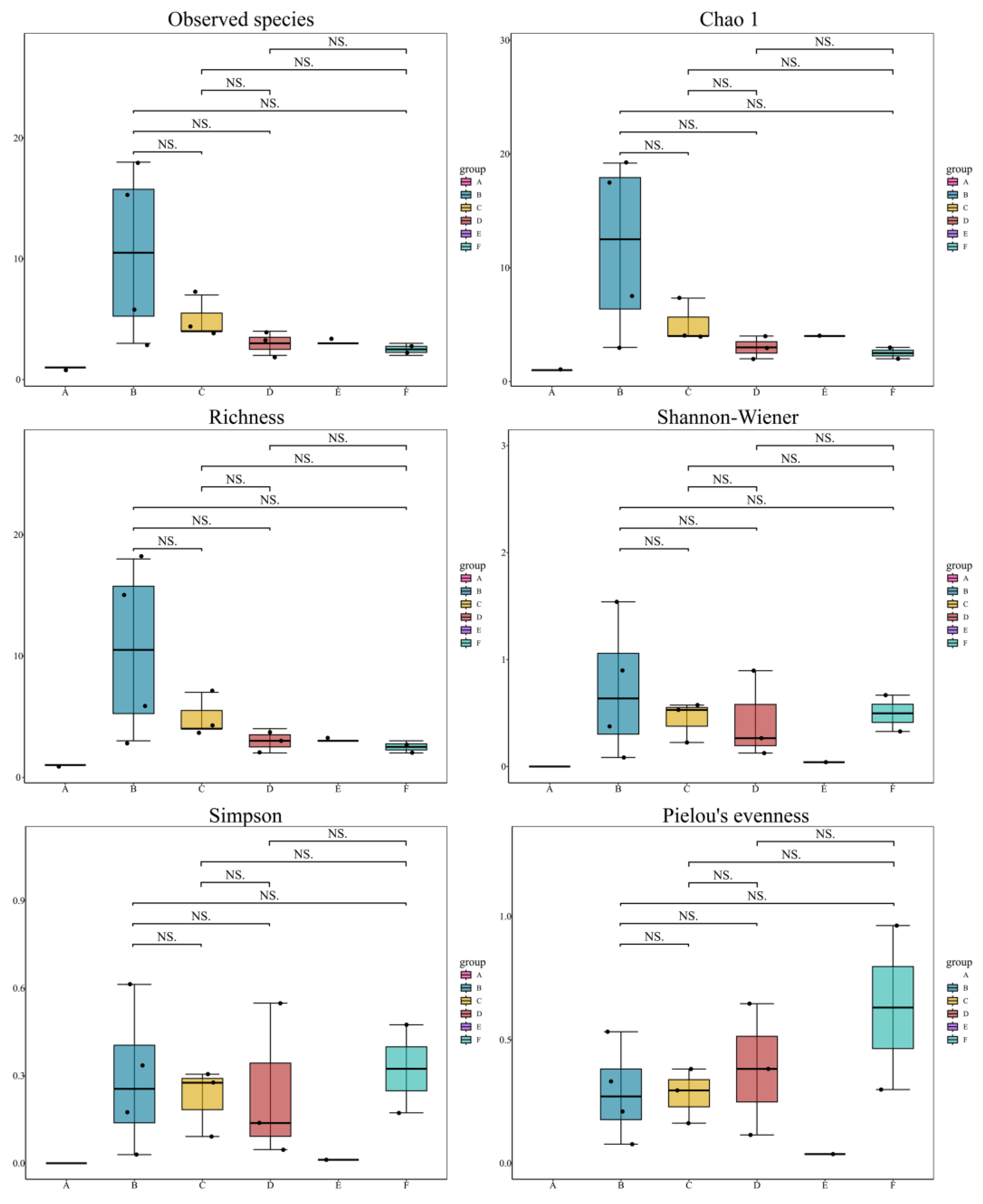
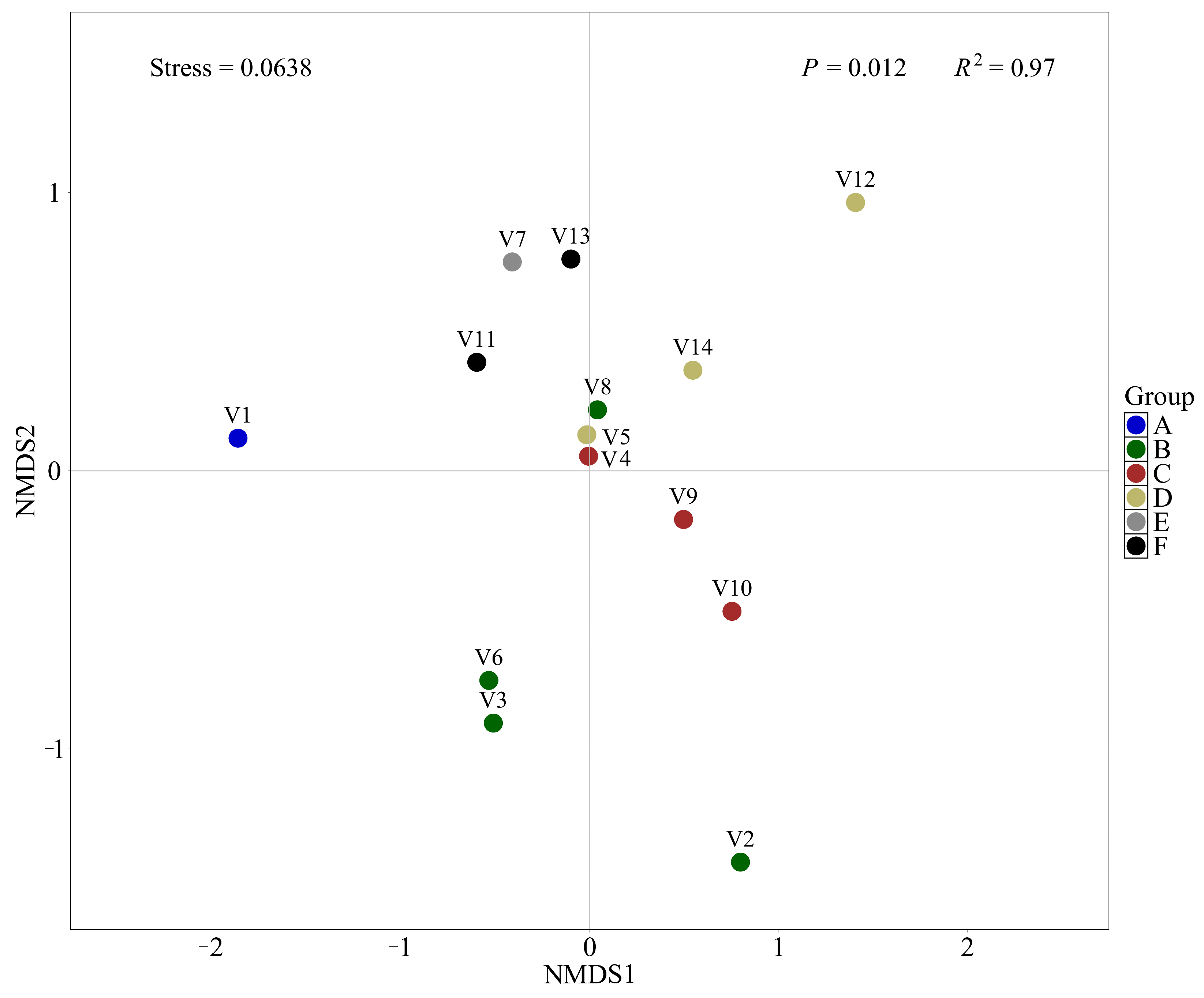
2.5. α- and β-Diversity Analysis
2.6. Non-Indigenous Fish Species Assessment
3. Results
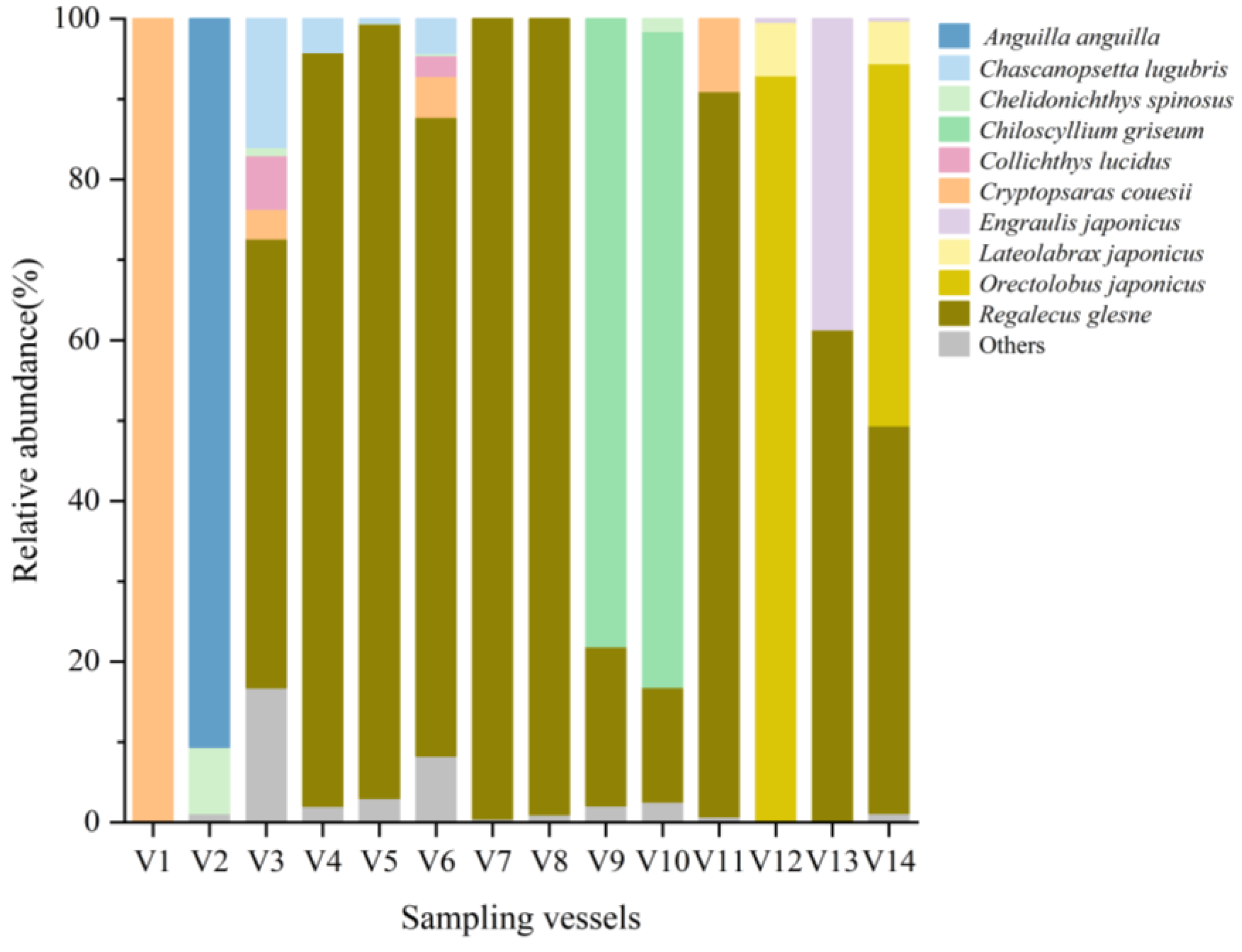
3.1. Fish Species Composition
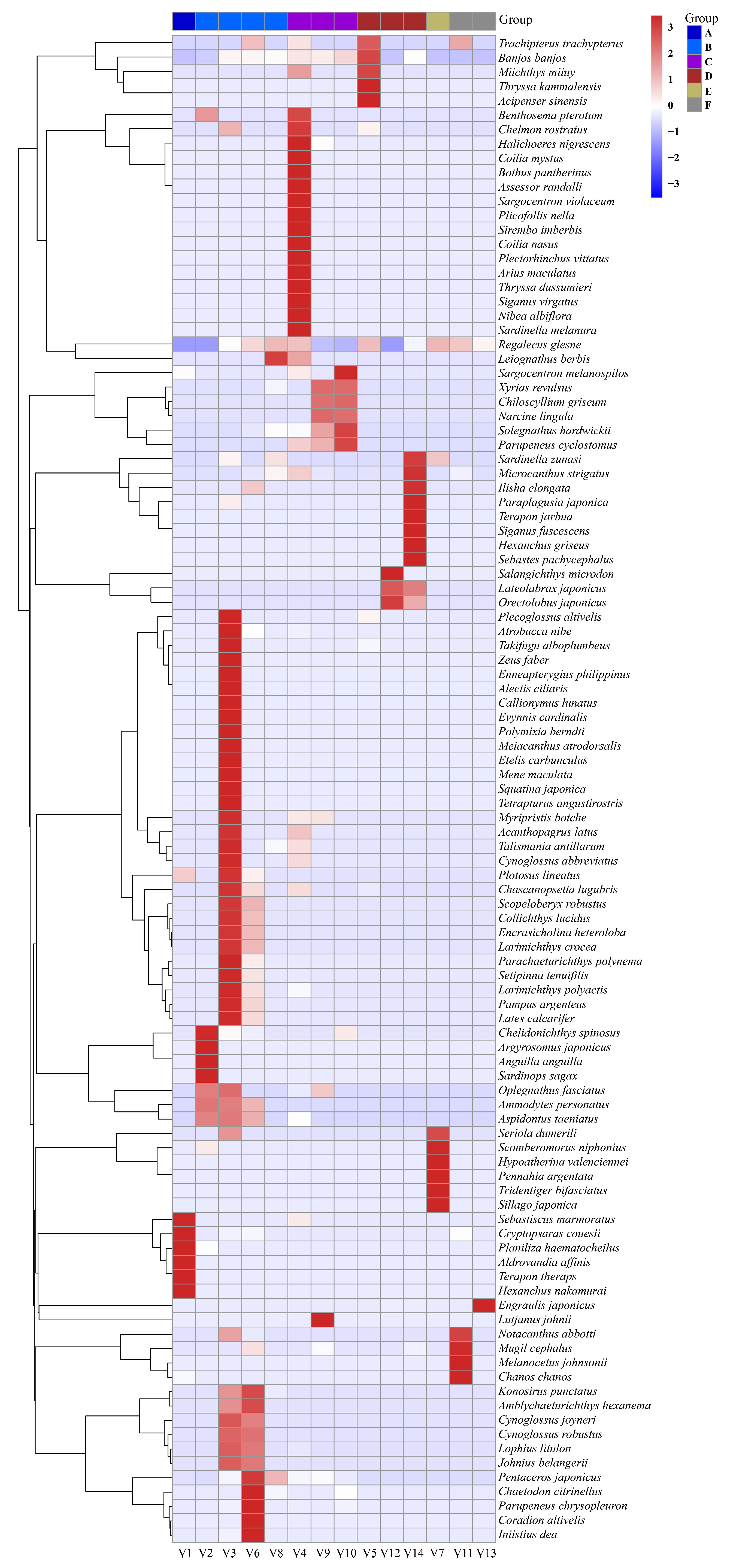
3.2. Results of Fish Diversity Analysis
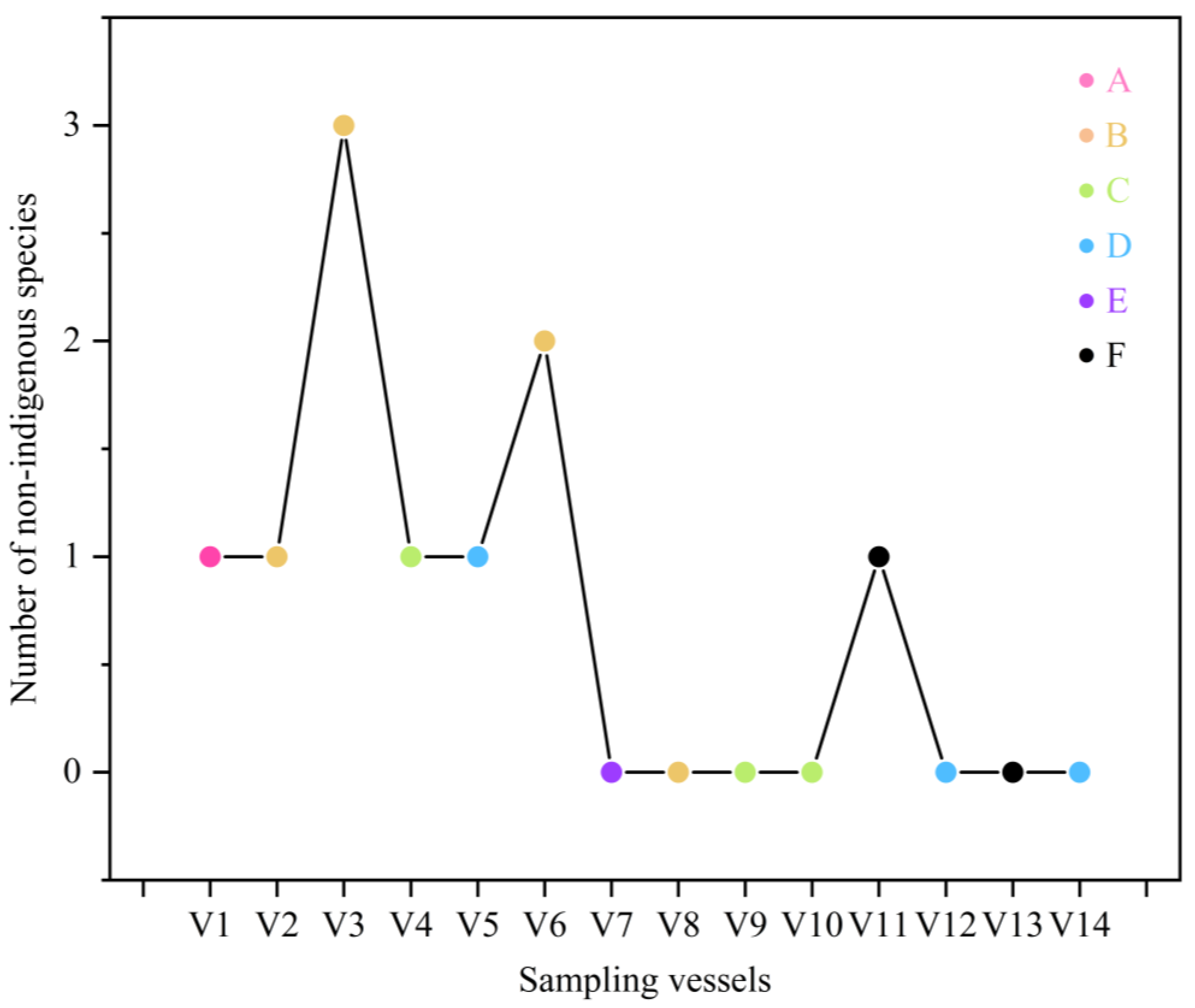
3.3. Non-Indigenous Fish Species Statistics
4. Discussion
4.1. Reliability and Effectiveness of eDNA Technology in Monitoring Non-Indigenous Fish Species
4.2. Distribution of Non-Indigenous Species and High-Risk Populations
4.3. Control Measures for Non-Indigenous Fish in Ballast Water
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salleh, N.A.; Rosli, F.N.; Akbar, M.A.; Yusof, A.; Sahrani, F.K.; Razak, S.A.; Ahmad, A.; Usup, G.; Bunawan, H. Pathogenic hitchhiker diversity on international ships’ ballast water at West Malaysia port. Mar. Pollut. Bull. 2021, 172, 112850. [Google Scholar] [CrossRef]
- Hwang, J.; Park, S.Y.; Lee, S.; Lee, T.K. High diversity and potential translocation of DNA viruses in ballast water. Mar. Pollut. Bull. 2018, 137, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Cope, R.C.; Prowse, T.A.A.; Ross, J.V.; Wittmann, T.A.; Cassey, P. Temporal modelling of ballast water discharge and ship-mediated invasion risk to Australia. R. Soc. Open Sci. 2015, 2, 150039. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, C.; Pienaar, R.N.; Sym, S.D. Possible introduction of alien phytoplankton via shipping ballast water: A South African perspective. S. Afr. J. Bot. 2001, 67, 465–474. [Google Scholar] [CrossRef]
- García-Garay, J.; Franco-Herrera, A.; Machuca-Martinez, F. Zooplankton sensitivity and phytoplankton regrowth for ballast water treatment with advanced oxidation processes. Environ. Sci. Pollut. Res. 2018, 25, 35008–35014. [Google Scholar] [CrossRef]
- Gavand, M.R.; McClintock, J.B.; Amsler, C.D.; Peters, R.W.; Angus, R.A. Effects of sonication and advanced chemical oxidants on the unicellular green alga Dunaliella tertiolecta and cysts, larvae and adults of the brine shrimp Artemia salina: A prospective treatment to eradicate invasive organisms from ballast water. Mar. Pollut. Bull. 2007, 54, 1777–1788. [Google Scholar] [CrossRef]
- Gollasch, S.; Lenz, J.; Dammer, M.; Andres, H.G. Survival of tropical ballast water organisms during a cruise from the Indian Ocean to the North Sea. J. Plankton Res. 2000, 22, 923–937. [Google Scholar] [CrossRef]
- Ardura, A.; Martinez, J.L.; Zaiko, A.; Garcia-Vazquez, E. Poorer diversity but tougher species in old ballast water: Biosecurity challenges explored from visual and molecular techniques. Mar. Pollut. Bull. 2021, 168, 112465. [Google Scholar] [CrossRef]
- Mandrak, N.E.; Cudmore, B. The fall of native fishes and the rise of non-native fishes in the Great Lakes Basin. Aquat. Ecosyst. Health 2010, 13, 255–268. [Google Scholar] [CrossRef]
- Inoue, K.; Odo, S.; Noda, T.; Nakao, S.; Takeyama, S.; Yamaha, E.; Yamazaki, F.; Harayama, S. A possible hybrid zone in the Mytilus edulis complex in Japan revealed by PCR markers. Mar. Biol. 1997, 128, 91–95. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Pattison, Z.; Taylor, N.G.; Verbrugge, L.; Diagne, C.; Ahmed, D.A.; Leroy, B.; Angulo, E.; Briski, E.; Capinha, C.; et al. Global economic costs of aquatic invasive alien species. Sci. Total Environ. 2021, 775, 145238. [Google Scholar] [CrossRef] [PubMed]
- Trebitz, A.S.; Hoffman, J.C.; Darling, J.A.; Pilgrim, E.M.; Kelly, J.R.; Brown, E.A.; Chadderton, W.L.; Egan, S.P.; Grey, E.K.; Hashsham, S.A.; et al. Early detection monitoring for aquatic non-indigenous species: Optimizing surveillance, incorporating advanced technologies, and identifying research needs. J. Environ. Manag. 2017, 202, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Global Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Scriver, M.; Marinich, A.; Wilson, C.; Freeland, J. Development of species-specific environmental DNA (eDNA) markers for invasive aquatic plants. Aquat. Bot. 2015, 122, 27–31. [Google Scholar] [CrossRef]
- Tsuji, S.; Takahara, T.; Doi, H.; Shibata, N.; Yamanaka, H. The detection of aquatic macroorganisms using environmental DNA analysis-A review of methods for collection, extraction, and detection. Environ. DNA 2019, 1, 99–108. [Google Scholar] [CrossRef]
- Jannel, L.A.; Guilhaumon, F.; Valade, P.; Chabanet, P.; Borie, G.; Grondin, H.; Jourand, P. eDNA metabarcoding, a promising tool for monitoring aquatic biodiversity in the estuaries of Reunion Island (South-West Indian Ocean). Environ. DNA 2024, 6, e70044. [Google Scholar] [CrossRef]
- Lymperopoulou, D.S.; Dobbs, F.C. Bacterial diversity in ships’ ballast water, ballast-water exchange, and implications for ship-mediated dispersal of microorganisms. Environ. Sci. Technol. 2017, 51, 1962–1972. [Google Scholar] [CrossRef]
- Gerhard, W.A.; Gunsch, C.K. Metabarcoding and machine learning analysis of environmental DNA in ballast water arriving to hub ports. Environ. Int. 2019, 124, 312–319. [Google Scholar] [CrossRef]
- Shang, L.; Hu, Z.; Deng, Y.; Li, J.; Liu, Y.; Song, X.; Zhai, X.Y.; Zhan, Z.F.; Tian, W.; Xu, J.X.; et al. Transoceanic ships as a source of alien dinoflagellate invasions of inland freshwater ecosystems. Harmful Algae 2024, 135, 102630. [Google Scholar] [CrossRef]
- Ghabooli, S.; Zhan, A.; Paolucci, E.; Hernandez, M.R.; Briski, E.; Cristescu, M.E.; MacIsaac, H.J. Population attenuation in zooplankton communities during transoceanic transfer in ballast water. Ecol. Evol. 2016, 6, 6170–6177. [Google Scholar] [CrossRef]
- Zaiko, A.; Martinez, J.L.; Schmidt-Petersen, J.; Ribicic, D.; Samuiloviene, A.; Garcia-Vazquez, E. Metabarcoding approach for the ballast water surveillance—An advantageous solution or an awkward challenge? Mar. Pollut. Bull. 2015, 92, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Stepien, C.A.; Neilson, M.E. What’s in a name? Taxonomy and nomenclature of invasive gobies in the Great Lakes and beyond. J. Great Lakes Res. 2013, 39, 555–559. [Google Scholar] [CrossRef]
- Yang, L. The role of new ports in port-related industries and urban development: A case study of Dongjiakou Port. J. Jinan Vocat. Coll. 2017, 95–96. [Google Scholar]
- Yu, Y.Q.; Zhang, B.M.; Teng, X.D.; Zhao, L. Surveillance on medical vectors at Qingdao Dongjiakou port. Chin. Front. Health Quar. 2017, 40, 173–176. [Google Scholar]
- Liu, J.W.; Yu, Z.X.; Zhu, C.B.; Hu, B.; Liu, Z. Determination of key factors for the moored operations of 400,000-DWT bulk carriers at Dongjiakou Port. Waterw. Eng. 2023, 72–77. [Google Scholar]
- Kumar, G.; Eble, J.E.; Gaither, M.R. A practical guide to sample preservation and pre-PCR processing of aquatic environmental DNA. Mol. Ecol. Resour. 2020, 20, 29–39. [Google Scholar] [CrossRef]
- IMO. International Convention for the Control and Management of Ship’s Ballast Water and Sediments; IMO: London, UK, 2004; Available online: https://www.imo.org/en/About/Conventions/Pages/International-Convention-for-the-Control-and-Management-of-Ships%27-Ballast-Water-and-Sediments-(BWM).aspx (accessed on 1 February 2024).
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef]
- Pagenkopp Lohan, K.M.; Fleischer, R.C.; Carney, K.J.; Holzer, K.K.; Ruiz, G.M. Amplicon-based pyrosequencing reveals high diversity of protistan parasites in ships’ ballast water: Implications for biogeography and infectious diseases. Microb. Ecol. 2016, 71, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y. The Checklist of Marine Biota of China Seas, 1st ed.; Science Press: Beijing, China, 2008. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 623–656. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Xiong, W.; Shen, C.; Wu, Z.; Lu, H.; Yan, Y. A brief overview of known introductions of non-native marine and coastal species into China. Aquat. Invasions 2017, 12, 109–115. [Google Scholar] [CrossRef]
- Zhao, S.J.; Zhang, X.J.; Li, C.D.; Zhu, A.Y.; Wu, C.W. Alien fishes of mariculture in China. Mar. Sci. 2006, 30, 75–80. [Google Scholar]
- Xu, H.; Qiang, S. China’s Invasive Alien Species, Revised Edition; Science Press: Beijing, China, 2018. [Google Scholar]
- Pusey, B.; Burrows, D.; Arthington, A.; Kennard, M. Translocation and spread of piscivorous fishes in the Burdekin River, north-eastern Australia. Biol. Invasions 2006, 8, 965–977. [Google Scholar] [CrossRef]
- Çinar, M.E. The alien ascidian Styela clava now invading the Sea of Marmara (Tunicata: Ascidiacea). ZooKeys 2016, 563, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Grammatiki, K.; de Jonge, N.; Nielsen, J.L.; Gómez-Gomez, S.C.; Avramidi, E.; Lymperaki, M.M.; Marcou, M.; Ioannou, G.; Papatheodoulou, M.; Dargent, O.; et al. eDNA metabarcoding of marine invertebrate communities at RO desalination plant outfalls in Cyprus. Mar. Pollut. Bull. 2025, 214, 117609. [Google Scholar] [CrossRef]
- Giroux, M.S.; Reichman, J.R.; Langknecht, T.; Burgess, R.M.; Ho, K.T. Environmental RNA as a tool for marine community biodiversity assessments. Sci. Rep. 2022, 12, 17782. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Davison, P.I.; Creach, V.; Stone, D.; Bass, D.; Tidbury, H.J. The application of eDNA for monitoring aquatic non-indigenous species: Practical and policy considerations. Diversity 2023, 15, 631. [Google Scholar] [CrossRef]
- Outinen, O.; Bailey, S.A.; Casas-Monroy, O.; Delacroix, S.; Gorgula, S.; Griniene, E.; Kakkonen, J.E.; Srebaliene, G. Biological testing of ships’ ballast water indicates challenges for the implementation of the Ballast Water Management Convention. Front. Mar. Sci. 2024, 11, 1334286. [Google Scholar] [CrossRef]
- Wonham, M.J.; Carlton, J.T.; Ruiz, G.M.; Smith, L.D. Fish and ships: Relating dispersal frequency to success in biological invasions. Mar. Biol. 2000, 136, 1111–1121. [Google Scholar] [CrossRef]
- Arai, K.; Itakura, H.; Yoneta, A.; Yoshinaga, T.; Shirotori, F.; Kaifu, K.; Kimura, S. Discovering the dominance of the non-native European eel in the upper reaches of the Tone River system, Japan. Fish. Sci. 2017, 83, 735–742. [Google Scholar] [CrossRef]
- Stern, N.; Rothman, S.B.S. An alarming mariculture breach in a coral reef: Alien barramundi Lates calcarifer (Bloch, 1790) at the northern Red Sea. BioInvasions Rec. 2021, 10, 181–187. [Google Scholar] [CrossRef]
- Buley, R.P.; Hasler, C.T.; Tix, J.A.; Suski, C.D.; Hubert, T.D. Can ozone be used to control the spread of freshwater Aquatic Invasive Species? Manag. Biol. Invasion 2017, 8, 13–24. [Google Scholar] [CrossRef]
- Lakshmi, E.; Priya, M.; Achari, V.S. An overview on the treatment of ballast water in ships. Ocean Coast. Manag. 2021, 199, 105296. [Google Scholar] [CrossRef]
- Tsolaki, E.; Diamadopoulos, E. Technologies for ballast water treatment: A review. J. Chem. Technol. Biot 2010, 85, 19–32. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Z.; Li, Y.; Fan, X. Fast detection technology for alien invasive fish in ship ballast water. J. Shandong Jiaotong Univ. 2023, 31, 110–118. [Google Scholar]
- Egan, S.P.; Grey, E.; Olds, B.; Feder, J.L.; Ruggiero, S.T.; Tanner, C.E.; Lodge, D.M. Rapid molecular detection of invasive species in ballast and harbor water by integrating environmental DNA and light transmission spectroscopy. Environ. Sci. Technol. 2015, 49, 4113–4121. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Miyata, K.; Yamane, M.; Honda, H. Environmental nucleic acid pollution: Characterization of wastewater generating false positives in molecular ecological surveys. ACS ES T Water 2023, 3, 756–764. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Grouping Type | Vessels | Ballast Tank Samples |
|---|---|---|---|
| A | Geographic grouping | V1 | - |
| B | Geographic grouping | V2, V3, V6, V8 | - |
| C | Geographic grouping | V4, V9, V10 | - |
| D | Geographic grouping | V5, V12, V14 | - |
| E | Geographic grouping | V7 | - |
| F | Geographic grouping | V11, V13 | - |
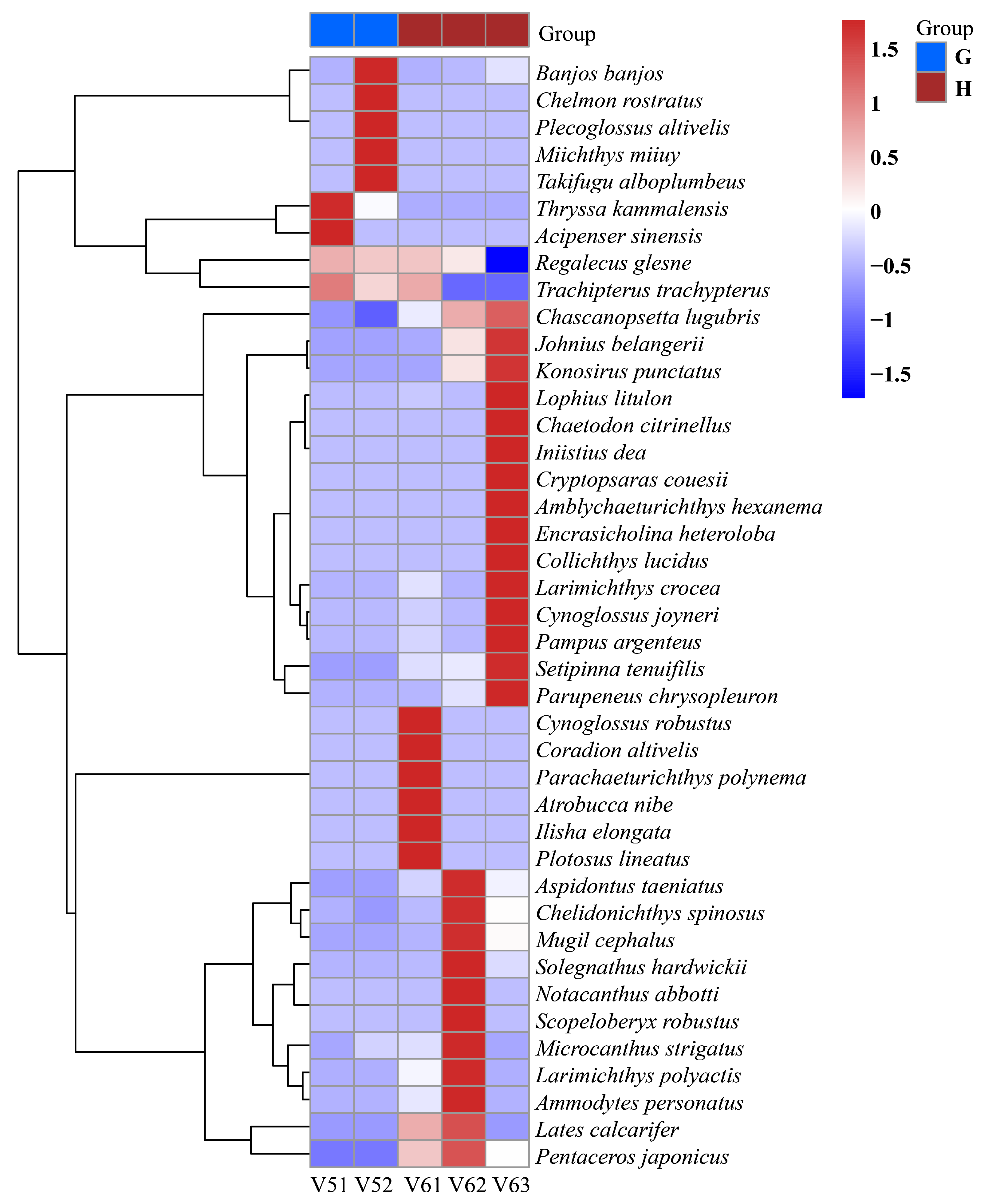
| G | Intra-vessel sampling | V5 | V51, V52 |
| H | Intra-vessel sampling | V6 | V61, V62, V63 |
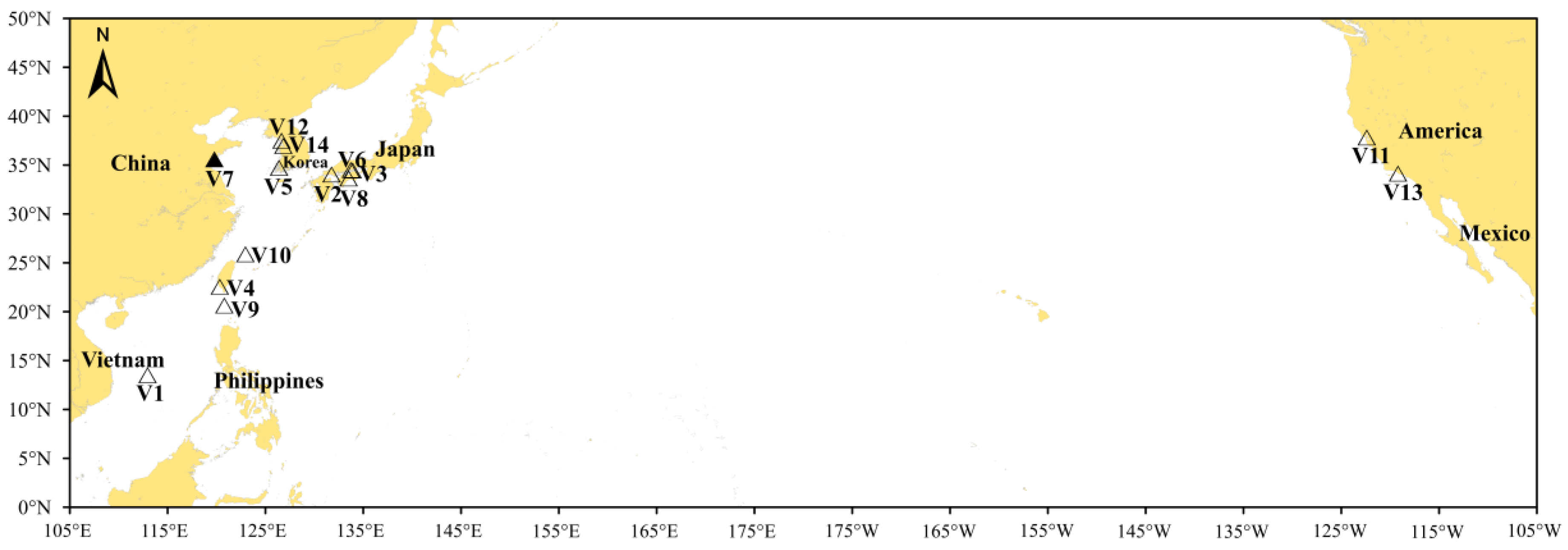
| Station | IMO Number | Vessel Name | Date of Uptake | Sampling Date | Ballast Tank Number and Sampling Volume | Location of Uptake | Sea Area 5 | Age of Ballast Water/Day 6 |
|---|---|---|---|---|---|---|---|---|
| V1 | 9735945 | Orion | 2.21 | 3.11 | BWT 1 (3 L) | 13.516° N, 112.955° E | Central South China Sea | 19 |
| V2 | 9485904 | Safeen al amal | 4.4 | 4.18 | BWT (3 L) | 34.050° N, 131.750° E | SetoNaikai | 14 |
| V3 | 9615042 | Friendly islands | 5.17 | 5.24 | DBT 2 (3 L) | 34.450° N, 133.927° E | SetoNaikai | 7 |
| V4 | 9146558 | Sea alice | 5.10 | 6.3 | BWT (3 L) | 22.562° N, 120.330° E | Taiwan Strait | 24 |
| V5 | 8747654 | Jin yang guan | 5.30 | 6.3 | FPT 3 (2 L), APT 4 (1 L) | 34.734° N, 126.379° E | South Yellow Sea | 4 |
| V6 | 9533335 | Mandarin river | 5.28 | 6.5 | FPT (1 L), APT (1 L), DBT (1 L) | 34.514° N, 133.737° E | SetoNaikai | 8 |
| V7 | 9362619 | Hong yang | 5.15 | 6.26 | BWT (3 L) | 35.6° N, 119.783° E | Langyatai Bay | 42 |
| V8 | 9815305 | Soc glory | 6.27 | 7.2 | BWT (3 L) | 33.643° N, 133.508° E | Tosa Bay | 5 |
| V9 | 9919723 | Theresa dua | 7.24 | 7.29 | BWT (3 L) | 20.65° N, 120.8° E | Northeastern South China Sea | 5 |
| V10 | 9483293 | Sg foundation | 8.19 | 8.26 | BWT (3 L) | 25.883° N, 122.96° E | Southern East China Sea | 7 |
| V11 | 9392377 | Argent sunrise | 8.25 | 9.19 | BWT (3 L) | 37.829° N, 122.393° W | San Francisco Bay | 25 |
| V12 | 9488396 | Xin zhong rui 7 | 9.8 | 11.1 | BWT (3 L) | 37.504° N, 126.637° E | Ganghwa Bay | 54 |
| V13 | 9288825 | Wenche victory | 10.20 | 11.18 | BWT (3 L) | 34.147° N, 119.211° W | Santa Barbara Channel | 29 |
| V14 | 1020851 | Express star | 11.19 | 11.23 | BWT (3 L) | 36.962° N, 126.838° E | Pyeongtaek Bay | 4 |
| Sample | Chao1 | Shannon Index | Simpson Index | Pielou Evenness |
|---|---|---|---|---|
| V51 | 7 | 0.104679399 | 0.034140565 | 0.053794569 |
| V52 | 10 | 0.214499753 | 0.097928275 | 0.093156059 |
| V61 | 36 | 0.279477463 | 0.092236373 | 0.085779368 |
| V62 | 18 | 0.489592095 | 0.176108518 | 0.16938724 |
| V63 | 22 | 1.576761115 | 0.62564492 | 0.510106587 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Jia, H.; Zhang, H. Utilizing Environmental DNA for Early Monitoring of Non-Indigenous Fish Species in Maritime Ballast Water. Fishes 2025, 10, 241. https://doi.org/10.3390/fishes10050241
Li H, Jia H, Zhang H. Utilizing Environmental DNA for Early Monitoring of Non-Indigenous Fish Species in Maritime Ballast Water. Fishes. 2025; 10(5):241. https://doi.org/10.3390/fishes10050241
Chicago/Turabian StyleLi, Hanglei, Hui Jia, and Hui Zhang. 2025. "Utilizing Environmental DNA for Early Monitoring of Non-Indigenous Fish Species in Maritime Ballast Water" Fishes 10, no. 5: 241. https://doi.org/10.3390/fishes10050241
APA StyleLi, H., Jia, H., & Zhang, H. (2025). Utilizing Environmental DNA for Early Monitoring of Non-Indigenous Fish Species in Maritime Ballast Water. Fishes, 10(5), 241. https://doi.org/10.3390/fishes10050241