Abstract
The mercury (Hg) present in ocean fish has caused concern regarding the effects of maternal consumption on child outcomes but it is now recognized that mothers that eat more ocean fish during pregnancy have children with higher social, scholastic, and IQ scores. These findings coincide with the current understanding of the mechanism of Hg toxicity which indicates ocean fish consumption will prevent rather than cause harm. High-Hg exposures sequester selenium (Se) and inhibit Se-dependent enzymes that prevent and reverse oxidative damage in the brain and support fetal brain development. However, aside from certain types of shark and other apex marine predators, seafood contains more Se than Hg and thus counteracts instead of contributing to Hg toxicity. This study evaluates the Hg and Se present in bigeye tuna, yellowfin tuna, albacore tuna, skipjack, swordfish, striped marlin, blue marlin, spearfish, mahimahi, wahoo, monchong, escolar, mako shark, and thresher shark to establish their health benefit values (HBVs). Positive HBVs (0.3–19.6), indicating a molar excess of Se over Hg, were found in 14 of the 15 species studied. Only mako shark uniformly contained Hg in excess of Se to produce a negative HBV (−16.4), indicating its consumption should be minimized during pregnancy.
Key Contribution:
Mercury (Hg) binds tissue selenium (Se), and although both are present in ocean fish, Se tends to be more abundant and improves the consumer’s Se status, thus preventing rather than causing Hg toxicity. Health benefit values provide a simple, easily understood, and reliable index for differentiating safe and beneficial ocean fish from marine mammals, sharks, and other foods which may not provide these benefits.
1. Introduction
Ocean fish are rich sources of nutrients required for fetal brain development including selenium (Se), iron, iodine, zinc, vitamins A, D, B12, and omega-3 fatty acids, particularly eicosapentaenoic acid and docosahexaenoic acid [1,2,3]. To optimize maternal and fetal health, women are advised to eat at least two servings of ocean fish per week during pregnancy and three servings per week during lactation [4]. However, fish contain methylmercury (CH3Hg+) which is neurotoxic at high exposures [5,6,7]. This has caused misunderstandings and confusion regarding the safety of eating ocean fish.
Concerns regarding maternal Hg exposures originated following poisoning incidents from eating fish from a Hg polluted bay in Minamata Japan in the 1950s [8,9], and bread made with Hg-treated wheat in Iraq in the 1950s and 1960s [10]. Toxic consequences appeared in adults with blood Hg levels above 1 µM and severe consequences were evident at blood levels above 2.5 µM, but unborn children were far more vulnerable than adults. Severe consequences were seen in children whose mothers were unaffected by their Hg exposures. Based on this, it was considered possible that low level Hg exposures during pregnancy might harm the developing fetal brain. This was the basis for initiating studies in populations with high seafood intakes, with seemingly conflicting results.
The Hg in fish is predominantly in the form of CH3Hg-Cys [11]. The population studied in the Seychelles eats 10–20 times more fish than is typical in the United States, but no adverse effects have been seen in association with their CH3Hg exposures [12,13,14]. The study performed in the Faroe Islands reported subtle effects in children associated with their mother’s high Hg and cadmium (Cd) exposures. Since Hg and Cd both bind Se, eating whale meat diminishes maternal Se availability. Even though far more ocean fish was consumed, over 90% of total Hg exposure in the Faroes originated from eating pilot whale blubber, muscle and organs. Less than 10% of Hg exposure came from eating ocean fish and although there were no clinically definable harms, uncertainties regarding potential risks resulted in federal and state agencies recommending that women limit ocean fish consumption to no more than two meals (~340 g/week) during pregnancy to ensure their Hg exposures were below the U.S. Food and Drug Administration (FDA) and U.S. Environmental Protection Agency (EPA) reference dose (RfD) of 0.1 µg Hg/kg/day [4]. The advice to limit maternal seafood Hg+ exposures was precautionary, and no adverse effects were expected to accompany maternal compliance. Only later was it recognized that reducing maternal seafood consumption would restrict their intakes of nutrients that are important for fetal brain development and potentially reduce their children’s social, scholastic, and IQ abilities [2,15].
A large (11,875 mother–child pairs) study of maternal fish consumption in the United Kingdom [2] indicated that substantial benefits were associated with increasing maternal fish consumption. Children of mothers who avoided ocean fish or ate amounts that complied with the recommended two-meal maximum had an increased risk of scoring in the lowest quartile for verbal intellectual quotients (IQ), compared to children of mothers exceeding the recommended fish intake. Children of mothers that avoided fish consumption during pregnancy showed loss of developmental benefits of a magnitude estimated to be 60 times greater than the worst-case effects associated with the highest pilot whale consumption in the Faroes. However, due to the subtle effects that accompanied high exposures to Hg, Cd, PCBs, and other bioaccumulative toxicants from eating pilot whale [16], widespread misunderstandings of the advisory have resulted in substantial benefits being lost to children whose mothers stopped eating ocean fish [17].
Assessments of fish consumption that considered the net effects from both fish nutrients and toxicants performed by the FDA [15] and the World Health Organization/Food Agriculture Organization (WHO/FAO) [18] established that children benefit when their mothers regularly consume fish during pregnancy, including those who eat well above the advisory limit of 340 g/week. These benefits include IQ gains of 2–5 points or more as well as improved social and scholastic abilities. Recently, a review of 30 seafood consumption studies involving ~212,000 mother–child pairs in the United States, six European countries, the Republic of Seychelles, Japan, and China included fish consumption rates that included groups eating over 2.8 kg per week [15]. Twenty-four of these studies reported a total of 52 beneficial outcomes. Two studies reported a single adverse outcome, but these were inconsistent and were not observed in any of the other studies.
Advice from the FDA and EPA currently recommends consuming at least two low-Hg-containing ocean fish meals per week during pregnancy [4]. Although the majority of ocean fish are safe and beneficial to eat, mistaken attribution of effects reported from the Faroes study resulted in pregnant women often being advised to avoid or limit their ocean fish consumption to levels below the recommended two or three fish meals per week [17,19]. Since the Se from ocean fish eaten by mothers in the Faroes prevented high Hg and Cd in pilot whale meat from harming their children [16,20], and eating ocean fish has substantial benefits [15]; avoiding them during pregnancy is unwise and unwarranted.
1.1. Paradigm Change and Advances in Understanding Seafood Safety and Benefits
Selenocysteine (Sec), the 21st genetically encoded amino acid, is the direct catalytic mediator in the active sites of enzymes (selenoenzymes) expressed in all vertebrate cells where they perform various elite functions including preventing and reversing oxidative damage [21]. Antioxidant thiomolecules are maintained in their reduced state and/or function in concert with selenoenzymes. The three thioredoxin reductases and five glutathione peroxidases are among the many selenoenzymes that control the intracellular redox state and prevent/reverse oxidative damage to cellular biomolecules. Their antioxidant activities are pivotally important in brain and endocrine tissues, but also make them vulnerable to inhibition by Hg and other soft electrophiles. Metallic and organic electrophiles accumulate in tissues due to aspects of their behavior at the molecular level, but the most prominent contributor is the nature of their electronic charges. Electrophiles readily bind with nucleophiles such as the thiolate of cysteine (Cys), particularly since thiomolecules are present in high molar abundance (~300 mM) in cells. This forms the CH3Hg-Cys which comprises the majority of Hg in fish tissue [11,22].
Methylmercury in fish is almost exclusively CH3Hg-Cys, with kinetically active CH3Hg+ readily exchanging covalent associations among thermodynamically equivalent thiols [6]. These include glutathione, thioredoxin, and other selenoenzyme substrates. Since the thiols of these molecules directly interact with selenoenzymes, they can function as suicide substrates that deliver bound CH3Hg+ into the enzyme’s active site in the proper orientation to place it in proximity with Sec. As the most potent intracellular nucleophile, Sec acquires CH3Hg+ from Cys to form CH3Hg-Sec, irreversibly inhibiting the enzyme. Due to the high affinity between Hg and Se, this bond permanently inhibits the enzyme’s activity. Therefore, by biochemical definition, CH3Hg+ is a highly specific irreversible selenoenzyme inhibitor.
The covalent HgSe bond that forms is highly stable and persists for the duration of Hg’s residence in the organism and beyond excretion [23]. Because the HgSe complex renders Se unavailable for subsequent cycles of Sec synthesis [23,24], high Hg-exposures (e.g., poisoning events or long-term exposures resulting in levels exceeding tissue Se present at ~1 µM) can gradually induce a conditioned Se-deficiency. If not offset by dietary Se intakes, high Hg exposures have the potential to prevent selenoenzyme synthesis and activities in brain and endocrine tissues. However, if dietary Se intakes are sufficient to maintain adequate Se availability to support selenoenzyme synthesis and activities, the pathological effects associated with Hg toxicity will not occur [21,25].
1.2. Health Benefit Values
The health benefit value (HBV) was created [26] to establish a reliably accurate criterion for differentiating fish and seafoods that are beneficial to consume during pregnancy from those which are less beneficial or potentially harmful. The HBV distinguishes food sources that supply more Se than Hg and therefore protect against Hg toxicity from those that contain more Hg than Se. Positive HBVs conservatively indicate the amount of bioavailable Se the food will provide [16]. Since additional Se from other foods will also protect against Hg exposure from fish or seafoods with negative HBVs, this criterion is solely a measure of Hg and Se intakes from the specific food source. The additional influence of beneficial nutrients present in the fish is not incorporated in this equation, nor is the dietary Se acquired from other sources, although these would augment the predicted beneficial health effects still further.
Likewise, the Hg, Cd, and other metallic or organic electrophile contaminants present in foods with negative HBVs would contribute to Se sequestration and pose the risk of inducing a conditioned Se deficiency [27], especially in populations that are already Se deficient. Due to their smaller sizes and shorter lives, the accumulation of these other electrophiles do not achieve meaningful levels in most ocean fish, but is an important consideration when assessing risks to populations that consistently consume apex predatory species of shark, certain marine mammals, or freshwater fish originating from watersheds with high point source contributions of metallic or organic electrophiles. To protect against the consequences of eating large amounts of foods with negative HBVs, it is necessary to also eat Se-rich foods such as ocean fish. It should be noted that although the role of Se from ocean fish was not known at the time [28], the authors of the Faroes Study were among the first to recognize that the fish eaten by mothers in the Faroes had provided nutritional benefits which counteracted the adverse effects otherwise expected from exposure to contaminants present in pilot whale blubber, meat, and organs [29].
This study assesses 15 varieties of ocean fish from the north central Pacific Ocean near Hawaii to establish their Hg, Se, HBVs and the formation of HgSe in relation to body weight and evaluates the relative safety and benefits expected in association with their consumption during pregnancy.
2. Materials and Methods
The data evaluated in this study were obtained from samples collected in 2006 from commercial landings of pelagic fish (see Table 1, Table 2 and Table 3) as reported in Kaneko and Ralston [30]. The original raw data is included as Supplementary Information.

Table 1.
Ocean fish species examined in this study.
Table 2.
Fish species, body weights, Hg contents, Se contents, and HBVs 1.
Table 3.
Tissue Hg and Se in relation to body weight, and Se in relation to Hg contents 1.
Briefly, ~100 g samples were obtained from market size classes representative of the sizes which are commonly consumed. Samples were weighed and stored frozen in trace-metal-free plastic bags and chain of custody was maintained from collection until analysis. Sample aliquots of ~0.4 g (wet weight) weighed to 0.0001 g from each sample were transferred into single use trace-element-free 50 mL digestion tubes (Environmental Express, Mt Pleasant, SC, USA) with every tenth fish sample being prepared in duplicate and with elemental spike recovery samples being performed accompanying each batch.
Each digestion batch was processed with blank and certified standard reference materials (dogfish muscle certified reference material DORM-2, National Research Council of Canada, Ottawa, ON, Canada) reporting certified concentrations of 4.64 ± 0.26 mg Hg/kg, 1.40 ± 0.09 mg Se/kg. Samples were treated with 5 mL of 16 N nitric acid (Fisher Trace Metal Grade, (Fisher Scientific, Pittsburgh, PA, USA)) and heated at 85 °C for 24 h in loosely capped tubes in deep-cell hot blocks (Environmental Express, Mt Pleasant, SC, USA). Samples were cooled for ~1 h. and 1.5 mL of 30% H2O2 (Fisher Certified A.C.S., Fisher Scientific, www.fishersci.com) was added. Samples were recapped and returned to heating in the dry block at 85 °C. Samples were cooled and 15 mL of 12 N HCl (Fisher Trace Metal Grade, Fisher Scientific, www.fishersci.com) was added. Samples were heated at 90 °C for 90 min to reduce Se-VI to Se-IV, then cooled, diluted to 50 mL with double distilled water.
Samples were analyzed for Hg by cold vapor atomic absorption spectrophotometry using a CETAC M-6000A (CETAC Technologies Inc., Omaha, NE, USA). Selenium was analyzed by hydride generation atomic fluorescence spectroscopy using a PS Analytical Dual Millennium Excalibur, (PS Analytical, Deerfield Beach, FL, USA).
The Hg and Se molar concentrations of these samples were evaluated in relation to body weights. Individual fish tissue Hg and Se concentrations were used to calculate HBVs [31] as shown in Equation (1).
HBV = [(Se − Hg)/Se] • (Se + Hg)
The original HBV equation applied in an earlier evaluation of this data [30] was updated [32] to better reflect Hg and Se molar relationships to health risks vs. potential benefits of seafood and fish consumption. To assess formation and accumulation of HgSe complexes in these fish, Se contents were plotted in relation to Hg contents.
Data Treatment and Statistical Analysis
Mercury and Se concentrations of digestion blanks prepared with each batch were subtracted from analysis results and elemental recoveries in certified standard reference materials were evaluated to ensure observed values were within the certified limits and spike recoveries were within the 100 ± 5% range before batches were considered qualified for inclusion in the database. Means, standard deviations, regression analyses, and data outlier assessments were performed using the Excel Data Analysis ToolPac. Version 2501.
3. Results
Elemental analysis data for individual fish from the 15 species assessed were plotted in relation to their body weights, as shown in Figure 1 (Hg) and Figure 2 (Se). Health benefit values are plotted in Figure 3, and correlations between body weight and elemental concentrations or HBVs are shown in Table 2 and Table 3.
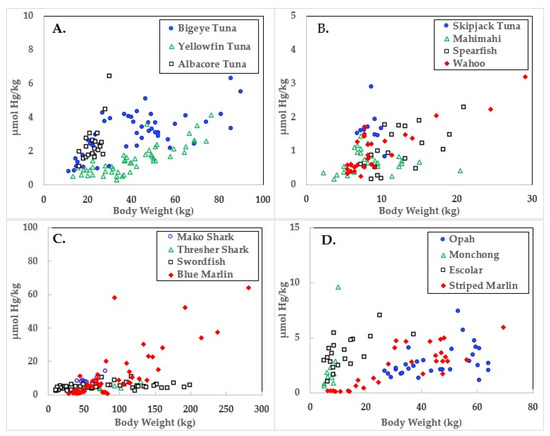
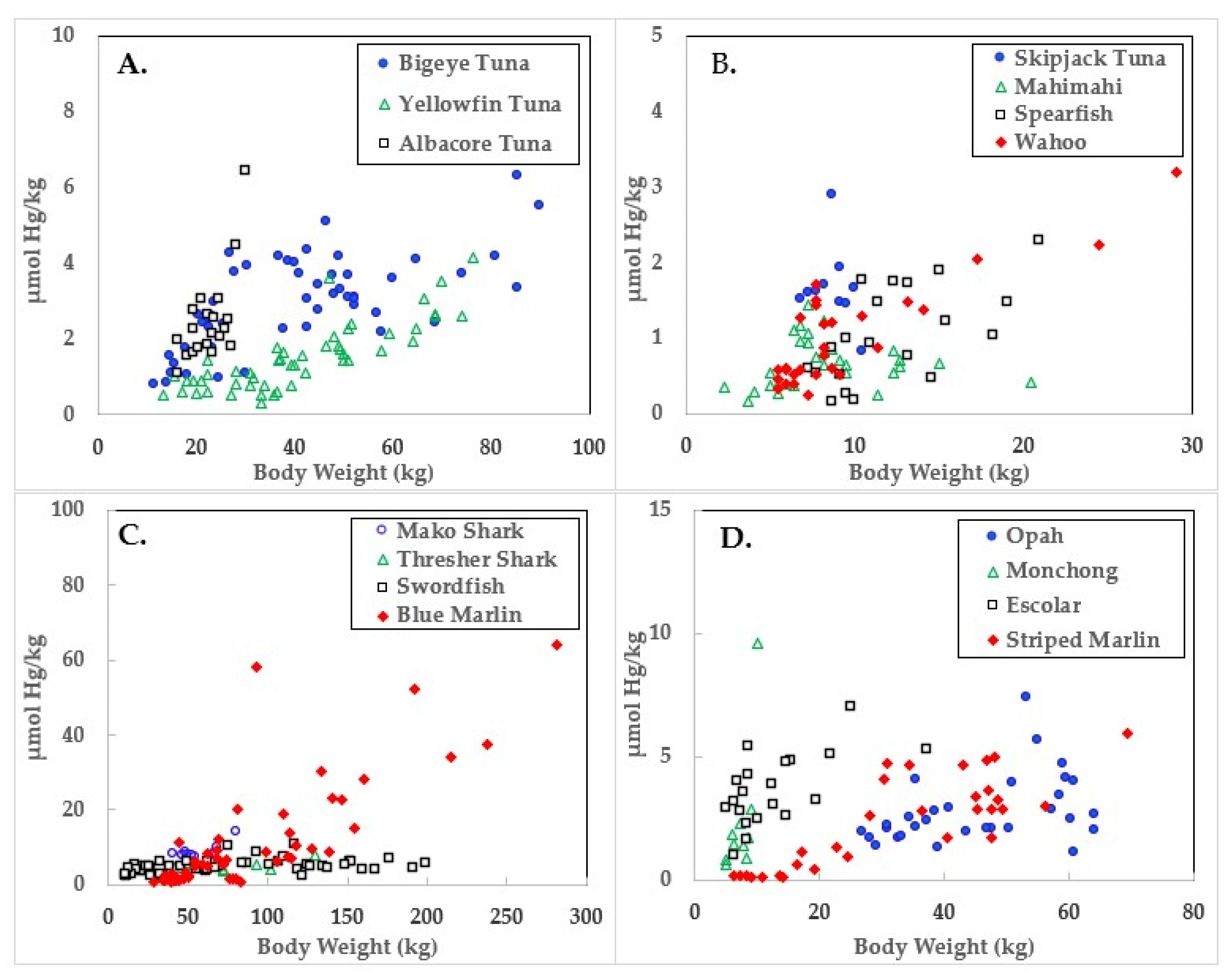
Figure 1.
Mercury concentrations in ocean fish plotted in relation to body weights.
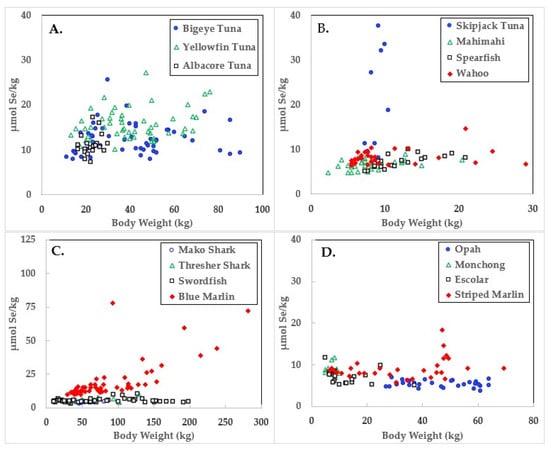
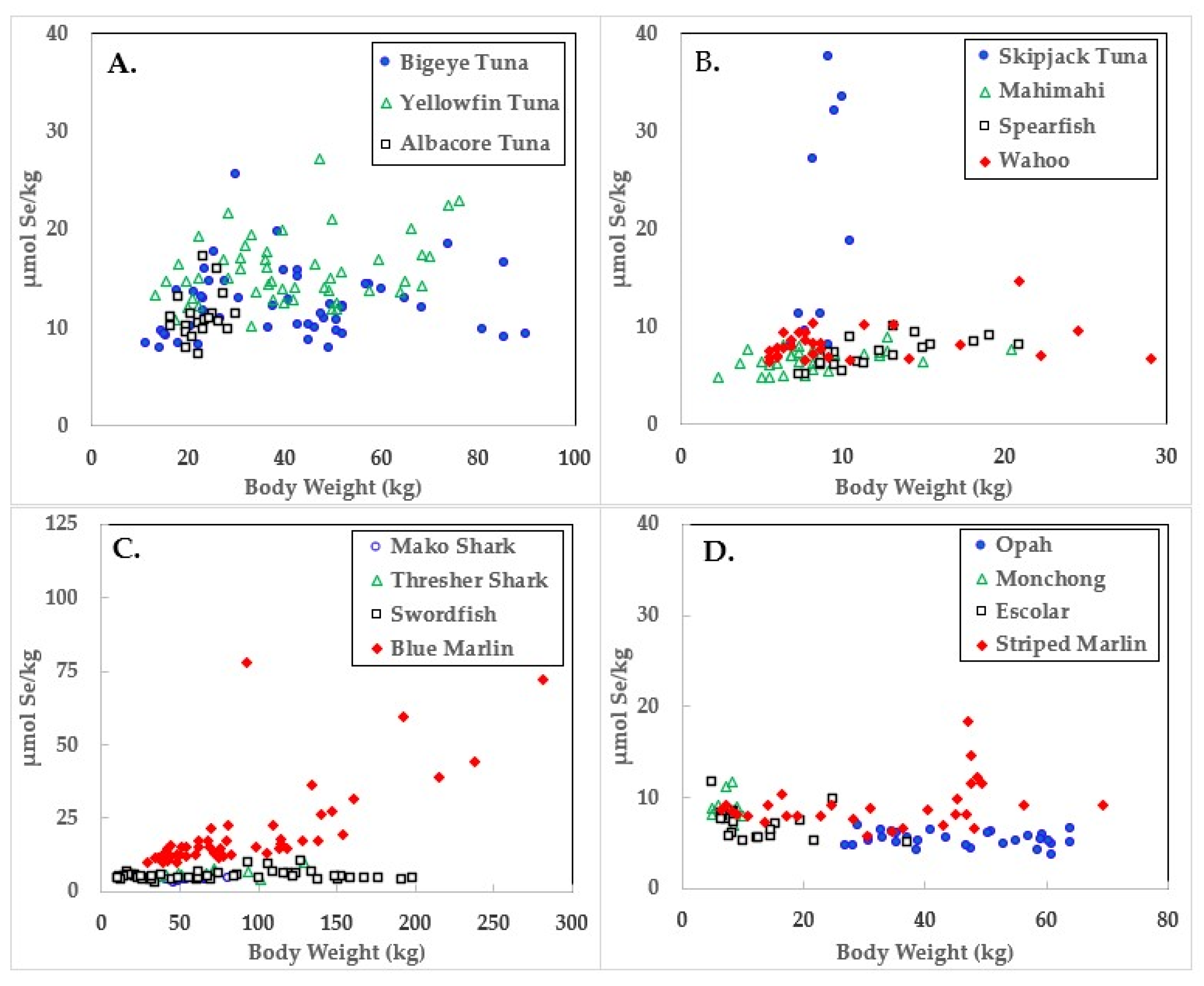
Figure 2.
Selenium concentrations in ocean fish plotted in relation to body weights.
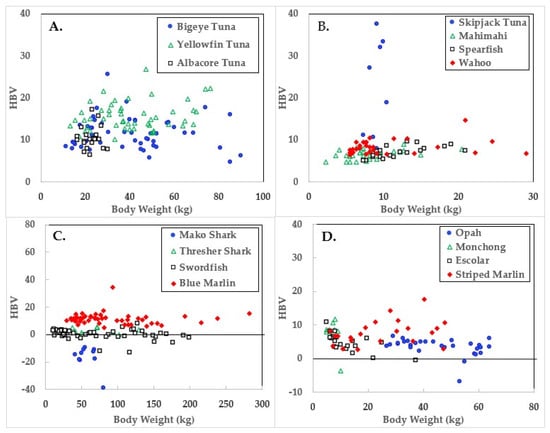
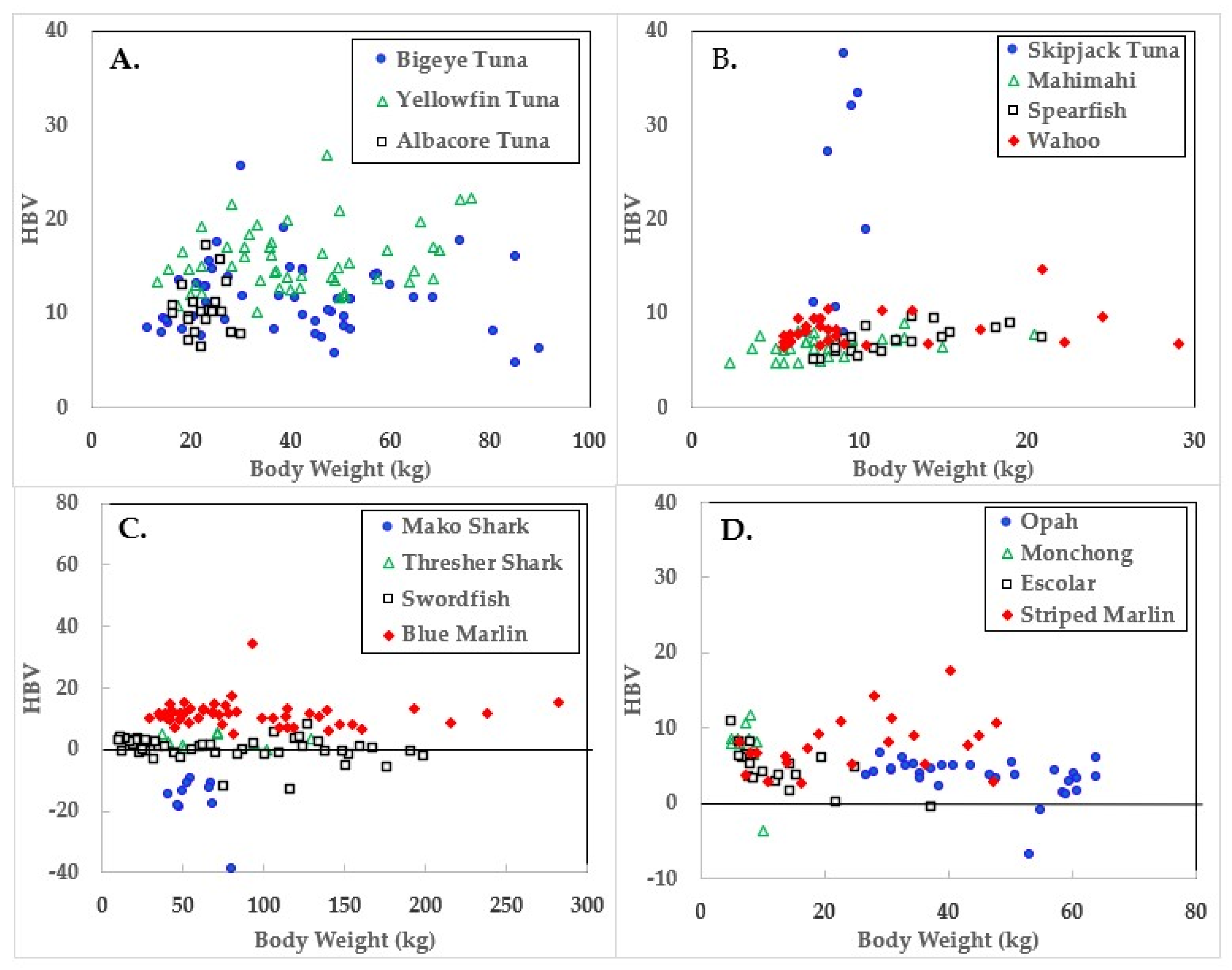
Figure 3.
Health benefit values (HBV) of ocean fish plotted in relation to body weights.
Data for the three large tuna varieties; bigeye, yellowfin, and albacore are shown in panel A of Figure 1, Figure 2 and Figure 3. Mercury concentrations are uniformly lower than Se for these fish, resulting in some of the highest HBVs observed in this study. Bigeye tuna had higher Hg contents than the other two forms, but Se contents were also high, resulting in HBVs which were uniformly positive and between those of yellowfin and albacore.
Due to its lower weight range, skipjack tuna was graphed with mahimahi, spearfish, and wahoo, as shown in panel B of Figure 1, Figure 2 and Figure 3. Skipjack had the highest HBVs observed in this study. The HBVs of the four tuna species averaged 14.2 ± 4.2, indicating their consumption is a particularly valuable source of dietary Se. Mahimahi, spearfish, and wahoo were low in Hg but rich in Se, resulting in uniformly positive HBVs (see Table 2).
Mako shark, thresher shark, swordfish, and blue marlin Hg results are shown in panel C of Figure 1, Figure 2 and Figure 3. Mako shark HBVs were uniformly negative and progressively more negative in direct association with their body weights. In contrast, the Se contents of the thresher sharks were consistently greater than those of Hg and all samples had positive HBVs. Smaller swordfish had more Se than Hg in their tissues, but larger specimens contained more Hg, resulting in some having negative HBVs. As a result, their average HBV was near zero. One swordfish sample was entirely excluded from the data set as an analytical outlier (21.6 µmol Hg/kg), a value ~9 SD higher than the group mean. No reasons were evident for this outlier, but it was confirmed by reanalysis. Opah, monchong, and escolar samples included in this study generally but not uniformly contained Se in excess of Hg, as seen in panel D of Figure 3, and had positive HBVs as a result (Table 2).
Mean body weights, Hg concentrations, relationships to body weights, and calculated HBVs are shown in Table 2. Although mako shark did not have the highest Hg contents or rate of bioaccumulation, it was the only species which consistently contained Hg in molar excess of Se, resulting in a negative HBV.
As seen in Figure 1 and Table 2, the accumulation of Hg in ocean fish tended to be directly proportional to body weight, but species-dependent differences were evident. In contrast, Figure 2 and Table 3 show there was no weight-dependent increase in Se for many species, possibly indicative of the homeostatic control mechanisms characteristic of Se physiology in all vertebrates.
Fish HBVs were stable in many ocean fish types regardless of body weight, but spearfish showed an increase with size (y =0.232x + 4.275, p < 0.01). Meanwhile escolar showed a downward trend with increasing weight (y = −0.225 + 7.706, p < 0.001) and non-significant downward trends of HBV decreasing in relation to increasing body weight were evident in swordfish (p = 0.07) and yellowfin tuna (p = 0.08). Mako shark HBVs were uniformly negative, but no trends were evident, possibly due to the low sample number and lack of heterogeneity in body weights.
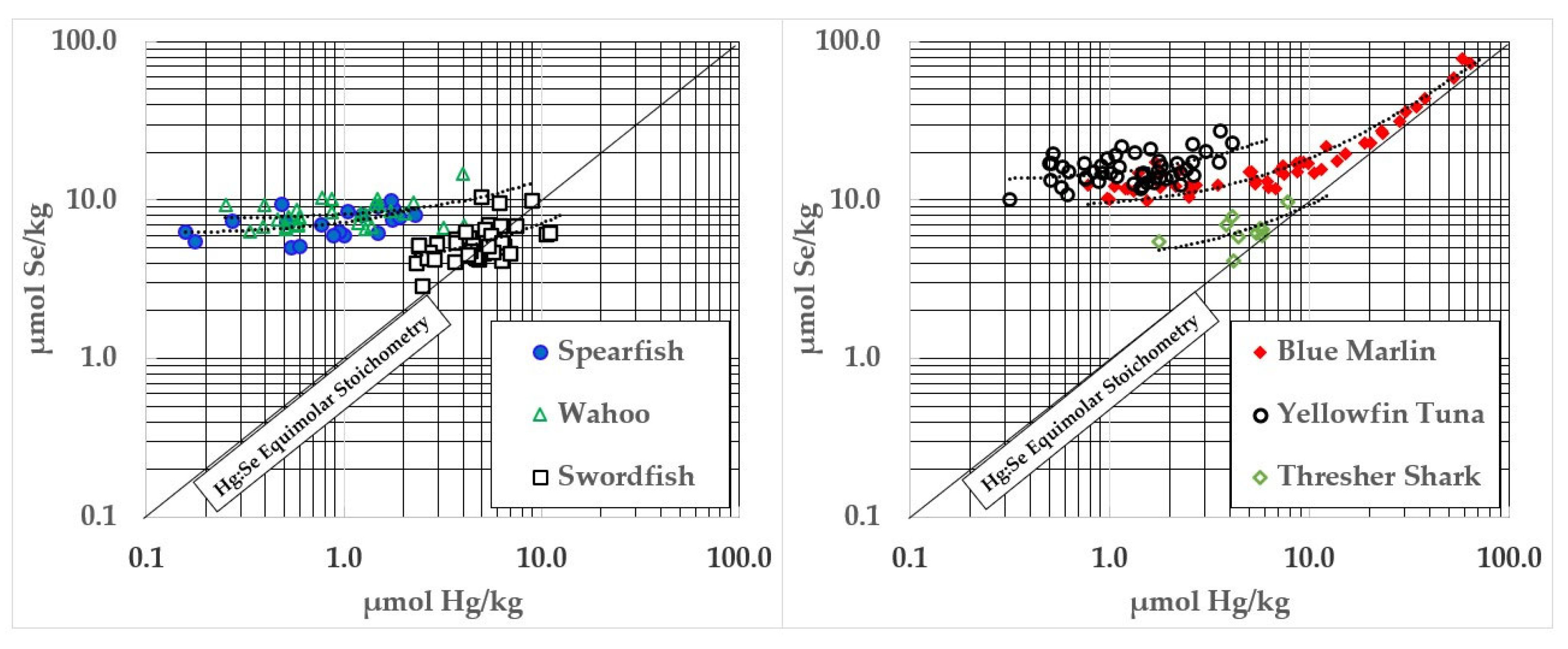
Several fish species demonstrate a statistically significant coaccumulation of Hg and Se (see Figure 4) which appears to reflect the formation of HgSe. Those complexes are likely to form in association with substantial quantities of HgS, and Se will similarly form covalent associations with other electrophiles (e.g., Cd) whose compositions, binding affinities, and tissue kinetics remain uncharacterized. Of the moieties formed, HgSe is the least soluble and most stable.
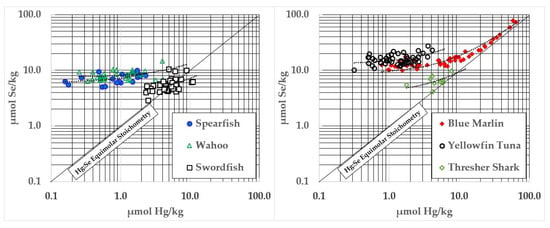
Figure 4.
Ocean fish with significant relationships between Hg and Se contents.
Coaccumulation of HgSe was particularly apparent in blue marlin, but other fish showed highly significant (p < 0.0001) accumulations of these elements (See Table 2). The mean of the intercepts observed in these fish was 7.17 ± 5.51 µmol Hg/kg—a value that coincides well with the ~10 µmol Hg/kg for the initiation of HgSe formation reported by Dietz et al. [33].
4. Discussion
Aside from mako shark and large swordfish, Se was more abundant than Hg in the ocean fish assessed in this study and would prevent rather than contribute to harm from the Hg present. While increasing fish weights were associated with increasing Hg contents [34], their HBVs generally held stable and remained positive. This is because Se homeostasis dynamically regulates tissue Se concentrations to ensure adequate amounts are available to support selenoenzyme synthesis and activities [7,32]. Only mako sharks had negative HBVs with values (−16.4) similar to those of pilot whales −14.8 [32]. In contrast, the pilot whales consumed during the Faroes study [35] had HBVs that averaged −82.3.
As noted by Dietz et al. [33] HgSe coaccumulation is observed in many species and appears at concentrations above ~10 µmol Hg/kg. Since the HgSe form resists degradation by concentrated acids other than aqua regia, this had initially been assumed to reflect a “mutual detoxification” of Hg and Se. Before the roles of Se in protecting the brain against oxidative damage were recognized, Se was often considered a toxic substance. While excessive Se intake from supplements has been associated with brittle hair and loss of hair and nails, the concentrations of Se even in Se-rich foods are unlikely to result in adverse effects. Selenosis characterized by brittleness and loss of hair and nails was observed in certain regions of China [36], but this was only after sustained dietary Se intakes of 3200–6690 µg Se/day (~40–80 µmol/day). Such chronically high exposures have not been seen elsewhere. These equate to eating 2–4 kg/day of skipjack, one of the fish with the highest Se contents observed in this study. As a precaution, 400 µg Se/day (~5 µmol/day) is the upper limit recommended for continuous dietary intake over a lifetime. This would be equivalent to eating ~250 g of skipjack or ~500 g of tuna per day. While these intakes are possible, they are unlikely to be consistently maintained.
As tissue Hg concentrations rose above ~7 µmol Hg/kg, there was an apparent coaccumulation of Hg and Se in spearfish, wahoo, swordfish, yellowfin, blue marlin, and thresher shark. This appears to indicate HgSe formation, but will need synchrotron analysis to be confirmed [37,38]. In blue marlin (Figure 4), the concentration of Se in relation to Hg in samples above 10 µmol Hg/kg was y = 1.145x + 1.904, Adjusted R2 = 0.97, p < 0.0001. These samples had an average Hg:Se molar ratio of 0.74 ± 0.17 to 1. Coaccumulation of HgSe was evident, but the Se in those samples averaged ~2 µmol/kg higher than Hg and increased faster than Hg (1.145x). However, HgSe formation in blue marlin does not appear to be universal. For example, mako shark significantly exceeded the 1:1 Hg:Se molar ratio (See Table 2) but Se contents did not increase in association with Hg (see Table 3). This may be due to the small number of samples and the limited weight range that was assessed but could also indicate metabolic or dietary differences between shark and other types of fish. Large swordfish, and a few samples of monchong, opah, and escolar also exceeded the 1:1 Hg:Se molar ratio, although not by large amounts (Figure 3). Thresher shark Hg did not exceed 1:1 stoichiometries with Se (Table 2 and Figure 4), so it would not be appropriate to recommend pregnant women limit consumption of all shark species. However, further work will need to be performed to determine if larger thresher sharks might develop negative HBVs above a certain size.
The current understanding of the mechanism of Hg toxicity is that Hg-dependent Se sequestration can cause a conditioned Se deficiency that diminishes the activities of the selenoenzymes required for neurodevelopment and to prevent and reverse oxidative damage in brain and other tissues. The formation of HgSe in fish muscle meats reflects this mechanism but it should be noted that formation of HgSe is far more extensive in the brain and other organs which have higher rates of selenoenzyme turnover [21,25]. Although tissue Se levels in the brains of Hg-poisoned individuals would gradually recover, the HgSe that forms during toxic Hg exposures remains long after the damage has been done to Se-deprived tissues [38].
Based on the mechanisms of Hg toxicity [6,7], instead of posing health risks, ocean fish that contain Se in molar excess of Hg will prevent rather than contribute to causing the conditional Se-deficiency responsible for the pathological effects of Hg toxicity [30,32]. Aside from certain species of shark and a few other apex predators, ocean fish generally provide rich amounts of Se and their consumption ensures selenoenzymes are able to continue to protect the brain from oxidative damage and perform the other physiological functions required for neurodevelopment and brain health [20]. Therefore, instead of causing harm due to Hg, eating ocean fish prevents its negative effects.
The findings of epidemiological studies show substantial benefits [15] accompany ocean fish consumption with no evidence of harm from the co-exposures of CH3Hg+. These results coincide with expectations that maternal Se intake from ocean fish consumption would prevent instead of induce a Se deficiency in the brains of their developing children [20,39]. The findings of the epidemiological studies coincide with expectations based on the positive HBVs of the ocean fish that were consumed.
The positive HBVs of bigeye tuna and marlin species indicate they clearly do not belong in the choices to avoid category in the guidance provided by the U.S. FDA [4]. Although mako shark HBVs were negative, the HBVs of the thresher sharks assessed in this study were consistently positive, so warnings should not be directed against all shark species and should be updated to reflect notable differences between species. Swordfish HBVs varied in relation to body size, but the mean value was positive. The relationships between Hg and Se in king mackerel, orange roughy, and tilefish from the Gulf of Mexico will need to be established to assess their HBVs. Progress in assessing Hg and Se in ocean fish is ongoing around the world [14,18,40,41,42,43,44,45,46,47,48,49,50,51,52,53]. Future work will need to examine time-dependent changes and identify any location-dependent differences in Hg, Se, and HBVs among these fish species collected from different regions of the world.
Freshwater fish assessments are occurring, but greater attention in establishing their HBVs will be required, particularly in fish from Se-poor watersheds [18,41,54,55,56,57]. While there is reassurance to be found in that fish from most freshwater bodies areas have substantially higher Se than Hg, this is not uniformly true [26,55] and risks of Hg and Cd exposures would be magnified among those with low-Se status. Therefore, risks to subsistence freshwater fish consumers from exposures to these and other soft electrophiles could pose greater risks than those reported in association with eating marine mammals in the Faroes. Subsistence consumers of marine mammals may have accentuated exposures to metallic and organic electrophiles and other persistent bioaccumulative toxicants (e.g., PCBs) and should be assessed to limit risks and improve health.
Ocean fish are rich sources of Se, iron, iodine, zinc, vitamins A, D, B12, and omega-3 fatty acids required for fetal brain development and also provide calcium, vitamins E and B6, potassium, choline, and lean protein which are essential for health throughout the lifespan [58]. While reduction of contaminant exposures is being accomplished through cautions to limit exposure from food sources containing potentially hazardous agents, there is a need for public health policies and advisories to devote more attention to the nutritional risk/benefit relationship in order to provide more balanced recommendations. The FAO/WHO has taken important steps in this direction with their recent report although misunderstandings of the Hg-Se relationship remains and there is a need to more clearly differentiate exposure sources considered in past evaluations and interpretations of data [18]. Further evaluations of ocean and freshwater fish Hg–Se relationships and an expansion of the database to include other foods which can be sources of soft electrophiles such as rice, marine mammals, and shellfish are necessary. Increasing the adoption of the HBV criterion will improve recognition of neurodevelopmental and other health benefits associated with ocean fish consumption.
5. Conclusions
Fourteen of the 15 commonly consumed ocean fish assessed in this study had positive HBVs indicating these fish are beneficial and safe to consume during pregnancy. Ocean fish with positive HBVs offer nutritional benefits while negative HBV seafoods contain more Hg than Se and may therefore decrease these benefits [21]. When Hg concentrations are high in fish tissues, HgSe appears to form. Since Se binding and the irreversible inhibition of selenoenzymes is recognized as the direct biochemical mechanism responsible for the consequences of Hg toxicity, this finding is consistent with expectations. Selenium from ocean fish consumption protects against Hg-dependent Se deficiencies and supports the synthesis and activities of selenoenzymes required for adult health and fetal brain development. Increased maternal intake of ocean fish is accompanied by improved child IQ, social and scholastic abilities, and provides maternal health benefits, therefore public health messages need to more clearly convey that avoiding or limiting seafood consumption during pregnancy will result in the loss of these benefits.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fishes10040158/s1.
Author Contributions
Conceptualization, methodology, formal analysis, original draft preparation, review, and editing; N.V.C.R., J.J.K. and L.J.R. All authors have read and agreed to the published version of the manuscript.
Funding
The original sampling and analysis were supported by NOAA Award No. NA05NMF4521112 to PacMar, Honolulu, Hawaii. Development and documentation of HBV data used in this study was performed with funding from the U.S. E.P.A. National Center for Environmental Research (NCER) Science to Achieve Results (STAR) grant RD834792-01: Fish Selenium Health Benefit Values in Mercury Risk Management. The reanalysis performed for this article received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived since only food portions obtained from the individual fish were assessed for this study.
Data Availability Statement
The original data presented in the study have been published in the Supplementary Materials of this paper.
Conflicts of Interest
The original funding agencies had no role in the preparation or decision to submit this article for publication. This article has not been reviewed by the funding agencies and no official endorsements should be inferred.
Abbreviations
The following abbreviations are used in this manuscript:
| Cd | Cadmium |
| CH3Hg+ | Methylmercury |
| Cys | Cysteine |
| FAO | Food and Agriculture Organization |
| HBV | Health benefit value |
| Hg | Mercury |
| HgSe | Mercury selenide |
| IQ | Intelligence quotient |
| PCB | Polychlorinated biphenyl |
| RfD | Reference dose |
| Se | Selenium |
| Sec | Selenocysteine |
| U.S. EPA | United States Environmental Protection Agency |
| U.S. FDA | United States Food and Drug Administration |
| WHO | World Health Organization |
References
- Calder, P.C.; Yaqoob, P. Omega-3 Polyunsaturated Fatty Acids and Human Health Outcomes. BioFactors 2009, 35, 266–272. [Google Scholar] [CrossRef]
- Hibbeln, J.R.; Davis, J.M.; Steer, C.; Emmett, P.; Rogers, I.; Williams, C.; Golding, J. Maternal Seafood Consumption in Pregnancy and Neurodevelopmental Outcomes in Childhood (ALSPAC Study): An Observational Cohort Study. Lancet 2007, 369, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Byrd, K.A.; Thilsted, S.H.; Fiorella, K.J. Fish Nutrient Composition: A Review of Global Data from Poorly Assessed Inland and Marine Species. Public Health Nutr. 2021, 24, 476–486. [Google Scholar] [CrossRef]
- FDA Advice About Eating Fish. Available online: https://www.fda.gov/food/consumers/advice-about-eating-fish (accessed on 4 January 2025).
- Clarkson, T.W.; Magos, L. The Toxicology of Mercury and Its Chemical Compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef] [PubMed]
- Ralston, N.V.C. Mercury’s Neurotoxic Effects on Brain Selenoenzymes. In Handbook of Neurotoxicity; Kostrzewa, R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 1–27. ISBN 978-3-030-71519-9. [Google Scholar]
- Ralston, N.V.C.; Raymond, L.J. Mercury’s Neurotoxicity Is Characterized by Its Disruption of Selenium Biochemistry. Biochim. Biophys. Acta (BBA) Gen. Subj. 2018, 1862, 2405–2416. [Google Scholar] [CrossRef]
- Nishigaki, S.; Harada, M. Methylmercury and Selenium in Umbilical Cords of Inhabitants of the Minamata Area. Nature 1975, 258, 324–325. [Google Scholar] [CrossRef]
- Sakamoto, M.; Marumoto, M.; Haraguchi, K.; Toyama, T.; Saito, Y.; Balogh, S.J.; Tohyama, C.; Nakamura, M. Assessing the Role of Selenium in Minamata Disease through Reanalysis of Historical Samples. Environ. Int. 2025, 195, 109242. [Google Scholar] [CrossRef]
- Bakir, F.; Damluji, S.F.; Amin-Zaki, L.; Murtadha, M.; Khalidi, A.; Al-Rawi, N.Y.; Tikriti, S.; Dhahir, H.I.; Clarkson, T.W.; Smith, J.C.; et al. Methylmercury Poisoning in Iraq: An Interuniversity Report. Science 1973, 181, 230–241. [Google Scholar] [CrossRef]
- Harris, H.H.; Pickering, I.J.; George, G.N. The Chemical Form of Mercury in Fish. Science 2003, 301, 1203. [Google Scholar] [CrossRef]
- Davidson, P.W.; Cory-Slechta, D.A.; Thurston, S.W.; Huang, L.-S.; Shamlaye, C.F.; Gunzler, D.; Watson, G.; van Wijngaarden, E.; Zareba, G.; Klein, J.D.; et al. Fish Consumption and Prenatal Methylmercury Exposure: Cognitive and Behavioral Outcomes in the Main Cohort at 17 Years from the Seychelles Child Development Study. Neurotoxicology 2011, 32, 711–717. [Google Scholar] [CrossRef]
- Klus, J.K.; Thurston, S.W.; Myers, G.J.; Watson, G.E.; Rand, M.D.; Love, T.M.; Yeates, A.J.; Mulhern, M.S.; McSorley, E.M.; Strain, J.J.; et al. Postnatal Methylmercury Exposure and Neurodevelopmental Outcomes at 7 Years of Age in the Seychelles Child Development Study Nutrition Cohort 2. NeuroToxicology 2023, 99, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, M.; Yeates, A.J.; McSorley, E.M.; Watson, G.E.; Van Wijngaarden, E.; Bodin, N.; Govinden, R.; Jean-Baptiste, J.; Desnousse, S.; Shamlaye, C.F.; et al. Dietary Selenium and Mercury Intakes from Fish Consumption during Pregnancy: Seychelles Child Development Study Nutrition Cohort 2. NeuroToxicology 2024, 101, 1–5. [Google Scholar] [CrossRef]
- Spiller, P.; Van Wijngaarden, E.; Adams, H.R.; Strain, J.J.; McSorley, E.M.; Mulhern, M.S.; Conway, M.C.; Yeates, A.J.; Carrington, C.; Bolger, P.M.; et al. Net Effects Explains the Benefits to Children from Maternal Fish Consumption despite Methylmercury in Fish. NeuroToxicology 2023, 99, 195–205. [Google Scholar] [CrossRef]
- Raymond, L.; Ralston, N. Seafood, Selenium, and Pregnancy. In Handbook of Public Health Nutrition: International, National, and Regional Perspectives; Springer International Publishing: Berlin/Heidelberg, Germany, 2025. [Google Scholar]
- Oken, E. Decline in Fish Consumption among Pregnant Women after a National Mercury Advisory. Obstet. Gynecol. 2003, 102, 346–351. [Google Scholar] [CrossRef]
- FAO; WHO. Joint FAO/WHO Expert Consultation on the Risks and Benefits of Fish Consumption; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2024; ISBN 978-92-5-139107-5. [Google Scholar]
- Oken, E.; Musci, R.J.; Westlake, M.; Gachigi, K.; Aschner, J.L.; Barnes, K.L.; Bastain, T.M.; Buss, C.; Camargo, C.A.; Cordero, J.F.; et al. Demographic and Health Characteristics Associated with Fish and n-3 Fatty Acid Supplement Intake during Pregnancy: Results from Pregnancy Cohorts in the ECHO Programme. Public Health Nutr. 2024, 27, e94. [Google Scholar] [CrossRef] [PubMed]
- Ralston, N.V.C.; Raymond, L.J.; Gilman, C.L.; Soon, R.; Seale, L.A.; Berry, M.J. Maternal Seafood Consumption Is Associated with Improved Selenium Status: Implications for Child Health. NeuroToxicology 2024, 101, 26–35. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Raymond, L.J. Dietary Selenium’s Protective Effects against Methylmercury Toxicity. Toxicology 2010, 278, 112–123. [Google Scholar] [CrossRef] [PubMed]
- George, G.N.; MacDonald, T.C.; Korbas, M.; Singh, S.P.; Myers, G.J.; Watson, G.E.; O’Donoghue, J.L.; Pickering, I.J. The Chemical Forms of Mercury and Selenium in Whale Skeletal Muscle. Metallomics 2011, 3, 1232. [Google Scholar] [CrossRef]
- Dyrssen, D.; Wedborg, M. The Sulphur-Mercury(II) System in Natural Waters. Water Air Soil Pollut. 1991, 56, 507–519. [Google Scholar] [CrossRef]
- Nuttall, K.L. A Model for Metal Selenide Formation under Biological Conditions. Med. Hypotheses 1987, 24, 217–221. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Blackwell, J.L.; Raymond, L.J. Importance of Molar Ratios in Selenium-Dependent Protection against Methylmercury Toxicity. Biol. Trace Elem. Res. 2007, 119, 255–268. [Google Scholar] [CrossRef]
- US EPA; Ralston, N.; Raymond, L. Final Report|Fish Selenium Health Benefit Values in Mercury Risk Management|Research Project Database|Grantee Research Project|ORD|US EPA. Available online: https://cfpub.epa.gov/ncer_abstracts//index.cfm (accessed on 18 January 2025).
- Ralston, N.V.C. Concomitant Selenoenzyme Inhibitor Exposures as Etiologic Contributors to Disease: Implications for Preventative Medicine. Arch. Biochem. Biophys. 2023, 733, 109469. [Google Scholar] [CrossRef]
- Choi, A.L.; Budtz-Jørgensen, E.; Jørgensen, P.J.; Steuerwald, U.; Debes, F.; Weihe, P.; Grandjean, P. Selenium as a Potential Protective Factor against Mercury Developmental Neurotoxicity. Environ. Res. 2008, 107, 45–52. [Google Scholar] [CrossRef]
- Budtz-Jørgensen, E.; Grandjean, P.; Weihe, P. Separation of Risks and Benefits of Seafood Intake. Env. Health Perspect. 2007, 115, 323–327. [Google Scholar] [CrossRef]
- Kaneko, J.J.; Ralston, N.V.C. Selenium and Mercury in Pelagic Fish in the Central North Pacific near Hawaii. Biol. Trace Elem. Res. 2007, 119, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Ralston, N.V.C.; Ralston, C.R.; Raymond, L.J. Selenium Health Benefit Values: Updated Criteria for Mercury Risk Assessments. Biol. Trace Elem. Res. 2016, 171, 262–269. [Google Scholar] [CrossRef]
- Ralston, N.V.C.; Kaneko, J.J.; Raymond, L.J. Selenium Health Benefit Values Provide a Reliable Index of Seafood Benefits vs. Risks. J. Trace Elem. Med. Biol. 2019, 55, 50–57. [Google Scholar] [CrossRef]
- Dietz, R.; Riget, F.; Born, E. An Assessment of Selenium to Mercury in Greenland Marine Animals. Sci. Total Environ. 2000, 245, 15–24. [Google Scholar] [CrossRef]
- IOM. Toxicological Effects of Methylmercury; National Academies Press: Washington, DC, USA, 2000; ISBN 0-309-56970-2. [Google Scholar]
- Grandjean, P.; Weihe, P.; Jørgensen, P.J.; Clarkson, T.; Cernichiari, E.; Viderø, T. Impact of Maternal Seafood Diet on Fetal Exposure to Mercury, Selenium, and Lead. Arch. Environ. Health Int. J. 1992, 47, 185–195. [Google Scholar] [CrossRef]
- Yang, G.; Wang, S.; Zhou, R.; Sun, S. Endemic Selenium Intoxication of Humans in China. Am. J. Clin. Nutr. 1983, 37, 872–881. [Google Scholar] [CrossRef]
- Huggins, F.E.; Raverty, S.A.; Nielsen, O.S.; Sharp, N.E.; Robertson, J.D.; Ralston, N.V.C. An XAFS Investigation of Mercury and Selenium in Beluga Whale Tissues. Environ. Bioindic. 2009, 4, 291–302. [Google Scholar] [CrossRef]
- Korbas, M.; O’Donoghue, J.L.; Watson, G.E.; Pickering, I.J.; Singh, S.P.; Myers, G.J.; Clarkson, T.W.; George, G.N. The Chemical Nature of Mercury in Human Brain Following Poisoning or Environmental Exposure. ACS Chem. Neurosci. 2010, 1, 810–818. [Google Scholar] [CrossRef]
- Ralston, N.; Azenkeng, A.; Ralston, C.; Blackwell, J.L.; Raymond, L. Seafood Science: Advances in Chemistry, Technology and Applications; CRC Press: Boca Raton, FL, USA, 2015; pp. 433–457. [Google Scholar]
- Polak-Juszczak, L. Selenium and Mercury Molar Ratios in Commercial Fish from the Baltic Sea: Additional Risk Assessment Criterion for Mercury Exposure. Food Control 2015, 50, 881–888. [Google Scholar] [CrossRef]
- Bridges, K.N.; Furin, C.G.; Gerlach, R.F. Subsistence Fish Consumption in Rural Alaska: Using Regional Monitoring Data to Evaluate Risk and Bioavailability of Dietary Methylmercury. Sci. Total Environ. 2020, 736, 139676. [Google Scholar] [CrossRef]
- Zhu, Y.; Ho, Q.T.; Dahl, L.; Azad, A.M.; Bank, M.S.; Boitsov, S.; Kjellevold, M.; Kögel, T.; Lien, V.S.; Lundebye, A.-K.; et al. Predicting Essential and Hazardous Element Concentrations in Marine Fish from the Northeast Atlantic Ocean: A Bayesian Approach. Sci. Total Environ. 2025, 968, 178748. [Google Scholar] [CrossRef]
- Ruelas-Inzunza, J.; Šlejkovec, Z.; Mazej, D.; Fajon, V.; Horvat, M.; Ramos-Osuna, M. Bioaccumulation of As, Hg, and Se in Tunas Thunnus Albacares and Katsuwonus Pelamis from the Eastern Pacific: Tissue Distribution and As Speciation. Env. Sci. Pollut. Res. 2018, 25, 19499–19509. [Google Scholar] [CrossRef]
- Ordiano-Flores, A.; Rosíles-Martínez, R.; Galván-Magaña, F. Biomagnification of Mercury and Its Antagonistic Interaction with Selenium in Yellowfin Tuna Thunnus Albacares in the Trophic Web of Baja California Sur, Mexico. Ecotoxicol. Environ. Saf. 2012, 86, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.J.; Butler, E.C.V.; Macleod, C.K. Spatial Variability in Selenium and Mercury Interactions in a Key Recreational Fish Species: Implications for Human Health and Environmental Monitoring. Mar. Pollut. Bull. 2013, 74, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Annibaldi, A.; Truzzi, C.; Carnevali, O.; Pignalosa, P.; Api, M.; Scarponi, G.; Illuminati, S. Determination of Hg in Farmed and Wild Atlantic Bluefin Tuna (Thunnus thynnus L.) Muscle. Molecules 2019, 24, 1273. [Google Scholar] [CrossRef]
- Lazarini, T.E.D.M.; Milani, R.F.; Morgano, M.A. Selenium, Total Mercury and Methylmercury in Sardine: Study of Molar Ratio and Protective Effect on the Diet. J. Environ. Sci. Health Part B 2019, 54, 387–393. [Google Scholar] [CrossRef]
- Melgar, M.J.; Núñez, R.; García, M.Á. Selenium Intake from Tuna in Galicia (Spain): Health Risk Assessment and Protective Role against Exposure to Mercury and Inorganic Arsenic. Sci. Total Environ. 2019, 694, 133716. [Google Scholar] [CrossRef] [PubMed]
- Vega-Sánchez, B.; Ortega-García, S.; Ruelas-Inzunza, J.; Frías-Espericueta, M.; Escobar-Sánchez, O.; Jara-Marini, M. Selenium and Mercury in Dolphinfish (Coryphaena hippurus) from the Gulf of California: Inter-Annual Variations and Selenium Health Benefit Value. Env. Sci. Pollut. Res. 2020, 27, 2311–2318. [Google Scholar] [CrossRef]
- Medina-Morales, S.A.; Corro-Espinosa, D.; Escobar-Sánchez, O.; Delgado-Alvarez, C.G.; Ruelas-Inzunza, J.; Frías-Espericueta, M.G.; Jara-Marini, M.E.; Páez-Osuna, F. Mercury (Hg) and Selenium (Se) Content in the Shark Mustelus Henlei (Triakidae) in the Northern Mexican Pacific. Env. Sci. Pollut. Res. 2020, 27, 16774–16783. [Google Scholar] [CrossRef]
- Teixeira, G.; Raimundo, J.; Goulart, J.; Costa, V.; Menezes, G.M.; Caetano, M.; Pacheco, M.; Martins, I. Hg and Se Composition in Demersal Deep-Sea Fish from the North-East Atlantic. Env. Sci. Pollut. Res. 2020, 27, 33649–33657. [Google Scholar] [CrossRef]
- Barone, G.; Storelli, A.; Meleleo, D.; Dambrosio, A.; Garofalo, R.; Busco, A.; Storelli, M.M. Levels of Mercury, Methylmercury and Selenium in Fish: Insights into Children Food Safety. Toxics 2021, 9, 39. [Google Scholar] [CrossRef]
- Rudershausen, P.J.; Cross, F.A.; Runde, B.J.; Evans, D.W.; Cope, W.G.; Buckel, J.A. Total Mercury, Methylmercury, and Selenium Concentrations in Blue Marlin Makaira Nigricans from a Long-Term Dataset in the Western North Atlantic. Sci. Total Environ. 2023, 858, 159947. [Google Scholar] [CrossRef]
- Cusack, L.K.; Eagles-Smith, C.; Harding, A.K.; Kile, M.; Stone, D. Selenium: Mercury Molar Ratios in Freshwater Fish in the Columbia River Basin: Potential Applications for Specific Fish Consumption Advisories. Biol. Trace Elem. Res. 2017, 178, 136–146. [Google Scholar] [CrossRef]
- Peterson, S.A.; Ralston, N.V.C.; Whanger, P.D.; Oldfield, J.E.; Mosher, W.D. Selenium and Mercury Interactions with Emphasis on Fish Tissue. Environ. Bioindic. 2009, 4, 318–334. [Google Scholar] [CrossRef]
- Johnson, T.K.B.; LePrevost, C.E.; Kwak, T.J.; Cope, W.G. Selenium, Mercury, and Their Molar Ratio in Sportfish from Drinking Water Reservoirs. Int. J. Environ. Res. Public Health 2018, 15, 1864. [Google Scholar] [CrossRef]
- Plessl, C.; Gilbert, B.M.; Sigmund, M.F.; Theiner, S.; Avenant-Oldewage, A.; Keppler, B.K.; Jirsa, F. Mercury, Silver, Selenium and Other Trace Elements in Three Cyprinid Fish Species from the Vaal Dam, South Africa, Including Implications for Fish Consumers. Sci. Total Environ. 2019, 659, 1158–1167. [Google Scholar] [CrossRef]
- Liu, C.; Ralston, N.V.C. Seafood and Health: What You Need to Know? In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2021; Volume 97, pp. 275–318. ISBN 978-0-12-824580-4. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).