Maneuvering Performance in the Colonial Siphonophore, Nanomia bijuga


Abstract
1. Introduction
2. Materials and Methods
2.1. Fluid Mechanics
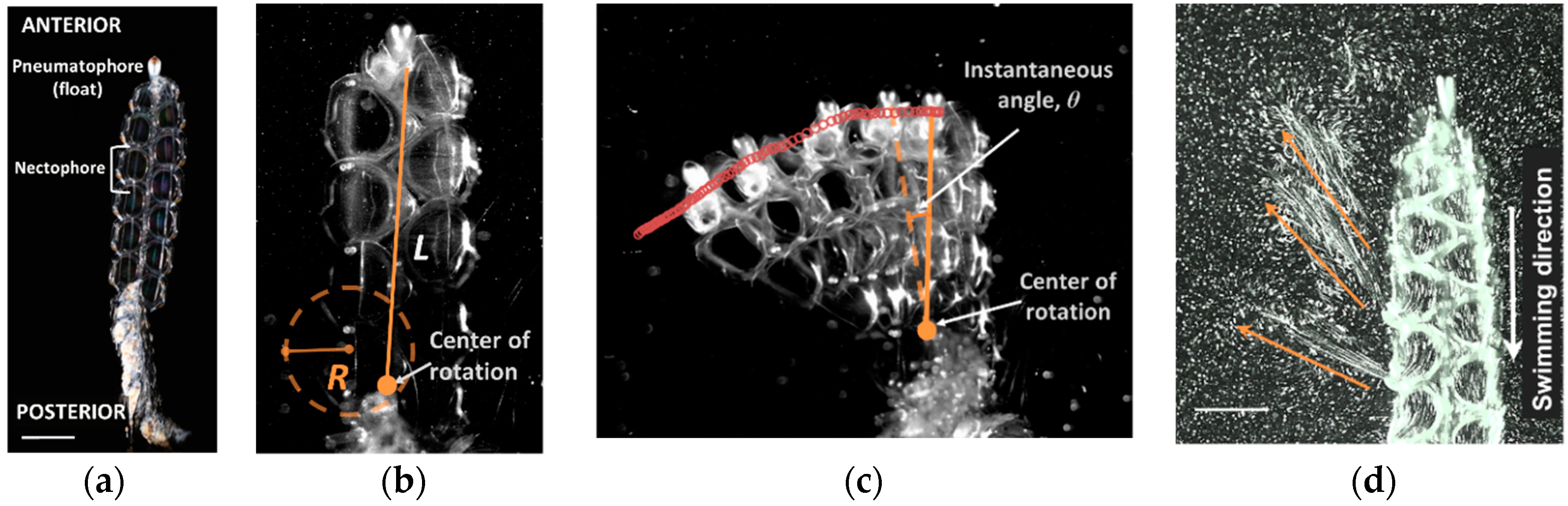
2.2. Kinematics
2.3. Turning Performance
3. Results
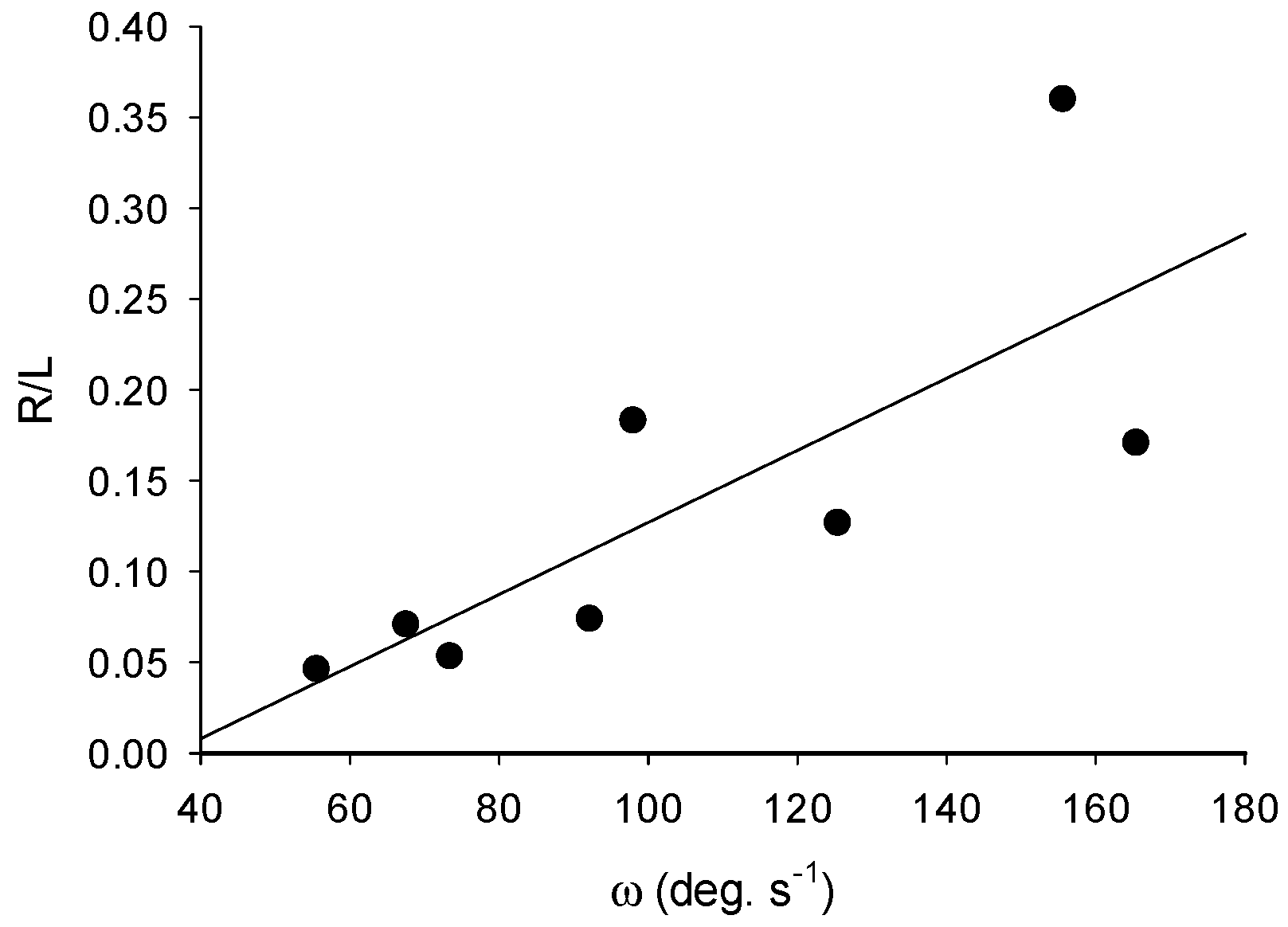
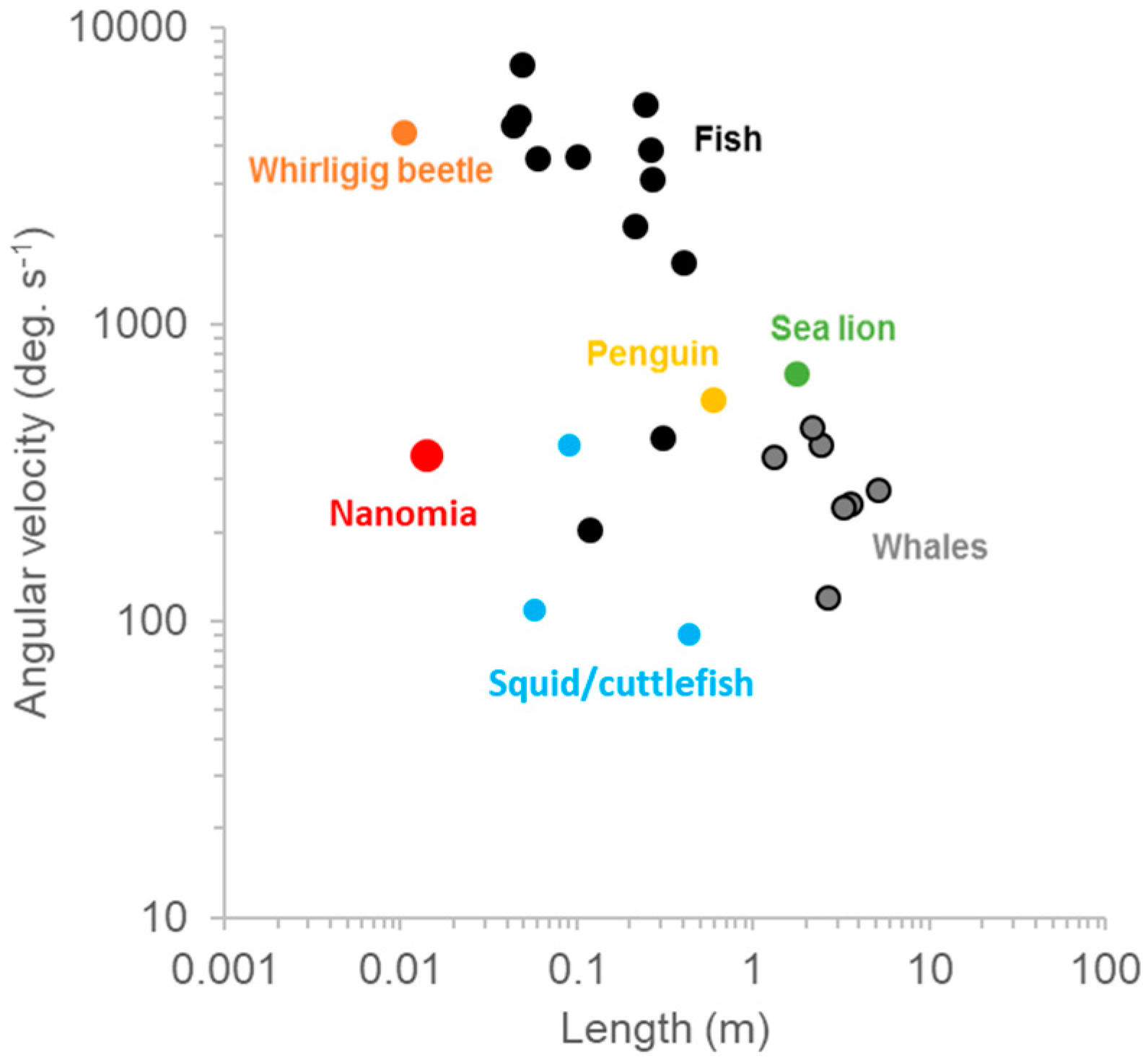
3.1. Turning
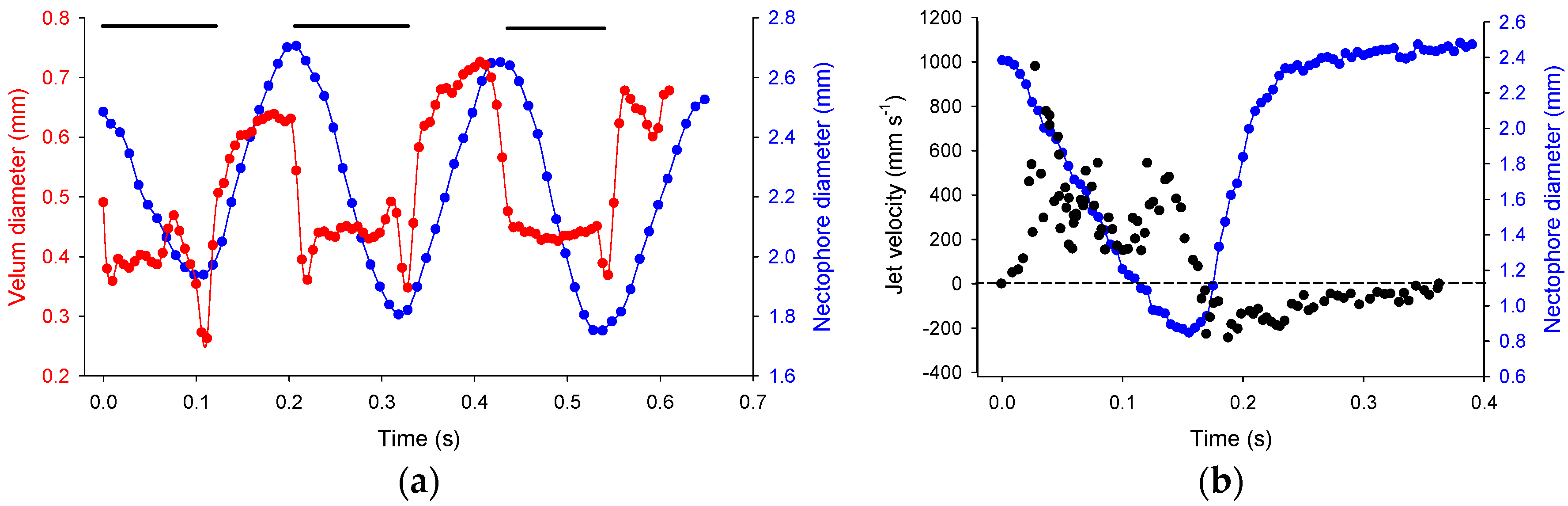
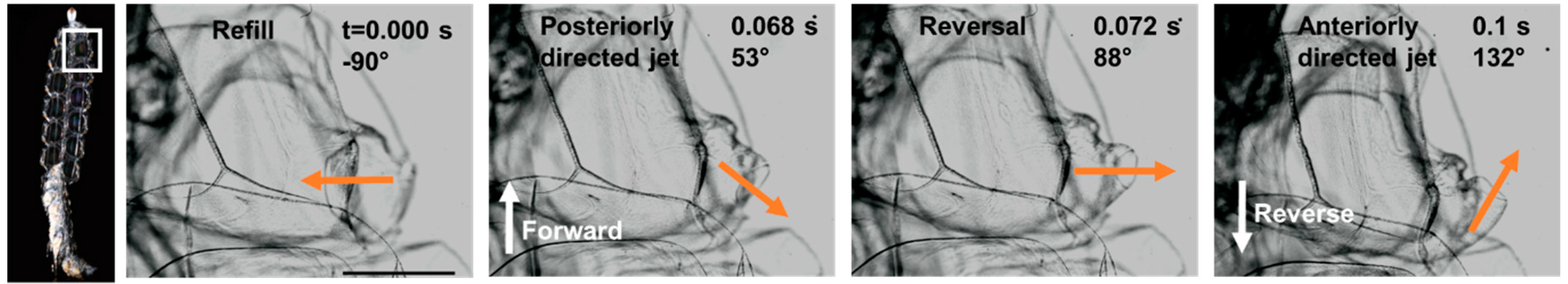
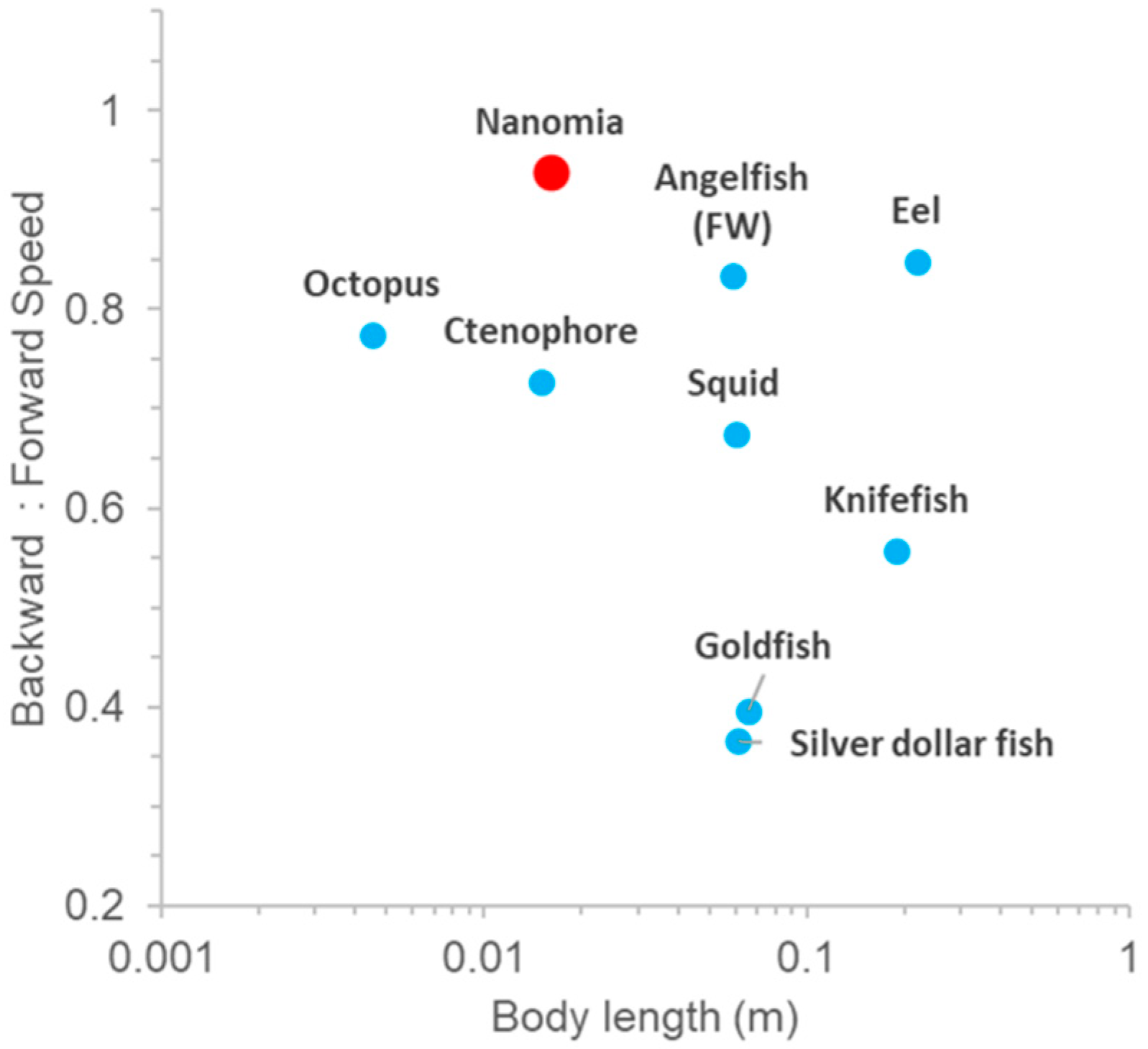
3.2. Reverse Swimming
4. Discussion
4.1. Turning
4.2. Reversal
4.3. Neurological Control
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruiz, L.A.; Whittlesey, R.W.; Dabiri, J.O. Vortex-enhanced propulsion. J. Fluid Mech. 2011, 668, 5–32. [Google Scholar] [CrossRef]
- Moslemi, A.A.; Krueger, P.S. Propulsive efficiency of a biomorphic pulsed-jet underwater vehicle. Bioinspir. Biomim. 2010, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Serchi, F.G.; Arienti, A.; Laschi, C. Biomimetic vortex propulsion: Toward the new paradigm of soft unmanned underwater vehicles. IEEE ASME Trans. Mechatron. 2012, 18, 484–493. [Google Scholar] [CrossRef]
- Mohensi, K. Zero-mass pulsatile jets for unmanned underwater vehicle maneuvering. In Proceedings of the AIAA 3rd “Unmanned Unlimited” Technical Conference, Workshop and Exhibit, Chicago, IL, USA, 20–23 September 2004; Volume 6386. [Google Scholar]
- Barham, E.G. Siphonophores and the Deep Scattering Layer. Science 1963, 140, 826–828. [Google Scholar] [CrossRef]
- Mackie, G.O. Analysis of locomotion in a siphonophore colony. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1964, 159, 366–391. [Google Scholar]
- Costello, J.H.; Colin, S.P.; Gemmell, B.J.; Dabiri, J.O.; Sutherland, K.R. Multi-jet propulsion organized by clonal development in a colonial siphonophore. Nat. Commun. 2015, 6, 8158. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, K.R.; Gemmell, B.J.; Colin, S.P.; Costello, J.H. Propulsive design principles in a multi-jet siphonophore. J. Exp. Biol. 2019, 222, jeb198242. [Google Scholar] [CrossRef]
- Sutherland, K.R.; Costello, J.H.; Colin, S.P.; Dabiri, J.O. Ambient fluid motions influence swimming and feeding by the ctenophore Mnemiopsis leidyi. J. Plankton Res. 2014, 36, 1310–1322. [Google Scholar] [CrossRef]
- Gemmell, B.J.; Jiang, H.; Buskey, E.J. A new approach to micro-scale particle image velocimetry (µPIV) for quantifying flows around free-swimming zooplankton. J. Plankton Res. 2014, 36, 1396–1401. [Google Scholar] [CrossRef]
- Robison, B.H.; Reisenbichler, K.R.; Sherlock, R.E.; Silguero, J.M.B.; Chavez, F.P. Seasonal abundance of the siphonophore, Nanomia bijuga, in Monterey Bay. Deep Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 1741–1751. [Google Scholar] [CrossRef]
- Satterlie, R.A. Neuronal control of swimming in jellyfish: A comparative story. Can. J. Zool. 2002, 80, 1654–1669. [Google Scholar] [CrossRef]
- Gemmell, B.J.; Costello, J.H.; Colin, S.P.; Stewart, C.J.; Dabiri, J.O.; Tafti, D.; Priya, S. Passive energy recapture in jellyfish contributes to propulsive advantage over other metazoans. Proc. Natl. Acad. Sci. USA 2013, 110, 17904–17909. [Google Scholar] [CrossRef]
- Costello, J.H.; Colin, S.P.; Dabiri, J.O. Medusan morphospace: Phylogenetic constraints, biomechanical solutions, and ecological consequences. Invertebr. Biol. 2008, 127, 265–290. [Google Scholar] [CrossRef]
- Dabiri, J.O.; Colin, S.P.; Katija, K.; Costello, J.H. A wake-based correlate of swimming performance and foraging behavior in seven co-occurring jellyfish species. J. Exp. Biol. 2010, 213, 1217–1225. [Google Scholar] [CrossRef]
- Gemmell, B.J.; Troolin, D.R.; Costello, J.H.; Colin, S.P.; Satterlie, R.A. Control of vortex rings for manoeuvrability. J. R. Soc. Interface 2015, 12. [Google Scholar] [CrossRef]
- Jastrebsky, R.A.; Bartol, I.K.; Krueger, P.S. Turning performance in squid and cuttlefish: Unique dual-mode, muscular hydrostatic systems. J. Exp. Biol. 2016, 219, 1317–1326. [Google Scholar] [CrossRef]
- Walker, J.A. Does a rigid body limit maneuverability? J. Exp. Biol. 2000, 203, 3391–3396. [Google Scholar]
- Fish, F.E.; Lauder, G.V. Control surfaces of aquatic vertebrates: Active and passive design and function. J. Exp. Biol. 2017, 220, 4351–4363. [Google Scholar] [CrossRef]
- Fish, F.E.; Nicastro, A.J. Aquatic turning performance by the whirligig beetle: Constraints on maneuverability by a rigid biological system. J. Exp. Biol. 2003, 206, 1649–1656. [Google Scholar] [CrossRef]
- D’Août, K.; Aerts, P. A kinematic comparison of forward and backward swimming in the eel anguilla anguilla. J. Exp. Biol. 1999, 202, 1511–1521. [Google Scholar]
- Villanueva, R.; Nozais, C.; Boletzky, S.V. Swimming behaviour and food searching in planktonic Octopus vulgaris Cuvier from hatching to settlement. J. Exp. Mar. Biol. Ecol. 1997, 208, 169–184. [Google Scholar] [CrossRef]
- Bartol, I.K.; Krueger, P.S.; Jastrebsky, R.A.; Williams, S.; Thompson, J.T. Volumetric flow imaging reveals the importance of vortex ring formation in squid swimming tail-first and arms-first. J. Exp. Biol. 2016, 219, 392–403. [Google Scholar] [CrossRef]
- Webb, P.W.; Fairchild, A.G. Performance and maneuverability of three species of teleostean fishes. Can. J. Zool. 2001, 79, 1866–1877. [Google Scholar] [CrossRef]
- Youngerman, E.D.; Flammang, B.E.; Lauder, G.V. Locomotion of free-swimming ghost knifefish: Anal fin kinematics during four behaviors. Zoology 2014, 117, 337–348. [Google Scholar] [CrossRef]
- Mackie, G.O.; Mills, C.E.; Singla, C.L. Giant Axons and Escape Swimming in Euplokamis dunlapae (Ctenophora: Cydippida). Biol. Bull. 1992, 182, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Mackie, G.O.; Lander, H.J.; Rosen, B.R.; White, J. From aggregates to integrates: Physiological aspects of modularity in colonial animals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1986, 313, 175–196. [Google Scholar] [CrossRef]
- Pugh, P.R.; Baxter, E.J. A review of the physonect siphonophore genera Halistemma (Family Agalmatidae) and Stephanomia (Family Stephanomiidae). Zootaxa 2014, 3897, 1–111. [Google Scholar] [CrossRef][Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ω (deg s−1) | ωmax (deg s−1) | Acc.max (deg s−2) | R (mm) | R/L | |
|---|---|---|---|---|---|
| Mean | 104 ± 41 | 215 ± 90 | 9795 ± 7469 | 1.64 ± 0.77 | 0.15 ± 0.10 |
| Max | 165 | 363 | 21635 | 3.10 | 0.36 |
| Min | 56 | 122 | 3510 | 0.72 | 0.05 |
| N | 6 | 6 | 6 | 7 | 9 |
| # seq. | 8 | 8 | 8 | 11 | 11 |
| Jetting (s) | Refilling (s) | Total (s) | Ratio | Max. Jet Speed (mm s−1) | Max. Body Speed (mm s−1) | |
|---|---|---|---|---|---|---|
| Mean | 0.14 ± 0.02 | 0.23 ± 0.08 | 0.37 ± 0.10 | 0.67 | 1063 ± 176 | 134 ± 28 |
| Max | 0.10 | 0.10 | 0.21 | 0.43 | 879 | 98 |
| Min | 0.16 | 0.30 | 0.44 | 0.99 | 1260 | 166 |
| N | 6 | 5 | 5 | 5 | 5 | 4 |
| # seq. | 6 | 5 | 5 | 5 | 5 | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutherland, K.R.; Gemmell, B.J.; Colin, S.P.; Costello, J.H. Maneuvering Performance in the Colonial Siphonophore, Nanomia bijuga. Biomimetics 2019, 4, 62. https://doi.org/10.3390/biomimetics4030062
Sutherland KR, Gemmell BJ, Colin SP, Costello JH. Maneuvering Performance in the Colonial Siphonophore, Nanomia bijuga. Biomimetics. 2019; 4(3):62. https://doi.org/10.3390/biomimetics4030062
Chicago/Turabian StyleSutherland, Kelly R., Brad J. Gemmell, Sean P. Colin, and John H. Costello. 2019. "Maneuvering Performance in the Colonial Siphonophore, Nanomia bijuga" Biomimetics 4, no. 3: 62. https://doi.org/10.3390/biomimetics4030062
APA StyleSutherland, K. R., Gemmell, B. J., Colin, S. P., & Costello, J. H. (2019). Maneuvering Performance in the Colonial Siphonophore, Nanomia bijuga. Biomimetics, 4(3), 62. https://doi.org/10.3390/biomimetics4030062