Segmentation and Modelling of the Nuclear Envelope of HeLa Cells Imaged with Serial Block Face Scanning Electron Microscopy †

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. HeLa Cells’ Preparation and Acquisition
2.2. Ground Truth (GT)
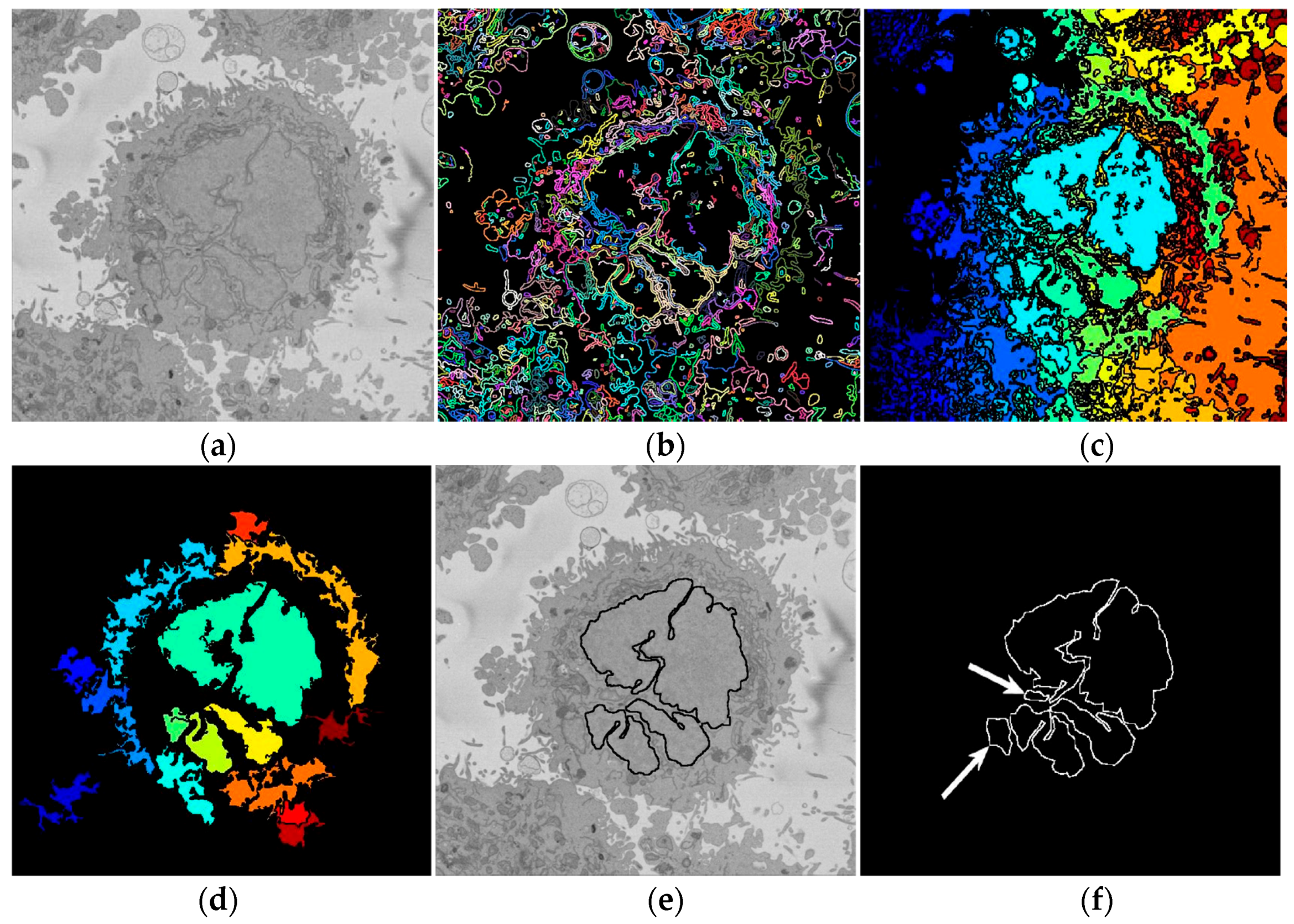
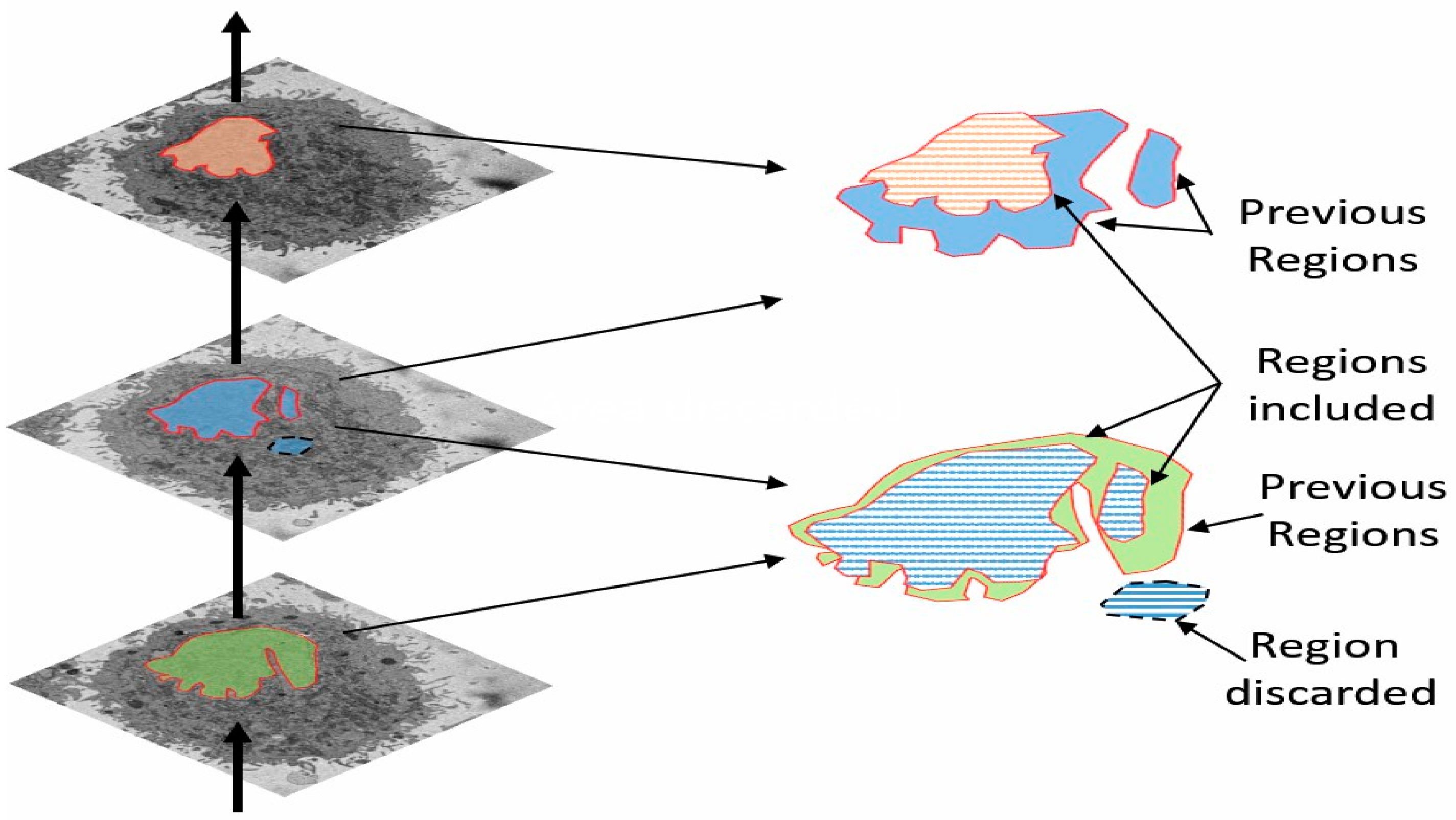
2.3. Automatic Segmentation of the Nuclear Envelope Algorithm
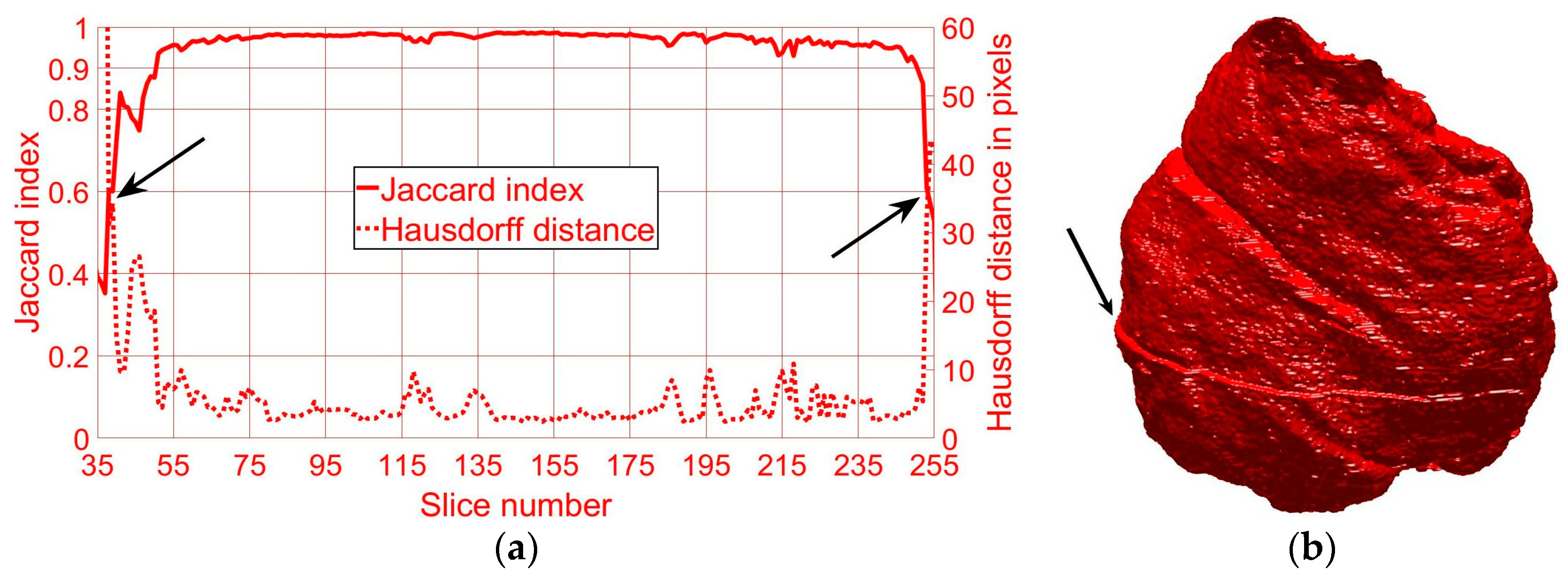
2.4. Quantitative Comparisons
2.5. Active Contours
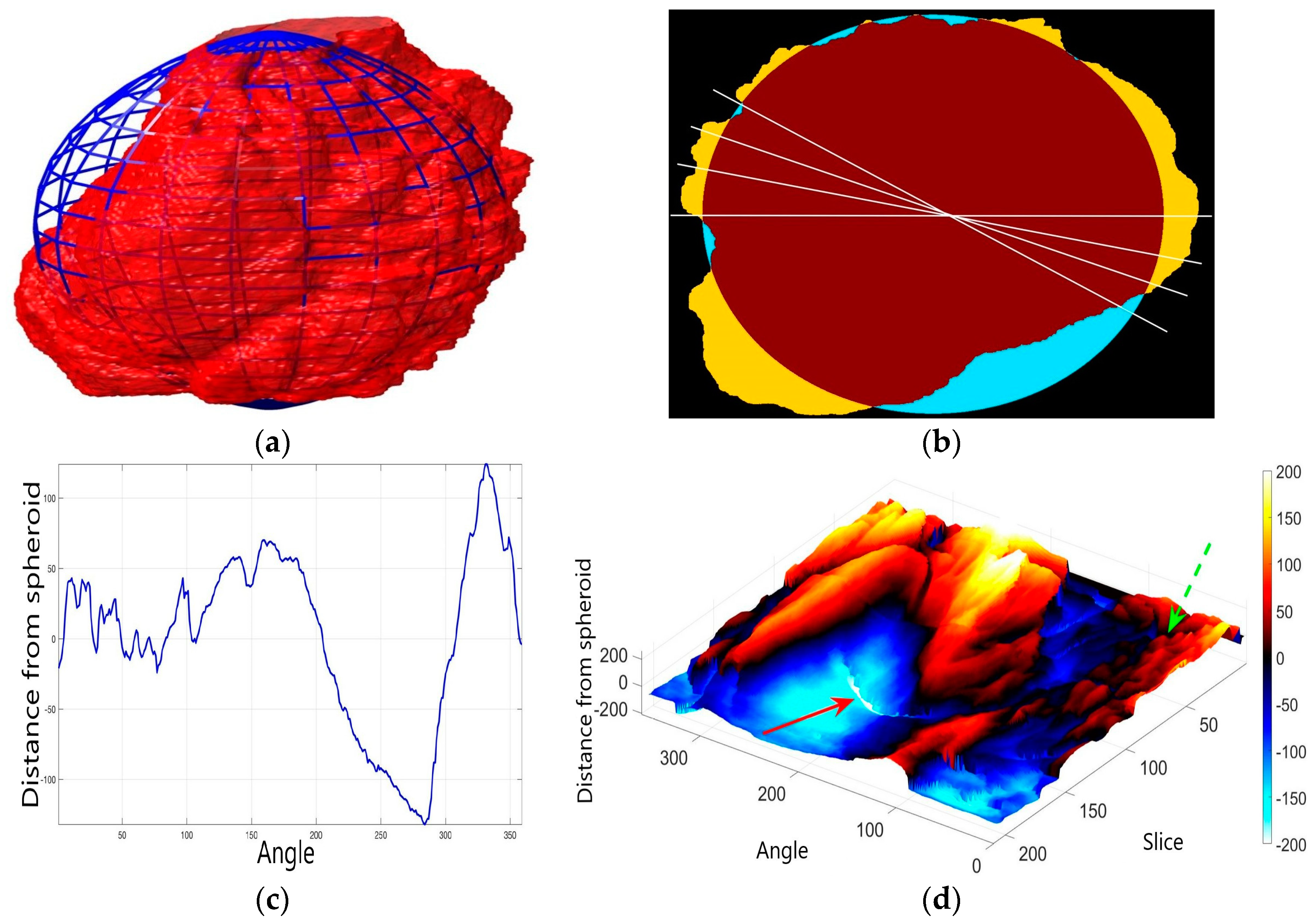
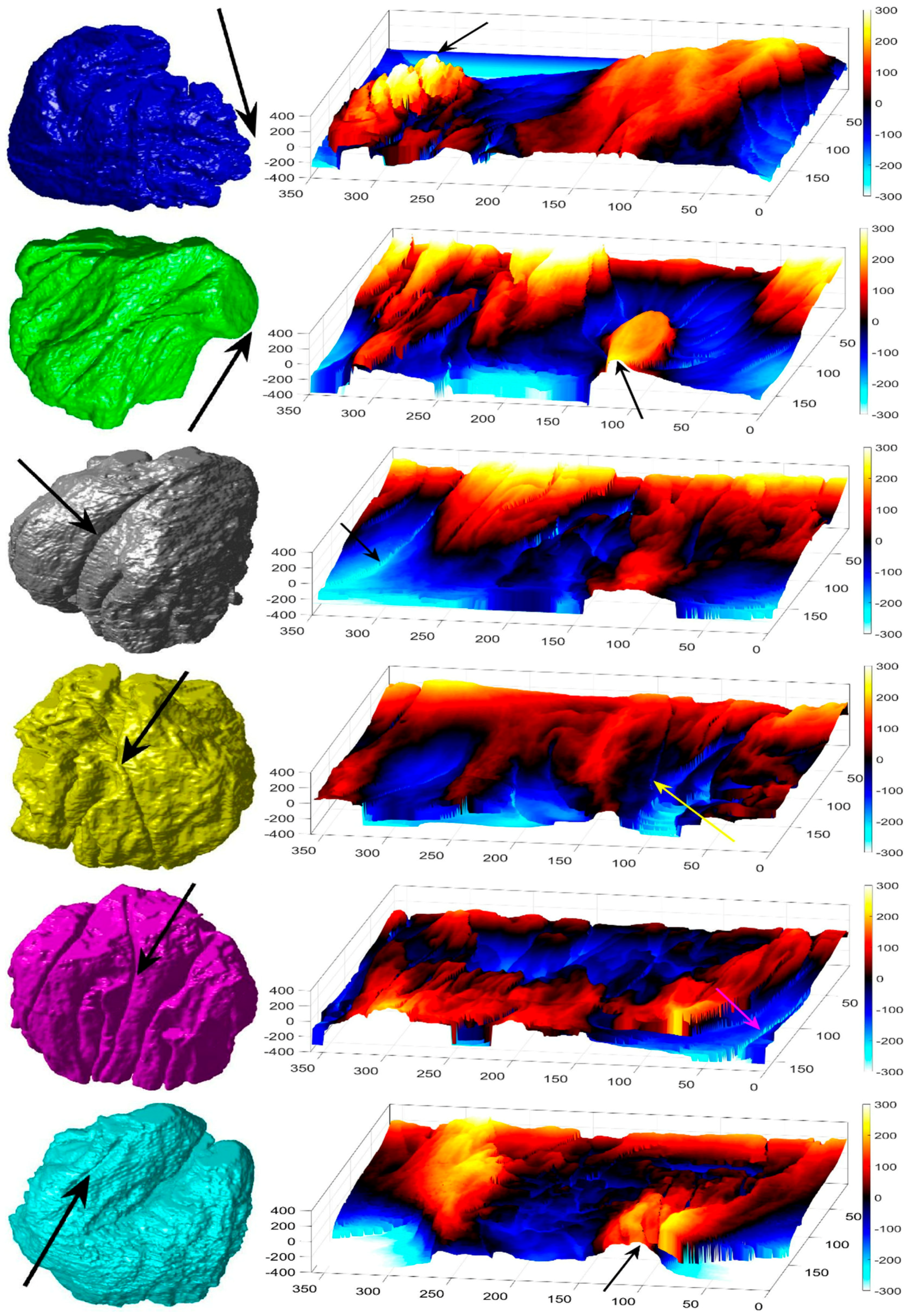
2.6. Nuclear Envelope Shape Modelling
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EM | Electron Microscopy |
| GT | Ground Truth |
| HD | Hausdorff Distance |
| JI | Jaccard Index |
| NCMIR | National Centre for Microscopy and Imaging Research |
| NE | Nuclear Envelope |
| ROI | Region of Interest |
| SBF SEM | Scanning Block Face Electron Microscopy |
| TIFF | Tagged Image File Format |
References
- Chankong, T.; Theera-Umpon, N.; Auephanwiriyakul, S. Automatic cervical cell segmentation and classification in Pap smears. Comput. Methods Programs Biomed. 2014, 113, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- NHS, UK. 2018. Available online: https://www.nhs.uk/conditions/cervical-cancer/ (accessed on 13 February 2019).
- Paul, J. The cancer cell in vitro: A review. Cancer Res. 1962, 22, 431–440. [Google Scholar] [PubMed]
- Leighton, J. Contributions of tissue culture studies to an understanding of the biology of cancer: A review. Cancer Res. 1957, 17, 929–941. [Google Scholar] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The cancer cell line encyclopedia enables predictive modeling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Aldasoro, C.C. The proportion of cancer-related entries in PubMed has increased considerably; is cancer truly “The emperor of all maladies”? PLoS ONE 2017, 12, e0173671. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.P.; Hansch, C. Chemical toxicity on HeLa cells. Curr. Med. Chem. 2006, 13, 423–448. [Google Scholar] [CrossRef] [PubMed]
- Masters, J.R. HeLa cells 50 years on: The good, the bad and the ugly. Nat. Rev. Cancer 2002, 2, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Lucey, B.P.; Nelson-Rees, W.A.; Hutchins, G.M. Henrietta Lacks, HeLa cells and cell culture contamination. Arch. Pathol. Lab. Med. 2009, 133, 1463–1467. [Google Scholar] [PubMed]
- Lacroix, M. Persistent use of “false” cell lines. Int. J. Cancer 2008, 122, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Denk, W.; Horstmann, H. Serial block-face scanning electron microscopy to reconstruct three-dimensional tissue nanostructure. PLoS Biol. 2004, 2, 329. [Google Scholar] [CrossRef] [PubMed]
- Peddie, C.; Collinson, L. Exploring the third dimension: Volume electron microscopy comes of age. Micron 2014, 61, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Briggman, K.L.; Bock, D.D. Volume electron microscopy for neuronal circuit reconstruction. Curr. Opin. Neurobiol. 2012, 22, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Briggman, K.L.; Denk, W. Towards neural circuit reconstruction with volume electron microscopy techniques. Curr. Opin. Neurobiol. 2006, 16, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Halbritter, F.; Vaidya, H.J.; Tomlinson, S.R. GeneProf: Analysis of high-throughput sequencing experiments. Nat. Methods 2012, 9, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Domon, B.; Aebersold, R. Challenges and opportunities in proteomics data analysis. Mol. Cell. Proteom. 2006, 5, 1921–1926. [Google Scholar] [CrossRef] [PubMed]
- Cloppet, F.; Boucher, A. Segmentation of overlapping/aggregating nuclei cells in biological images. In Proceedings of the IEEE International Conference Pattern Recognition, Tampa, FL, USA, 8–11 December 2008; pp. 1–4. [Google Scholar]
- Bajcsy, P.; Cardone, A.; Chalfoun, J.; Hlater, M.; Juba, D.; Kociolek, M.; Majurski, M.; Peskin, A.; Simon, C.; Simon, M.; et al. Survey statistics of automated segmentations applied to optical imaging of mammalian cells. BMC Bioinform. 2015, 16, 330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, H. Generalizing cell segmentation and quantification. BMC Bioinform. 2017, 18, 189. [Google Scholar] [CrossRef] [PubMed]
- Solis-Lemus, J.A.; Stramer, B.; Slabaugh, G.; Reyes-Aldasoro, C.C. Segmentation and shape analysis of macrophages using anglegram analysis. J. Imaging 2017, 4, 2. [Google Scholar] [CrossRef]
- Gurcan, M.N.; Boucheron, L.E.; Can, A.; Madabhushi, A.; Rajpoot, N.M.; Yener, B. Histopathological image analysis: A review. IEEE Rev. Biomed. Eng. 2009, 2, 147–171. [Google Scholar] [CrossRef]
- Lu, L.; Ladinsky, M.S.; Kirchhausen, T. Formation of the postmitotic nuclear envelope from extended ER cisternae precedes nuclear pore assembly. J. Cell Biol. 2011, 194, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Mahamid, J.; Pfeffer, S.; Schaffer, M.; Villa, E.; Danev, R.; Cuellar, L.K.; Forster, F.; Hyman, A.A.; Plitzko, J.M.; Baumeister, W. Visualizing the molecular sociology at the HeLa cell nuclear periphery. Science 2016, 351, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Haguenau, F.; Hawkes, P.W.; Hutchison, J.L.; Satiat-Jeunemaitre, B.; Simon, G.T. Key events in the history of electron microscopy. Microsc. Microanal. 2003, 9, 96–138. [Google Scholar] [CrossRef] [PubMed]
- Masters, B.R. History of the electron microscope in cell biology. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; pp. 1–9. [Google Scholar]
- Perez, A.; Seyedhosseini, M.; Deerinck, T.; Bushong, E.; Panda, S.; Tasdizen, T.; Ellisman, M. A workflow for the automatic segmentation of organelles in electron microscopy image stacks. Front. Neuroanat. 2014, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wilke, S.; Antonios, J.; Bushong, E.; Badkoobehi, A.; Malek, E.; Hwang, M. Deconstructing complexity: Serial block-face electron microscopic analysis of the hippocampal mossy fiber synapse. J. Neurosci. 2013, 33, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Bohorquez, D.; Samsa, L.; Roholt, A.; Medicetty, S.; Chandra, R.; Liddle, R. An enteroendocrine cell-enteric glia connection revealed by 3D electron microscopy. PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Rajapakse, J.C. Segmentation of clustered nuclei with shape markers and marking function. IEEE Trans. Biomed. Eng. 2009, 56, 741–748. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Zhou, X. Nuclei segmentation using marker-controlled watershed, tracking using mean-shift, and Kalman filter in time-lapse microscopy. IEEE Trans. Circuits Syst. 2006, 53, 2405–2414. [Google Scholar] [CrossRef]
- Kaya, B.; Goceri, E.; Beker, A.; Elder, B.; Puduvalli, V.; Winter, J.; Gurcan, M.; Otero, J.J. Automated fluorescent miscroscopic image analysis of PTBP1 expression in glioma. PLoS ONE 2017, 12, e0170991. [Google Scholar] [CrossRef]
- Goceri, E.; Goksel, B.; Elder, J.B.; Puduvalli, V.K.; Otero, J.J. Quantitative validation of anti-PTBP1 antibody for diagnostic neuropathology use: Image analysis approach. J. Numer. Methods Biomed. Eng. 2017, 33, e2862. [Google Scholar] [CrossRef]
- Beckham, C.; Hall, M.; Frank, E. WekaPyScript: Classification, regression, and filter schemes for WEKA implemented in Python. J. Open Res. Softw. 2016, 4, e33. [Google Scholar] [CrossRef]
- Luengo, I.; Darrow, M.; Spink, M.; Sun, Y.; Dai, W.; He, C.; Chiu, W.; Pridmore, T.; Ashton, A.; Duke, E.; et al. SuRVoS: Super-region volume segmentation workbench. J. Struct. Biol. 2017, 198, 43–53. [Google Scholar] [CrossRef] [PubMed]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Nurzynska, K. Deep learning as a tool for automatic segmentation of corneal endothelium images. Symmetry 2018, 10, 60. [Google Scholar] [CrossRef]
- Caicedo, J.C.; Roth, J.; Goodman, A.; Becker, T.; Karhohs, K.W.; McQuin, C.; Singh, S.; Carpenter, A.E. Evaulation of deep learning strategies for nucleus segmentation in fluorescence images. IEEE Rev. Biomed. Eng. 2018, 2, 147–171. [Google Scholar]
- Quan, T.M.; Hildebrand, D.G.C.; Jeong, W. FusionNet: A deep fully residual convolutional neural network for image segmentation in connectomics. arXiv 2016, arXiv:1612.05360. [Google Scholar]
- Kramer, N.; Haney, M.; Morgens, D.; Jovičić, A.; Couthouis, J.; Li, A.; Ousey, J.; Ma, R.; Bieri, G.; Tsui, C.; et al. CRISPR-Cas9 screens in human cells and primary neurons identify modifiers of C9 or f72 dipeptide repeat protein toxicity. Nat. Genet. 2018, 50, 603–612. [Google Scholar] [CrossRef]
- Sepulveda, G.; Antkowiak, M.; Brust-Mascher, I.; Mahe, K.; Ou, T.; Castro, N.; Christensen, L.; Cheung, L.; Jiang, X.; Yoon, D.; et al. Co-translational protein targeting facilitates centrosomal recruitment of PCNT during centrosome maturation in vertebrates. eLife 2018, 7, 1–28. [Google Scholar] [CrossRef]
- Luckner, M.; Wanner, G. From light microscopy to analytical scanning electron microscopy (SEM) and focused ion beam (FIB)/SEM in Biology: Fixed coordinates, flat embedding, absolute references. Microsc. Microanal. 2018, 24, 526–544. [Google Scholar] [CrossRef] [PubMed]
- Schnoor, J. Citizen science. Environ. Sci. Technol. 2007, 41, 5923. [Google Scholar] [CrossRef][Green Version]
- Etch a Cell. 2015. Available online: https://etchacell.crick.ac.uk/ (accessed on 13 February 2019).
- Henry, K.M.; Pase, L.; Ramos-Lopez, C.F.; Lieschke, G.J.; Renshaw, S.A.; Reyes-Aldasoro, C.C. PhagoSight: An open-source MATLABQR package for the analysis of fluorescent neutrophil and macrophage migration in a zebrafish model. PLoS ONE 2013, 8, e72636. [Google Scholar] [CrossRef] [PubMed]
- Arganda-Carreras, I.; Andrey, P. Designing image analysis pipelines in light microscopy: A rational approach. In Light Microscopy: Methods and Protocols; Markaki, Y., Harz, H., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; pp. 185–207. [Google Scholar]
- Mazzamuto, G.; Silvestri, L.; Silvestri, L.; Costantini, I.; Orsini, F.; Roffilli, M.; Frasconi, P.; Sacconi, L.; Sacconi, L.; Pavone, F.S.; et al. Software tools for efficient processing of high-resolution 3D images of macroscopic brain samples. Biophotonics Congress: Biomedical Optics Congress 2018 (Microscopy/Translational/Brain/OTS) (2018). Opt. Soc. Am. 2018. [Google Scholar] [CrossRef]
- Miura, K.; Tosi, S. Epilogue, a Framework for Bioimage Analysis. In Standard and Super-Resolution Bioimaging Data Analysis; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 269–284. [Google Scholar]
- Canny, J. A computational approach to edge detection. IEEE Trans. Pattern Anal. Mach. Intell. 1986, 8, 679–698. [Google Scholar] [CrossRef] [PubMed]
- Karabag, C.; Jones, M.L.; Peddie, C.J.; Weston, A.E.; Collinson, L.M.; Reyes-Aldasoro, C.C. Automated segmentation of HeLa nuclear envelope from electron microscopy images. In Medical Image Understanding and Analysis; Nixon, M., Mahmoodi, S., Zwiggelaar, R., Eds.; Springer International Publishing: New York, NY, USA, 2018; pp. 241–250. [Google Scholar]
- Deerinck, T.J.; Bushong, E.; Thor, A.; Ellisman, M.H. NCMIR Methods for 3D EM: A New Protocol for Preparation of Biological Specimens for Serial Block-Face SEM Microscopy; NCMIR—National Center for Microscopy and Imaging Research: La Jolla, CA, USA, 2010. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Dubuisson, M.P.; Jain, A.A. A modified Hausdorff distance for object matching. In Proceedings of the 12th IAPR International Conference on Pattern Recognition, Jerusalem, Israel, 9–13 October 1994; Volume 1, pp. 566–568. [Google Scholar]
- Maheshwari, A.; Yi, J. On computing fréchet distance of two paths on a convex polyhedron. Proceedings the 21st European Workshop on Computational Geometry, Eindhoven, The Netherlands, 9–11 March 2005; pp. 41–44. [Google Scholar]
- Chambers, E.W.; De Verdiere, E.C.; Erickson, J.; Lazard, S.; Lazarus, F.; Thite, S. Homotopic Fréchet distance between curves or, walking your dog in the woods in polynomial time. Comput. Geom. Theory Appl. 2010, 43, 295–311. [Google Scholar] [CrossRef]
- Chan, T.; Vese, L. Active contours without edges. IEEE Trans. Image Process. 2001, 10, 266–277. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell | Volume Metrics | Surface Metrics | |||||
|---|---|---|---|---|---|---|---|
| 3D Shape | Volume (μm3) | Jaccard Index | Surface Modelling against a Spheroid | Mean (μ) | Standard dev. (σ) | Range | Pixel Ratio for μ ± σ |
 | 393 | 0.5538 |  | −23.424 | 142.47 | 681 | 16% & 19% |
 | 442 | 0.6610 |  | −17.018 | 105.11 | 517 | 13% & 18% |
 | 454 | 0.6989 |  | −11.013 | 96.968 | 553 | 12% & 17% |
 | 487 | 0.7084 |  | −16.467 | 98.528 | 577 | 15% & 15% |
 | 502 | 0.6643 |  | −27.290 | 116.11 | 544 | 13% & 19% |
 | 580 | 0.5991 |  | −27.882 | 135.57 | 703 | 19% & 18% |
 | 600 | 0.5801 |  | −29.894 | 163.80 | 894 | 17% & 15% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karabağ, C.; Jones, M.L.; Peddie, C.J.; Weston, A.E.; Collinson, L.M.; Reyes-Aldasoro, C.C. Segmentation and Modelling of the Nuclear Envelope of HeLa Cells Imaged with Serial Block Face Scanning Electron Microscopy. J. Imaging 2019, 5, 75. https://doi.org/10.3390/jimaging5090075
Karabağ C, Jones ML, Peddie CJ, Weston AE, Collinson LM, Reyes-Aldasoro CC. Segmentation and Modelling of the Nuclear Envelope of HeLa Cells Imaged with Serial Block Face Scanning Electron Microscopy. Journal of Imaging. 2019; 5(9):75. https://doi.org/10.3390/jimaging5090075
Chicago/Turabian StyleKarabağ, Cefa, Martin L. Jones, Christopher J. Peddie, Anne E. Weston, Lucy M. Collinson, and Constantino Carlos Reyes-Aldasoro. 2019. "Segmentation and Modelling of the Nuclear Envelope of HeLa Cells Imaged with Serial Block Face Scanning Electron Microscopy" Journal of Imaging 5, no. 9: 75. https://doi.org/10.3390/jimaging5090075
APA StyleKarabağ, C., Jones, M. L., Peddie, C. J., Weston, A. E., Collinson, L. M., & Reyes-Aldasoro, C. C. (2019). Segmentation and Modelling of the Nuclear Envelope of HeLa Cells Imaged with Serial Block Face Scanning Electron Microscopy. Journal of Imaging, 5(9), 75. https://doi.org/10.3390/jimaging5090075