1. Introduction
Apple is a globally important fruit crop, both economically and nutritionally. The Loess Plateau is the largest apple-growing area in the world, with apple cover and yield of 1.3 million ha and 23 million tons, respectively, accounting for 25.2% of global land cover and 26.3% of global apple production in 2016 [
1]. However, the Loess Plateau is vulnerable to various environmental factors, including hailstorms, which can cause substantial damage to apple trees and their fruit. Hail damage not only impacts fruit production within the current season but also affects fruit yield in subsequent seasons by harming flower buds [
2]. Traditional anti-hail measures, such as cloud seeding, anti-hail guns, nanocomposites, or expanding planting areas have proven to be expensive and ineffective [
3]. Previous studies have indicated that hail nets can impact various environmental factors, including light, air flow, temperature, and humidity. Recently, photo-selective colored netting, a promising agro-technical approach, has emerged as an alternative solution that utilizes nets that not only offer vital protection against hail, wind, pests, and excessive solar radiation but also alter the quality of transmitted light [
4,
5]. By selectively manipulating light wavelengths, the photo-selective netting optimizes plant growth and enhances crop quality [
2,
5,
6,
7,
8,
9]. Therefore, it is crucial to understand the impact of photo-selective nets on apple tree physiology and fruit quality to effectively utilize anti-hail nets and maximize their benefits.
One of the primary advantages of photo-selective nets is their ability to reduce the amount of solar radiation reaching the orchard environment beneath them [
2]. The subtle shading effect caused by photo-selective nets can decrease leaf temperature and evaporative demand, enhancing photosynthesis and subsequently promoting carbohydrate production, potentially resulting in improved yield quality [
4,
10,
11]. Several studies have also emphasized the role of photo-selective nets in modifying the orchard environment, affecting factors such as light intensity, light quality, canopy temperature, air humidity, and soil temperature [
7,
9,
12,
13,
14]. While photo-selective nets allow solar radiation to pass through, they also scatter it, mitigating its impact [
5,
11,
13]. In a separate study conducted by Shahak et al., it was found that apple trees covered with red nets displayed a superior rate of leaf photosynthesis compared to those covered with blue, pearl, gray, and black nets [
15]. Furthermore, an investigation comparing various protective netting colors discerned that the net photosynthesis rate in ‘Fuji’ apples showed notable elevation under blue and grey nets, as opposed to pearl-colored nets [
16]. Variations in microclimatic conditions created by photo-selective nets have been found to significantly influence the physiological responses of fruit trees, which are closely linked to their growth, fruit production, and fruit quality [
2,
7,
17,
18]. In a comparative study on ‘Mondial Gala’ apples, Iglesias and Alegre reported that fruits grown under black nets exhibited significantly lower red coloration compared to those exposed to sunlight in three out of four growing seasons [
19]. Similarly, Solomakhin and Blanke discovered that apple peels under photo-selective nets had higher chlorophyll levels but four to five times lower anthocyanin levels [
20]. Furthermore, Blanke suggested the use of black nets specifically for monocolor green apple varieties and bicolor apple cultivars that require good coloration [
21].
Over the past decade, numerous field studies have consistently demonstrated that photo-selective nets have varying effects on vegetative and reproductive growth in a wide range of cultivated species, with red and yellow nets promoting vegetative growth and blue nets inducing dwarfism [
22,
23]. Conversely, gray and pearl nets have been found to effectively enhance branching in ornamental crops [
24,
25,
26]. In the context of apple cultivation, Solomakhin and Blanke observed that different types of photo-selective nets, particularly the green-black type, resulted in increased vegetative growth compared to uncovered trees [
27]. In contrast, Bastías et al. found that blue nets stimulated a higher rate of apple shoot growth compared to red, gray, and white nets [
28]. Additionally, Giaccone et al. reported an improvement in the vigor of nectarine trees when cultivated under red nets [
29].
The importance of optimal internal fruit quality is increasingly recognized by consumers, and studies have shown that the use of photo-selective nets can affect the internal quality of fruits, particularly apples [
2]. For instance, the use of black nets has been found to increase the total acidity of apples compared to those grown without any covering [
19]. The firmness of apple fruits, such as ‘Fuji’ and ‘Pinova’, is subject to variation depending on the type of photo-selective netting employed for cultivation, with apples grown under green-black and red-black netting yielding softer fruits compared to those grown under red-white nets, while the firmest fruits are produced in the control group without any netting [
20]. In a study by Do Amarante et al., it was observed that ‘Gala’ apples grown under white net exhibited a significant decrease in fruit flesh firmness at harvest, in contrast to ‘Fuji’ apples [
30]. Additionally, fruits grown under white nets showed a decrease in total soluble solids content, which was attributed to shading and resulted in reduced carbohydrate reserves in the fruit, ultimately leading to lower levels of soluble sugar at commercial maturity [
30].
This study aimed to assess the impact of four photo-selective nets (white, blue, black, and green nets) on apple orchards. As a result of the frequent hail storms in the Loess Plateau, it was impossible to utilize control plants that were exposed to direct sunlight. To evaluate the photo-selective effect of the different colors, the white net, the most used locally, was considered the control net. The evaluation encompassed various aspects, including environmental factors, growth and development indices, fruit quality, and overall yield. To determine the most effective color for orchard hail net coverage, principal component analysis was employed for a comprehensive evaluation. The research findings have significant theoretical and practical implications, providing valuable insights for improving apple tree growth, fruit yield, and quality.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
The experiment was conducted at the Apple Research Center in Luochuan County, Shaanxi Province, China (109°32′40″ E, 35°42′28″ N). The experiment was conducted over a period of two years, from June 2019 to November 2021. The orchard area employed a dwarfing rootstock-mediated high-density planting system, with 4-year-old apple trees selected as the experimental materials. The rootstock used was M26, and the cultivar was Yanfu No.8. The row spacing and plant spacing were set at 3.5 × 1.5 m.
Based on the colors of photo-selective hail nets, four treatments were established: white, blue, black, and green. The hail nets were installed at a height of 5 m above the ground in a roof-shaped structure. The installation of the nets began in April and continued until the end of November each year. The nets were made with polyethylene material by adding UV stabilizers and anti-oxidants with hed quad crossover, 4 × 7 mm mesh, 25 mm mesh size, 480 denier, and 60 gsm (Dongshen Development Ltd., Xiamen, China).
The experimental layout employed a randomized block design. A single-colored net enveloped three rows of apple trees, encompassing no less than 60 trees. The measurements were conducted on nine trees per individual colored net (treatment) within the central row to mitigate any potential border effects.
2.2. Measurement of Air Humidity, Air Temperature, Light Intensity, and Light Quality
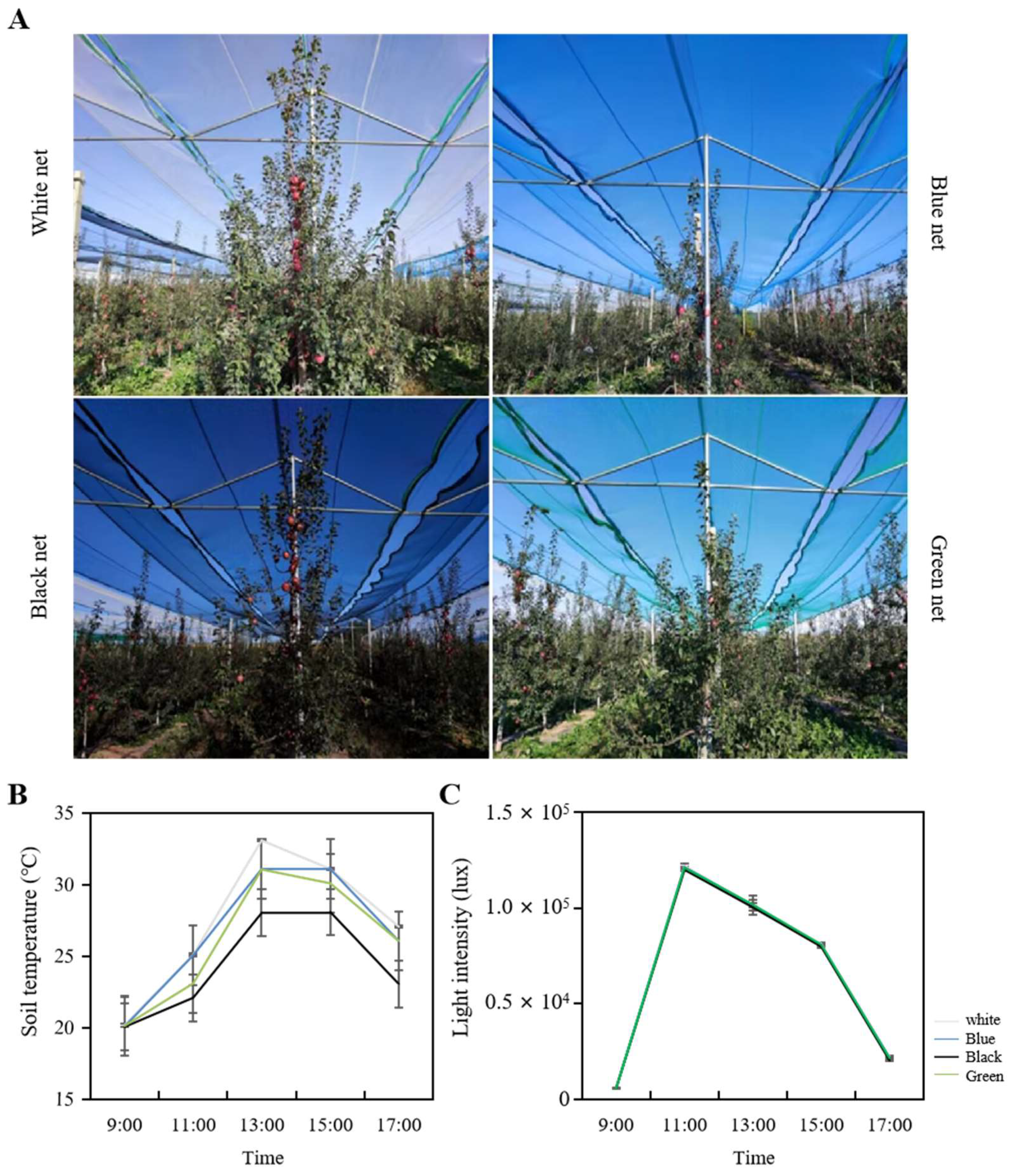
Temperature and illuminance measurements were conducted using a temperature and illuminance recorder (TPJ-22-G, Zhejiang topu yunnong Technology Co., Ltd., Hangzhou, China), as well as a spectroradiometer (HR-450, HiPoint, Taiwan, China), from 9:00 am to 5:00 pm in early August. The devices were placed at a distance of 20 cm from the outer edge of the canopy and at a height of 1.7 m above the ground, which roughly corresponded to the center of the canopy. To ensure precision and consistency, the measurements were repeated 10 times, and the obtained results were recorded for subsequent analysis.
2.3. Measurement of Soil Temperature
To measure soil temperature, a soil thermometer (TPJ-21-G, Zhejiang topu yunnong Technology Co., Ltd., Hangzhou, China) was inserted 5 cm deep and 20 cm away from the trunk. Each treatment had three biological replicates, and within each biological replicate, three trees were selected as the three basic replicates. Data changes were recorded between 9:00 am and 5:00 pm in early August.
2.4. Measurement of New Shoot Growth
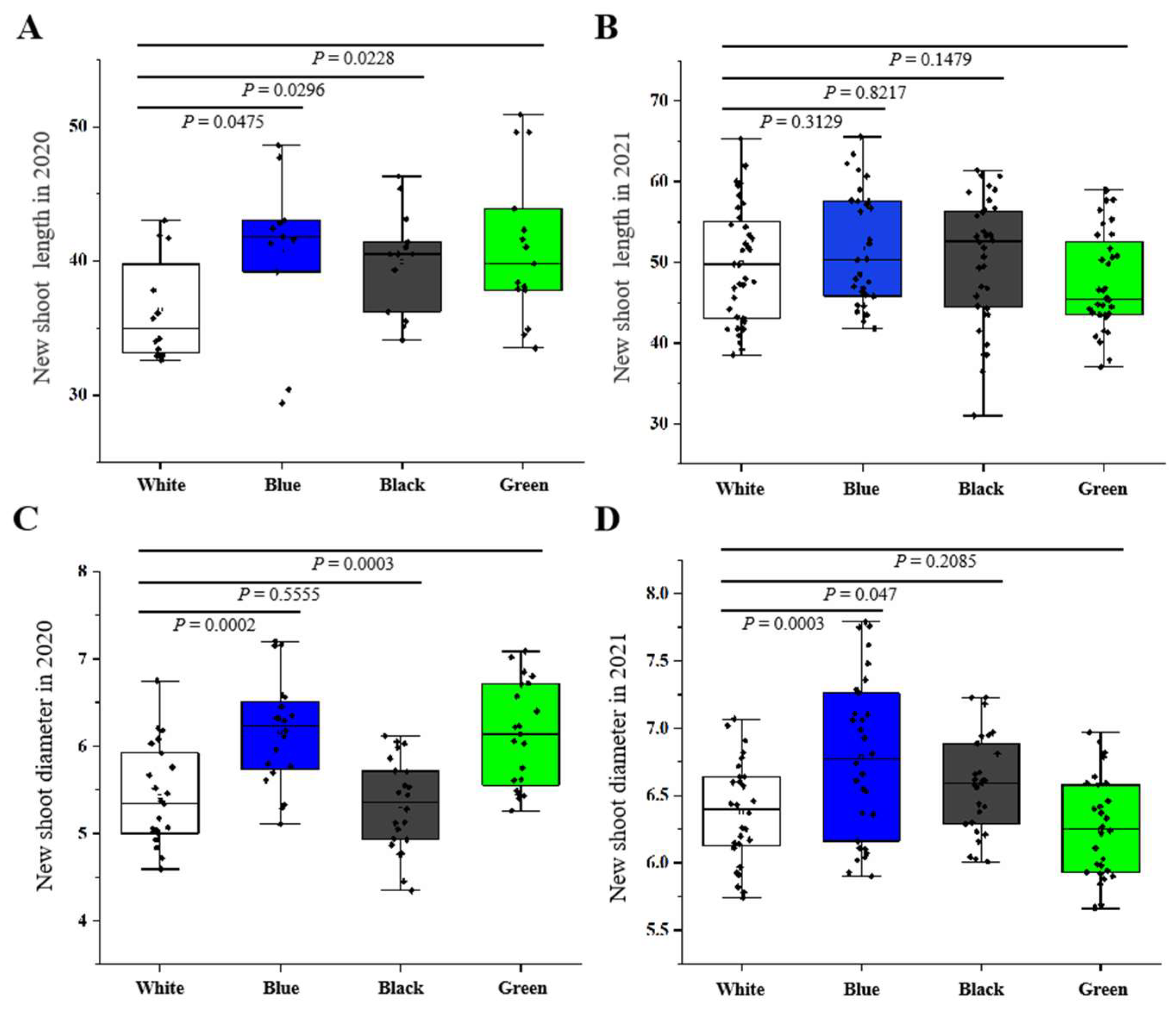
The new shoot growth was calculated by measuring the shoot length and diameter at the end of the annual vegetative growth, specifically in early August. A minimum of fifteen non-fruiting bourse shoots were selected for each treatment.
2.5. Measurement of Leaf Relative Water Content
To determine the leaf relative water content, we followed a specific procedure described previously [
31]. First, we collected fully expanded leaves and measured their weights while fresh. Next, we soaked the leaves in water for a period of 12 h and recorded their weight as saturated weight. Finally, we transferred the leaves to an oven and dried them until a constant weight (dry weight) was achieved. Fifty leaves were selected for each treatment. The relative water content was calculated using the following formula:
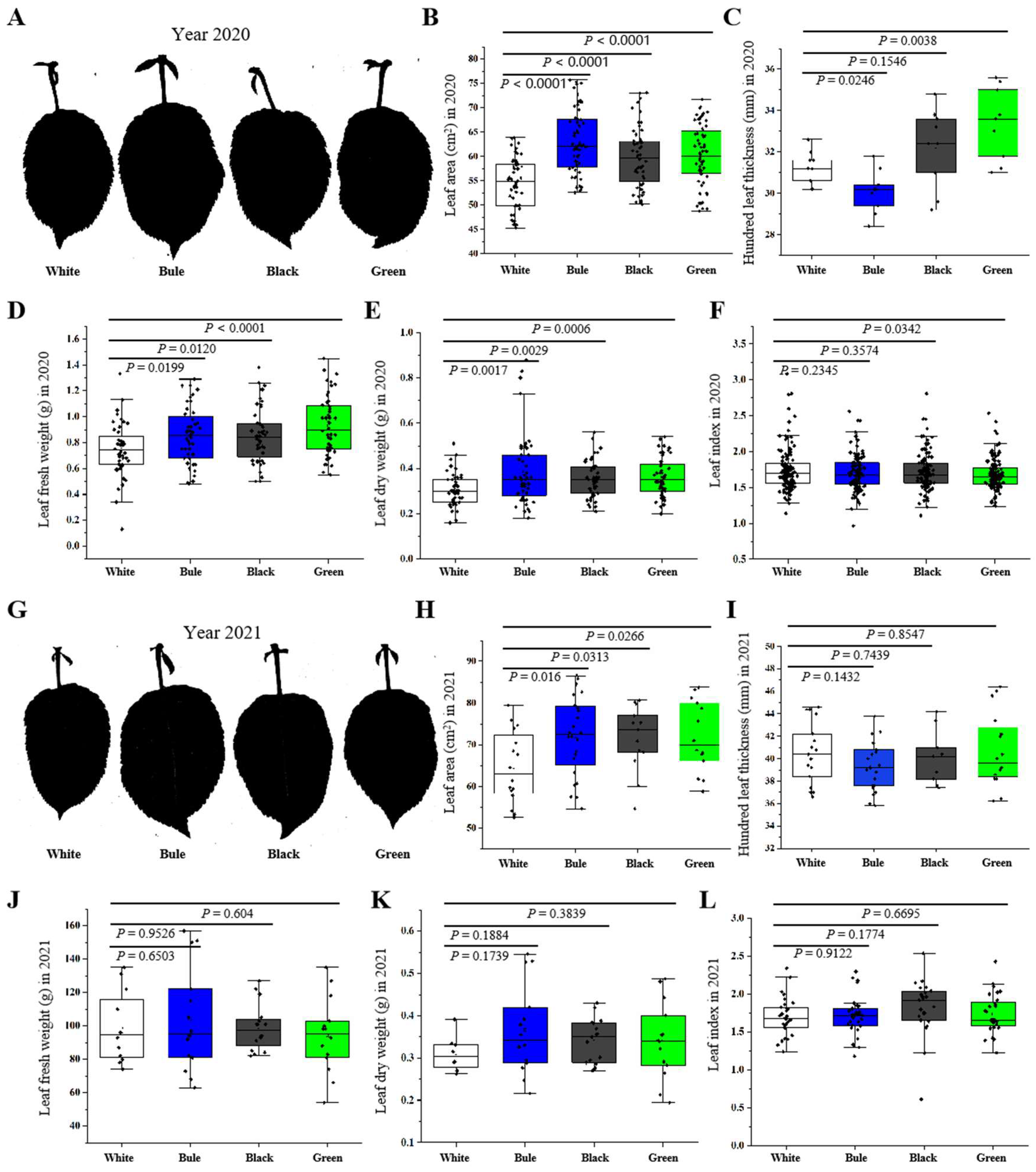
2.6. Measurement of Leaf Area
The leaves were scanned using a scanner (Epson, Suwa, Japan), and leaf auto compute software was employed for accurate calculation. Fifty leaves were selected for each treatment.
2.7. Measurement of Photosynthetic Parameters
The determination of leaf photosynthesis was conducted following the previously described methods [
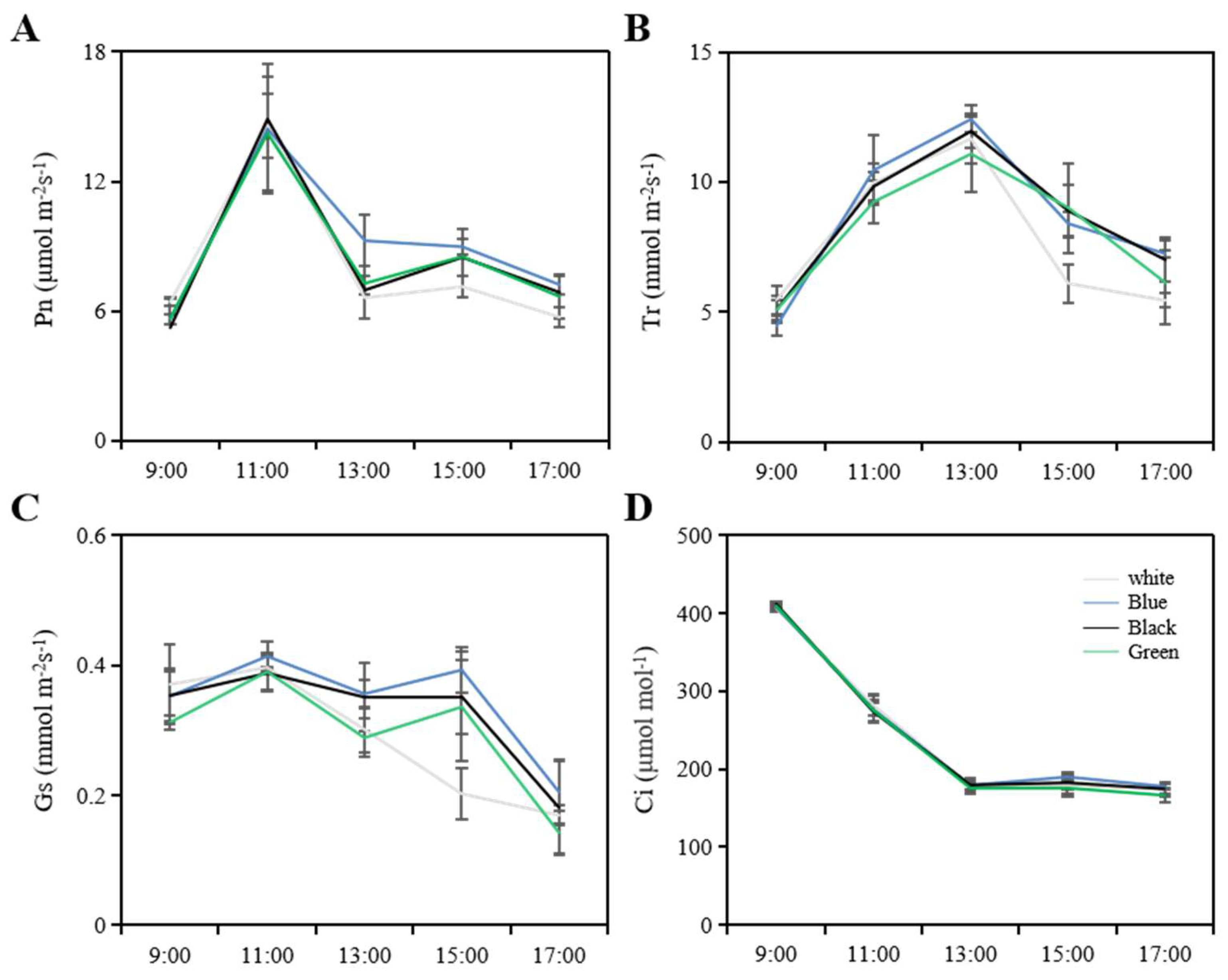
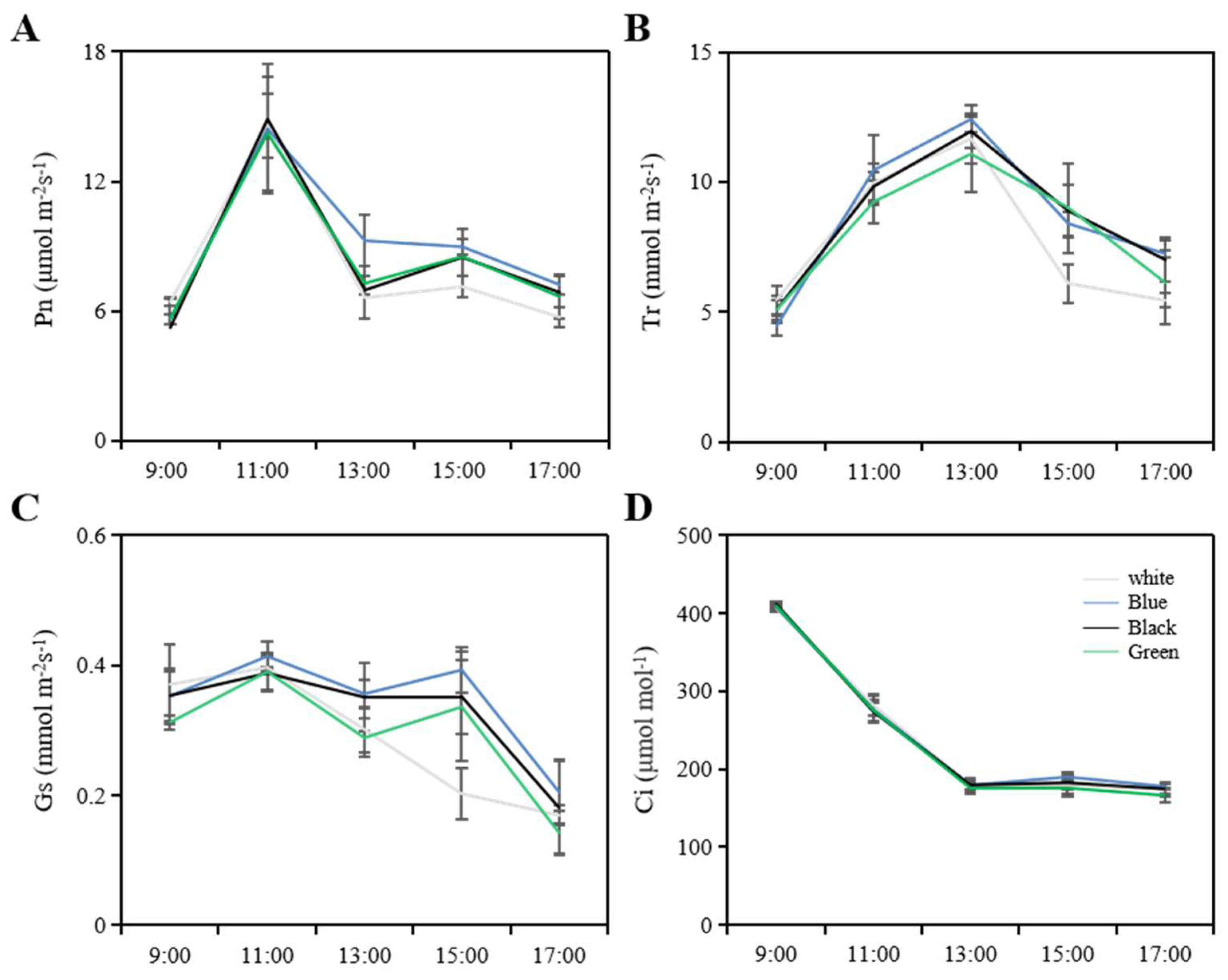
31]. During the new shoot growth period, photosynthetic parameters were measured under sunny conditions from 9:00 am to 5:00 pm. For each treatment, three branches with consistent tree vigor were selected from each biological replicate, and the sixth mature leaf from the top of each branch was used for measurement. The portable LI-6400 photosynthesis system (LI-COR, Lincoln, NE, USA) was used to measure the net photosynthetic rate, transpiration rate, intercellular carbon dioxide concentration, and stomatal conductance.
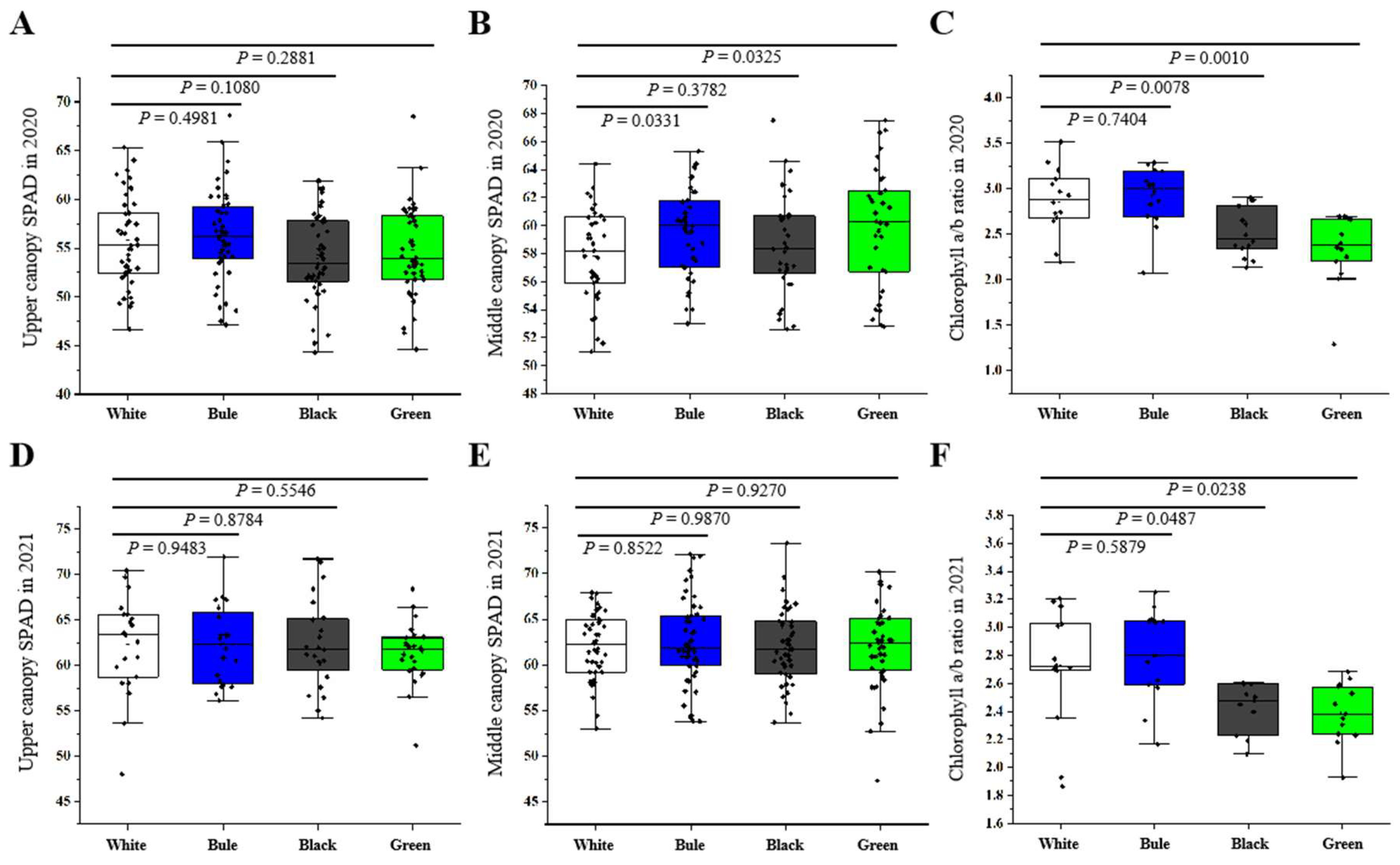
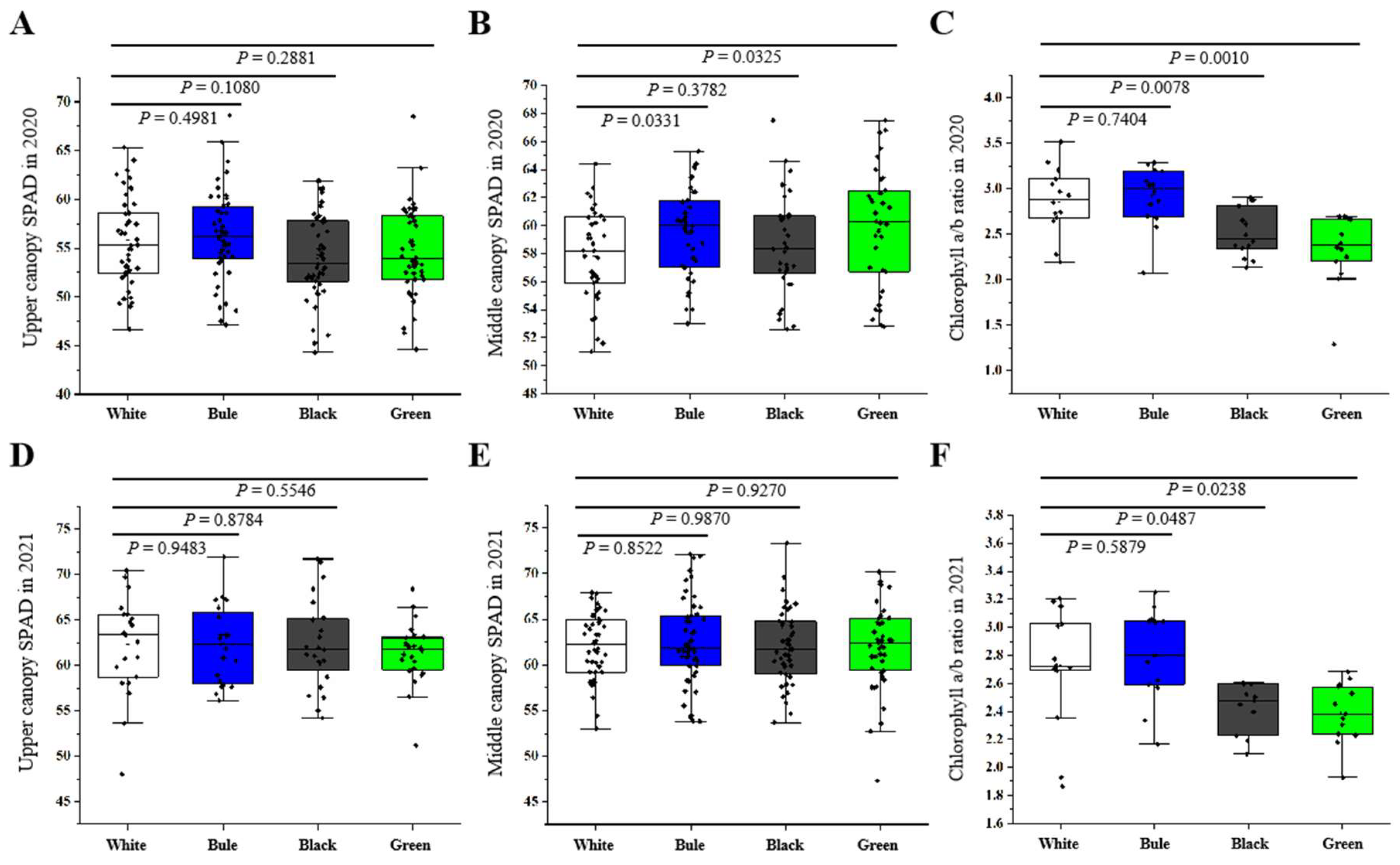
2.8. Determination of Relative Chlorophyll Content
The relative chlorophyll content of leaves in the top or mid-canopy was assessed by measuring the SPAD values of five selected leaves. The measurement of chlorophyll a and b was performed as described previously [
32]. Simply, fresh leaves were collected, and the large veins were removed. The leaves were then cut into small pieces. Approximately 0.1 g of the leaf fragments was weighed and placed in a mortar. A small amount of 80% acetone was added to the mortar, along with a pinch of calcium carbonate and quartz sand. The mixture was thoroughly ground until it became a homogeneous paste. An additional 80% acetone was added to the paste, and the resulting mixture was transferred to a centrifuge tube. The volume was adjusted to 10 mL with 80% acetone. The extraction process was carried out at room temperature in a dark place for 24 h. After the extraction, the solution was collected, and the absorbance values at wavelengths of 663 nm (A
663) and 645 nm (A
645) were measured using 80% acetone as a reference. Chlorophyll a or b was calculated using the formula:
2.9. Determination of Fruit Quality
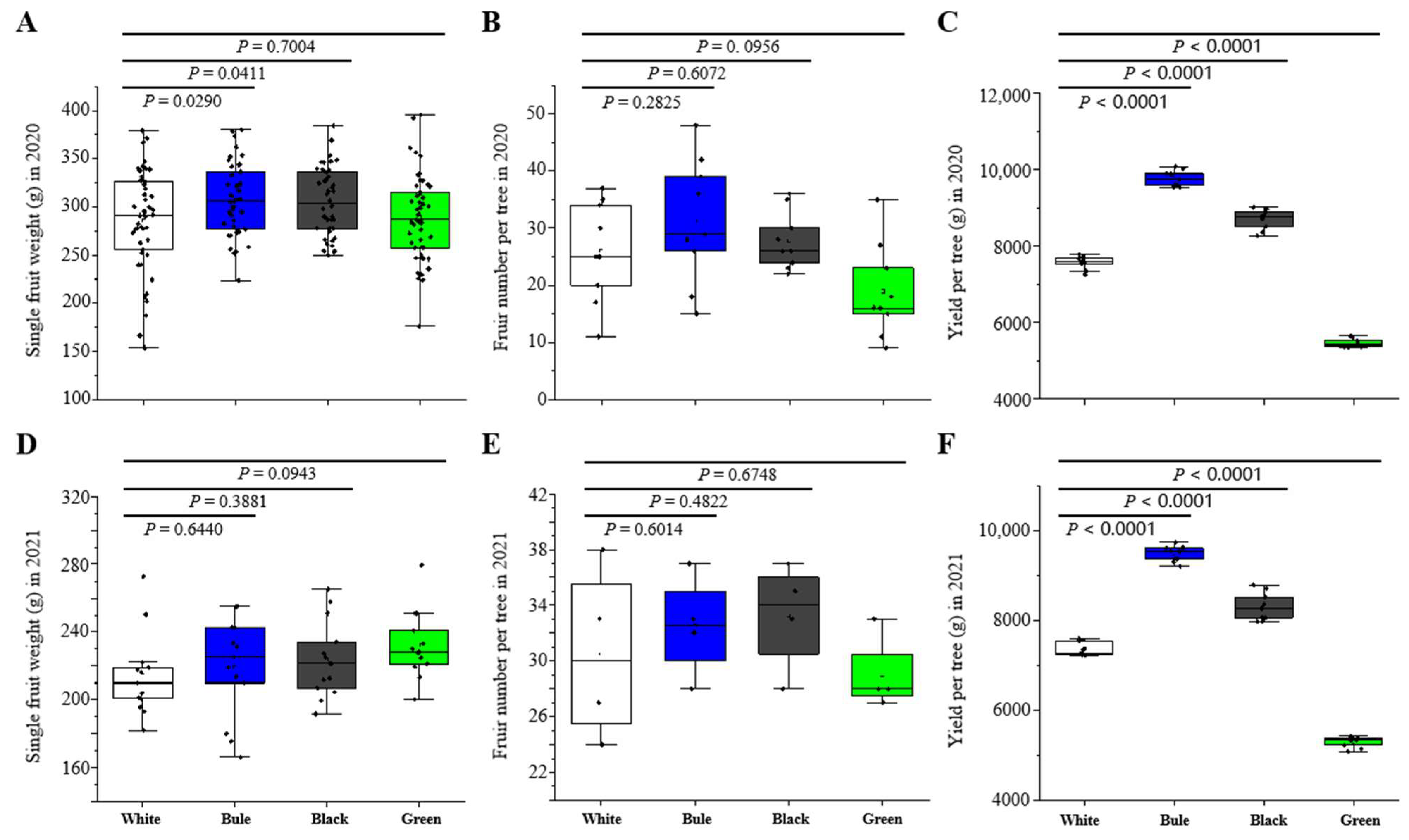
For the assessment of external quality, 15 similarly sized fruits were randomly chosen from each treatment for evaluation. Parameters such as fruit weight, shape, and skin color were measured. An electronic vernier caliper was used to measure the maximum longitudinal and transverse diameters of the fruit. A ratio of these diameters was then used to define the shape of the fruit. A portable Cr-100 colorimeter (X-Rite, Granville, MI, USA) was employed to measure skin color parameters. Variations in skin color were denoted using brightness (L*), red-greenness (a*), and yellow-blueness (b*) values. To compute the yield per plant, these fruits were harvested.
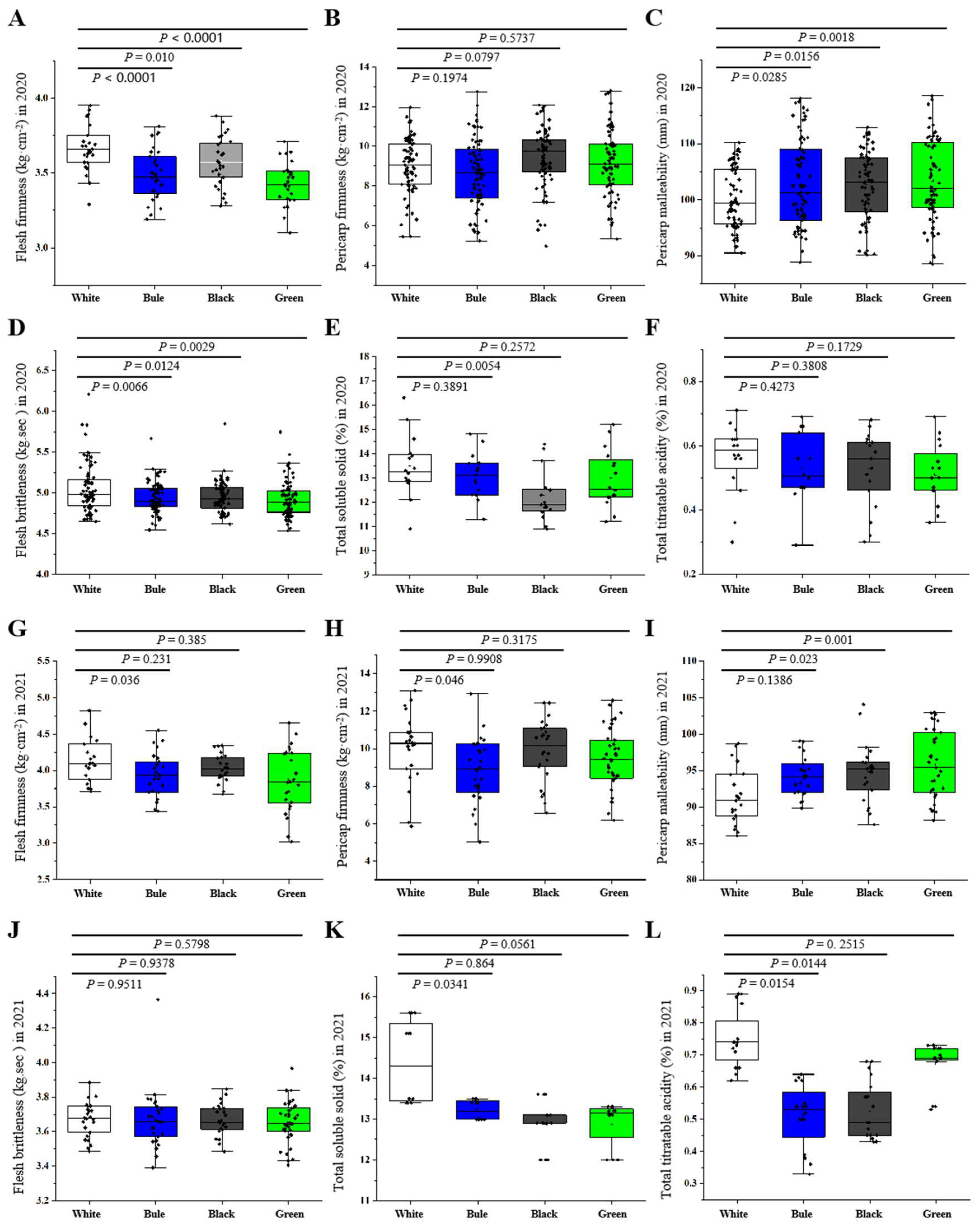
To evaluate internal quality, various parameters were measured, including flesh firmness, pericarp firmness, pericarp malleability, and flesh brittleness. These assessments were carried out at five distinct points on the fruit’s equatorial surface using a fruit texture analyzer (TMS-Touch, FTC, Frederick, MD, USA). Subsequently, these individual measures were averaged to yield a single value for each parameter. Additional measurements included soluble solid content gauged using a PAL-1 digital refractometer (Atago, Tokyo, Japan). Lastly, the fruit’s acidity level was determined with the use of a digital GMK-835F device (G-WON, Seongnam-si, Republic of Korea).
2.10. Principal Component Analysis
We conducted dimensionality reduction and principal component analysis on the data from 2020 and 2021 using IBM SPSS Statistics 20. The correspondence between factors and items was determined by analyzing the factor loading coefficient matrix table after rotation. A factor loading coefficient with an absolute value greater than 0.4 indicates a significant relationship between the item and the dimension (factor). In cases where a research item corresponds to multiple factors, professional knowledge is taken into account to determine its specific attribution to a particular factor.
2.11. Statistical Analysis
All statistical analyses were performed using Origin 2019b software. The significance of differences between treatments for the various measured parameters was evaluated through one-way ANOVA Tukey’s test.
4. Discussion
Apple crops are increasingly being cultivated under protective netting systems, providing protection against extreme weather events [
6]. Technological advancements have facilitated the development of colored nets equipped with photo-selective plastic filters. These nets not only offer differential filtration of solar radiation and physical protection but also drastically alter the light conditions, notably the spectral light composition [
37]. Plant perception of light is affected by both its intensity and spectral characteristics. [
2]. According to our research findings, these photo-selective nets do not noticeably influence light intensity indicators, a result that is consistent with the findings by Bastías et al. that red and blue nets curtailed the photosynthetically active radiation equivalently as compared to the white net [
28]. In a related study, Serra et al. found that apple trees cultivated under photo-selective nets intercepted more light than their uncovered counterparts. Nevertheless, the net’s color had no significant effect on the tree’s light interception over the course of two years [
38]. In addition, several studies have demonstrated the direct impact of photo-selective nets on the transmission spectra of sunlight. Specifically, the blue net was found to reduce transmission in the 600–720 nm range, while increasing transmission in the blue and blue-green wavelengths, particularly within the 440–520 nm range [
39,
40]. Consistent with these findings, our research revealed that the blue net increased the proportion of blue light while significantly decreasing the proportion of red and far-red light, and the red to far-red light ratio. Similarly, the black and green nets in our study partially reduced the proportion of red and far-red light. In conclusion, it appears that photo-selective nets modify the spectrum of light that reaches the orchard.
Photo-selective nets have been found to alter the microenvironment of orchards, leading to changes in soil temperature [
2,
14,
19]. The decrease in soil temperature is largely due to the decreased amount of light that penetrates the ground as a result of the shade net [
12,
41]. In our study, we found the black net to be especially efficient in reducing soil temperature when compared with other colors, such as white, blue, and green. These results are consistent with findings by Kalcsits et al., where soil temperatures under pearl and blue nets were significantly lower than those under uncovered control and red nets [
14]. Although there is no documented evidence regarding the varying impact of photo-selective netting on soil temperature in the prevalent literature, we speculate that there may be other characteristics of the net’s transmittance spectrum that influence soil temperature. However, further research is required to explore this possibility.
Multiple studies have proposed that photo-selective nets not only partially transmit solar radiation but also diffuse it, which is essential for the photosynthesis of leaves in the lower part of the canopy [
8,
11]. A comparison has shown that the net photosynthesis in ‘Fuji’ apples was considerably higher under blue and grey nets than those grown under pearl net [
16]. Similar patterns have also been observed in ornamental plants as well [
42]. Our research supports these findings and further reveals that photosynthesis was most effective under the blue net, followed by the black and green nets, with the white net demonstrating the lowest efficiency. We also noted a slight increase in SPAD values when measurements were taken under the blue net in comparison to the white net. This could potentially be attributed to the enhanced absorption capacity of the primary photosynthetic pigments (chlorophylls a/b) in the blue spectrum. Moreover, our study uncovered a positive correlation between the blue net and higher stomatal conductance, leading to an increase in photosynthesis. This observation aligns with evidence obtained from research on ornamental plants, which states that blue light wavelength is more effective in triggering stomatal opening and inhibiting stomatal closure [
43]. Thus, the elevated stomatal conductance observed in leaves grown under the blue net in our research can be attributed to the effect of blue light on the stomatal aperture.
One crucial organ for analyzing crop growth is the leaf, as it helps us comprehend the crop’s ability to convert radiation into dry matter via photosynthesis [
44]. Microscopy analysis has shown that under blue net, both the palisade thickness and the ratio of palisade to spongy mesophyll decreased by 19% compared to leaves under white net [
45]. It is important to note that earlier studies have underlined the importance of a low red to far-red (R/FR) ratio in governing plant growth. This not only stimulates an enlargement of the cell wall, leading to an increase in leaf area, but also improves leaf photosynthetic capacity, dry matter accumulation, and overall plant growth [
46,
47,
48]. Our research affirms these findings, as it revealed an increase in leaf area and a reduction in leaf thickness under blue net, suggesting that these are adaptive responses for shaded leaves to maximize light transmission to the chloroplasts. Moreover, our results are in line with the significantly lower R/FR ratios observed under blue netting, reinforcing the role of the R/FR ratio in facilitating these responses. Adjusting the R/FR ratio has been recommended as a strategy for managing shoot extension, particularly to encourage greater shoot length under low R/FR ratios [
13]. Previous investigations have consistently demonstrated that a reduction in the R/FR ratio leads to shoot elongation across various plant species, such as kiwifruit [
49], grapevines [
50], peach [
29], and ornamental plants [
51]. Similarly, our study found that trees grown under blue net with a low R/FR ratio exhibited significantly greater total shoot length and roughness compared to those grown under white net. These effects may be the result of responses induced by the phytochrome, which are triggered by the lowered R/FR ratio and the consequent decrease in phytochrome photo-equilibrium as observed under blue net.
Fruit color is a critical aspect that influences consumers’ fruit consumption decisions [
52]. Our study revealed that diversely colored nets have no significant impact on fruit color or fruit shape index. These observations align with most previous studies, which reported minimal or no effects of netting on apple fruit color and shape [
5,
39,
53]. However, it is important to note that the influence of netting can differ based on various factors, including the type of net and apple variety.
Consumer preference for apples hinges heavily on their sweetness, which is generally determined by the total soluble solids (TSS) content [
54]. In three growing seasons, the use of a black shade net significantly reduced TSS in ‘Mondial Gala’ apples compared to both a crystal shade net and a control group without any shade. However, these differences were not apparent in another growing season [
19]. Similarly, for ‘Elstar’ apples, both white and black shade nets resulted in a reduction of TSS compared to the control group [
55]. Do Amarante et al. also reported a significant reduction in TSS for ‘Gala’ apples grown under a white shade net, a phenomenon not observed in ‘Fuji’ apples during harvest [
30]. Our study found that apples grown under blue nets presented a decrease in TSS accompanying a significant decrease in titratable acidity compared to white nets. These observations indicate the influence of net color on soluble solids and titratable acidity, potentially due to the modulation of light diffusion. This further emphasizes the importance of integrating photo-selective nets in apple cultivation practices, as they markedly affect the levels of soluble solids and titratable acidity in apples.
While consumers initially judge a product by its appearance, their ultimate decision to repurchase it is based on its edible quality [
56]. High initial firmness values at harvest can extend the duration of flesh firmness retention [
57]. Prior research suggests that consumers favor firmer apples [
58]. Compared to those cultivated under red-white nets, “Fuji” and “Pinova” apples grown under green-black and red-black netting were found to have a softer texture. Fruit from the control group, not covered by any netting, displayed the highest firmness [
20]. Even though limited research has been conducted on pericarp malleability, pericarp firmness, and flesh brittleness, these factors are closely associated with postharvest storage quality. They are critical for the long-term storage of fruits. However, the impact of photo-selective netting on pericarp firmness, malleability, and flesh brittleness at both pre- and post-harvest stages remains largely unexplored. In this study, the use of colored nets resulted in a reduction in flesh firmness, suggesting a potentially negative effect of photo-selective netting on either postharvest fruit storage or consumer purchasing behavior.
The illumination conditions created by photo-selective nets can influence plant physiology, thereby affecting both the average fruit weight and plant yield [
59]. Previous studies found that the use of blue or grey netting significantly increased the weight of ‘Fuji’ apples compared to a control group using white netting [
28]. Likewise, cucumbers grew heavier when under aluminized, pearl, blue, or red nets [
60]. Consistent with these results, our study showed that blue nets yielded heavier individual fruits compared to white nets. Additionally, research has indicated that prolonged exposure to blue light may improve photosystem II, stomatal conductance, and dry matter production [
61,
62]. Therefore, adjusting the combination of blue, red, and far-red light using photo-selective nets could manipulate the processes controlling carbohydrate availability, which is crucial for apple growth and yield. Our study also observed a significant rise in apple yield when using blue netting instead of white nets, supporting previous studies by Hemming et al. and Zheng et al., who reported that shading nets that enhance diffuse light can improve fruit yield in horticultural crops by increasing plant photosynthetic capacity [
63,
64].


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}