

Enhancing Post-Harvest Resilience: Investigating the Synergistic Effects of Essential Oil Combinations on Biochemical Profiles in Botrytis cinerea-Infected Apples


Abstract
1. Introduction
2. Materials and Methods
2.1. Fruit Materials, Pathogen, Chemicals
2.2. Inoculation and Storage of Fruits
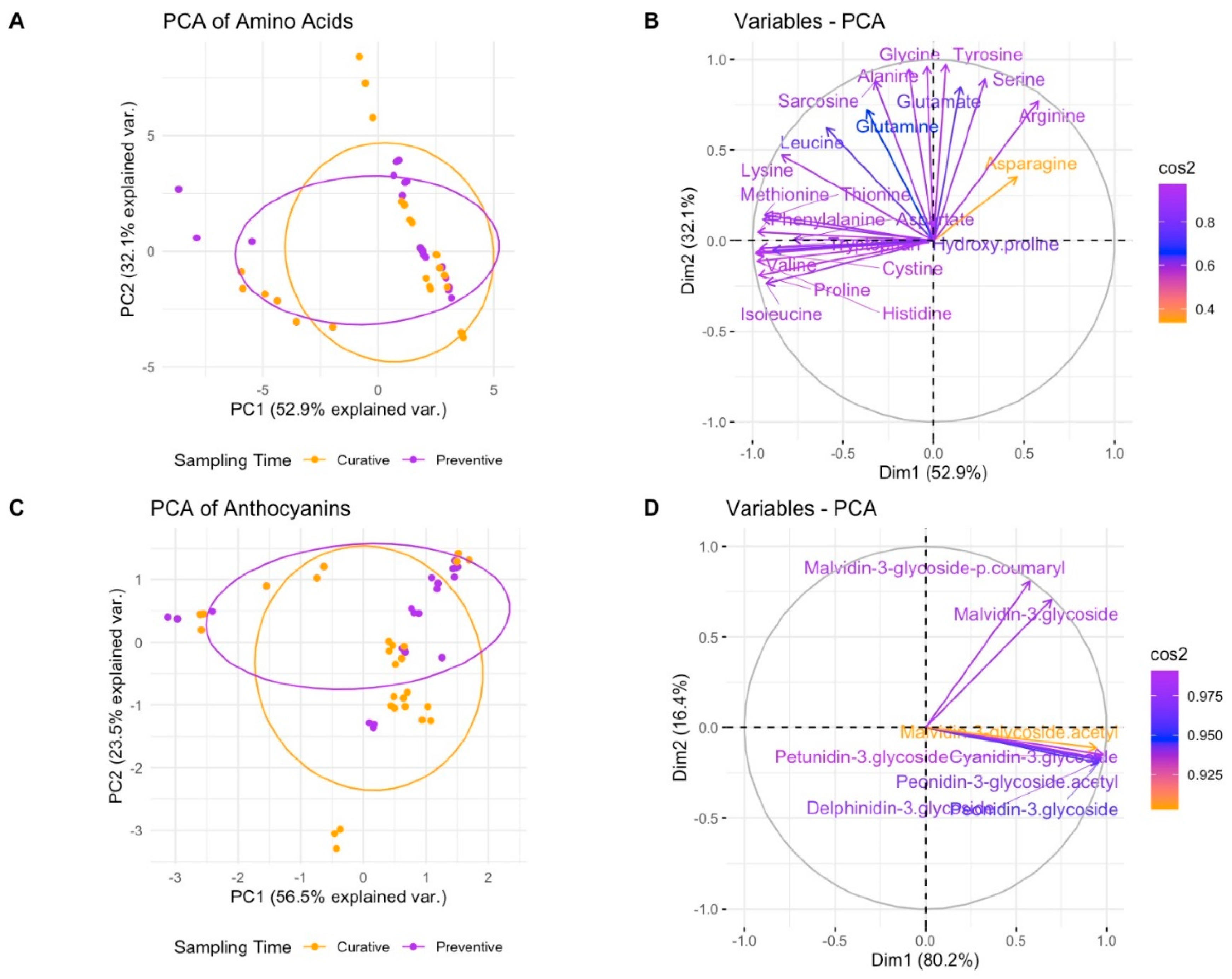
2.3. Identification of Amino Acid Profiling in Apple by HPLC
2.4. Identification of Anthocyanins Profiling in Apple by HPLC
2.5. Identification of Organic Acids from Apple by HPLC
2.6. Identification of Phenolic Compounds from Apple by HPLC
2.7. Identification of Hormones from Apple by HPLC
2.8. Data Analysis
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hirst, K.K. The History and Health Benefits of Eating Apples. 2017, Verywell Fit. Available online: https://www.verywellfit.com/the-history-and-health-benefits-of-eating-apples-505977 (accessed on 15 May 2023).
- Garcia-Mazcorro, J.F.; Pedreschi, R.; Yuan, J.; Kawas, J.R.; Chew, B.; Dowd, S.E.; Noratto, G. Apple consumption is associated with a distinctive microbiota, proteomics and metabolomics profile in the gut of Dawley Sprague rats fed a high-fat diet. PLoS ONE 2019, 14, e0212586. [Google Scholar] [CrossRef]
- Holmberg, T.; Logander, M.; Lindqvist, F. “Living on the Edge”—A Case Study of Important Factors for the Survival of Apple Computers, Inc. Master’s Thesis, Lund University, Lund, Sweden, 2005. [Google Scholar]
- Keesing, R.G. The history of Newton’s apple tree. Contemp. Phys. 1998, 39, 377–391. [Google Scholar] [CrossRef]
- Gulsen, O.; Kaymak, S.; Ozongun, S.; Uzun, A. Genetic analysis of Turkish apple germplasm using peroxidase gene-based markers. Sci. Hortic. 2010, 125, 368–373. [Google Scholar] [CrossRef]
- Malladi, A. Molecular physiology of fruit growth in apple. Hortic. Rev. 2020, 47, 1–42. [Google Scholar]
- Aydın, B.; Aktürk, D.; Özkan, E.; Hurma, H.; Kiracı, M.A. Comparative energy use efficiency and economic analysis of apple production in Turkey: Case of Thrace Region. Erwerbs-Obstbau 2019, 61, 39–45. [Google Scholar] [CrossRef]
- Gil, K.A.; Wojdyło, A.; Nowicka, P.; Montoro, P.; Tuberoso, C.I.G. Effect of apple juice enrichment with selected plant materials: Focus on bioactive compounds and antioxidant activity. Foods 2023, 12, 105. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Dietary anthocyanin-rich plants: Biochemical basis and recent progress in health benefits studies. Mol. Nutr. Food Res. 2012, 56, 159–170. [Google Scholar] [CrossRef]
- Guiné, R.P.; Barroca, M.J.; Coldea, T.E.; Bartkiene, E.; Anjos, O. Apple fermented products: An overview of technology, properties and health effects. Processes 2021, 9, 223. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, B.; Liu, J.; Kuang, Y. Plant hormones in aging: An apples-to-humans comparison. Aging Cell 2019, 18, e12837. [Google Scholar]
- Whiley, H.; Gaskin, S.; Schroder, T.; Ross, K. Antifungal properties of essential oils for improvement of indoor air quality: A review. Rev. Environ. Health 2018, 33, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Tuyet, B. Preharvest Conditions Affecting Apple Quality, Antioxidant Responses and Susceptibility to the Infection by Grey mould (Botrytis cinerea). Ph.D. Thesis, Department of Chemistry and Molecular Biology, University of Gothenburg, Gothenburg, Sweden, 2020. Available online: http://hdl.handle.net/2077/63731 (accessed on 20 May 2023).
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef]
- Yelboğa, B.; Karakuş, S. A study on antifungal activity of thymol, eugenol, and 1,8-cineole against Botrytis cinerea Persoon isolated from grapevine (Vitis vinifera Linné). J. Cent. Eur. Agric. 2023, 24, 888–898. [Google Scholar] [CrossRef]
- Shao, X.; Wang, H.; Xu, F.; Cheng, S. Effects and possible mechanisms of tea tree oil vapor treatment on the main disease in postharvest strawberry fruit. Postharvest Biol. Technol. 2013, 77, 94–101. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; El Ghaouth, A.; Wilson, C. Influence of food additives on the control of postharvest rots of apple and peach and efficacy of the yeast-based biocontrol product Aspire. Postharvest Biol. Technol. 2003, 27, 127–135. [Google Scholar] [CrossRef]
- Barrado, E.; Rodriguez, J.A.; Castrillejo, Y. Determination of primary amino acids in wines by high-performance liquid magneto-chromatography. Talanta 2009, 78, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Yousef, G.G.; Brown, A.F.; Funakoshi, Y.; Mbeunkui, F.; Grace, M.H.; Ballington, J.R.; Loraine, A.; Lila, M.A. Efficient quantification of the health-relevant anthocyanin and phenolic acid profiles in commercial cultivars and breeding selections of blueberries (Vaccinium spp.). J. Agric. Food Chem. 2013, 61, 4806–4815. [Google Scholar] [CrossRef]
- Keskin, N.; Kaya, O.; Ates, F.; Turan, M.; Gutiérrez-Gamboa, G. Drying grapes after the application of different dipping solutions: Effects on hormones, minerals, vitamins, and antioxidant enzymes in Gök Üzüm (Vitis vinifera L.) raisins. Plants 2022, 11, 529. [Google Scholar] [CrossRef]
- Kojima, K.; Ikarashi, H.; Andou, D.; Matsumoto, T. Endogenous Plant Hormone Profiles in Growing Campbell Early Grape Berries. Hortic. J. 2020, 89, 509–515. [Google Scholar] [CrossRef]
- Kaya, O.; Ates, F.; Kara, Z.; Turan, M.; Gutiérrez-Gamboa, G. Study of Primary and Secondary Metabolites of Stenospermocarpic, Parthenocarpic, and Seeded Raisin Varieties. Horticulturae 2022, 8, 1030. [Google Scholar] [CrossRef]
- Gholamnezhad, J. Effect of plant extracts on activity of some defense enzymes of apple fruit in interaction with Botrytis cinerea. J. Integr. Agric. 2019, 18, 115–123. [Google Scholar] [CrossRef]
- Abd-Elkader, D.Y.; Salem, M.Z.; Komeil, D.A.; Al-Huqail, A.A.; Ali, H.M.; Salah, A.H.; Hassan, H.S. Post-harvest enhancing and Botrytis cinerea control of strawberry fruits using low cost and eco-friendly natural oils. Agronomy 2021, 11, 1246. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Gorni, P.H.; Pacheco, A.C.; Moro, A.L.; Silva, J.F.A.; Moreli, R.R.; de Miranda, G.R.; da Silva, R.M.G. Salicylic acid foliar application increases biomass, nutrient assimilation, primary metabolites and essential oil content in Achillea millefolium L. Sci. Hortic. 2020, 270, 109436. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Minhas, P.S. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Ou-Ani, O.; Moujane, S.; Oucheikh, L.; Youssefi, Y.; Ansari, A.; Oubair, A.; Znini, M. Essential oil from Teucrium luteum subsp. flavovirens: Optimization of yield extraction using response surface methodology, molecular docking and biocontrol against apple gray mold caused by Botrytis cinerea. Arch. Phytopathol. Plant Prot. 2022, 55, 1504–1529. [Google Scholar] [CrossRef]
- Mbili, N.C. Management of Postharvest Diseases of Apples Using Essential Oils. Doctoral Dissertation, Stellenbosch University, Stellenbosch, South Africa, 2015. [Google Scholar]
- Grande-Tovar, C.D.; Chaves-López, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan coatings enriched with essential oils: Effects on fungi involved in fruit decay and mechanisms of action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Freires, I.D.A.; Murata, R.M.; Furletti, V.F.; Sartoratto, A.; Alencar, S.M.D.; Figueira, G.M.; Rosalen, P.L. Coriandrum sativum L. (coriander) essential oil: Antifungal activity and mode of action on Candida spp., and molecular targets affected in human whole-genome expression. PLoS ONE 2014, 9, e99086. [Google Scholar]
- Behshti, M.; Jahani, M.; Aminifard, M.H.; Hosseini, S.A. Essential oils to control Botrytis cinerea in vitro and in vivo on grape fruits. J. Hortic. Postharvest Res. 2020, 3, 161–172. [Google Scholar]
- Long, H.; Bi, Y.; Pu, L.; Xu, W.; Xue, H.; Fu, G.; Prusky, D. Preparation of chitosan/fennel seed essential oil/starch sodium octenyl succinate composite films for apple fruit preservation. LWT 2022, 167, 113826. [Google Scholar] [CrossRef]
- Fanyuk, M.; Kumar Patel, M.; Ovadia, R.; Maurer, D.; Feygenberg, O.; Oren-Shamir, M.; Alkan, N. Preharvest application of phenylalanine induces red color in mango and apple fruit’s skin. Antioxidants 2022, 11, 491. [Google Scholar] [CrossRef]
- Ghanati, F.; Bakhtiarian, S.; Parast, B.M.; Behrooz, M.K. Production of new active phytocompounds by Achillea millefolium L. after elicitation with silver nanoparticles and methyl jasmonate. Biosci. Biotechnol. Res. Asia 2014, 11, 391–399. [Google Scholar] [CrossRef]
- Sivakumar, D.; Bill, M.; Korsten, L.; Thompson, K. Integrated application of chitosan coating with different postharvest treatments in the control of postharvest decay and maintenance of overall fruit quality. In Chitosan in the Preservation of Agricultural Commodities; Academic Press: Cambridge, MA, USA, 2016; pp. 127–153. [Google Scholar]
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Mekawi, E.M.; Khafagi, E.Y.; Abdel-Rahman, F.A. Effect of pre-harvest application with some organic acids and plant oils on antioxidant properties and resistance to Botrytis cinerea in pepper fruits. Sci. Hortic. 2019, 257, 108736. [Google Scholar] [CrossRef]
- Bika, R.; Baysal-Gurel, F.; Jennings, C. Botrytis cinerea management in ornamental production: A continuous battle. Can. J. Plant Pathol. 2021, 43, 345–365. [Google Scholar] [CrossRef]
- Rguez, S.; Hamrouni Sellami, I.; Abid, G.; Papetti, A.; Hammami, M.; Souissi, F.; Djébali, N. Tetraclinis articulata essential oil emulsion use as an alternative to chemical fungicide to control tomato grey mould disease. Int. J. Environ. Health Res. 2023, 34, 1100–1112. [Google Scholar] [CrossRef]
- Khursheed, A.; Rather, M.A.; Jain, V.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant-based natural products as potential eco-friendly and safer biopesticides: A comprehensive overview of their advantages over conventional pesticides, limitations and regulatory aspects. Microb. Pathog. 2022, 173, 105854. [Google Scholar] [CrossRef] [PubMed]
- Vega-Vásquez, P.; Mosier, N.S.; Irudayaraj, J. Hormesis-inducing essential oil nanodelivery system protects plants against broad host-range necrotrophs. ACS Nano 2021, 15, 8338–8349. [Google Scholar] [CrossRef]
- Pusztahelyi, T.; Holb, I.J.; Pócsi, I. Secondary metabolites in fungus-plant interactions. Front. Plant Sci. 2015, 6, 145754. [Google Scholar] [CrossRef] [PubMed]
- Risuleo, G.; La Mesa, C. Resveratrol: Biological activities and potential use in health and disease. In Nutraceuticals in Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2019; pp. 215–226. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Applications (A) X | Aspartate | Glutamate | Asparagine | Serine | Glutamine | Histidine | Glycine | Thionine | Arginine | Alanine | ||
| Preventive | 109 ± 1.1 | 200 ± 3.2 | 322 ± 5.3 a | 381 ± 8.8 b | 292 ± 6.3 b | 271 ± 11.3 b | 116 ± 2.2 b | 328 ± 10.9 | 275 ± 6.0 a | 274 ± 3.4 | ||
| Curative | 111 ± 2.1 | 201 ± 4.1 | 535 ± 4.5 b | 352 ± 6.8 a | 251 ± 8.3 a | 188 ± 10.3 a | 97 ± 2.9 a | 305 ± 11.8 | 327 ± 7.1 b | 267 ± 6.1 | ||
| Treatments (T) Y | ||||||||||||
| C | 189.8 ± 4.1 d | 96.7 ± 8.3 a | 98.8 ± 10.1 a | 189.8 ± 3.5 d | 260 ± 17.7 bc | 424.3 ± 4.1 b | 56.2 ± 6.2 a | 427.1 ± 22.3 b | 53.3 ± 11.9 a | 204.0 ± 9.5 a | ||
| F | 185.2 ± 3.5 d | 147.2 ± 7.8 b | 133.3 ± 9.7 a | 185.2 ± 4.1 d | 285 ± 17.7 bc | 643.8 ± 3.5 a | 70.4 ± 5.1 ab | 503.2 ± 20.1 b | 67.1 ± 13.3 a | 227.8 ± 12.9 a | ||
| T+F | 128.6 ± 4.4 c | 318.2 ± 5.2 f | 515.1 ± 11.1 d | 128.6 ± 4.2 c | 426 ± 17.7 d | 471.6 ± 4.3 b | 201.7 ± 6.1 e | 479.8 ± 19.3 b | 377.4 ± 14.9 bc | 423.1 ± 6.9 c | ||
| C+F | 79.3 ± 2.2 ab | 209.1 ± 8.3 cde | 351.0 ± 11.4 bc | 79.3 ± 5.1 ab | 270 ± 17.7 bc | 98.2 ± 4.2 a | 102.0 ± 7.2 c | 230.0 ± 17.1 a | 353.0 ± 15.0 bc | 256.3 ± 4.1 ab | ||
| E+F | 95.2 ± 4.3 b | 240.3 ± 5.5 e | 529.0 ± 10.7 d | 95.2 ± 3.5 b | 307 ± 17.7 c | 111.9 ± 3.5 a | 162.5 ± 3.3 d | 276.2 ± 25.3 a | 458.5 ± 11.9 d | 316.8 ± 9.9 b | ||
| C+E+F | 82.8 ± 4.1 ab | 192.6 ± 6.8 cd | 690.0 ± 9.7 e | 82.8 ± 2.9 ab | 208 ± 17.7 ab | 66.3 ± 4.1 a | 109.2 ± 3.1 c | 243.0 ± 23.1 a | 371.1 ± 10.5 bc | 273.5 ± 7.3 ab | ||
| T+E+F | 83.5 ± 2.5 ab | 228.2 ± 7.7 de | 397.8 ± 11.7 c | 83.5 ± 4.6 ab | 272 ± 17.7 bc | 104.3 ± 3.5 a | 108.8 ± 5.1 c | 238.1 ± 13.3 a | 381.6 ± 13.1 c | 263.2 ± 5.9 ab | ||
| T+C+F | 75.4 ± 3.5 ab | 197.5 ± 8.4 cd | 342.7 ± 13.1 b | 75.4 ± 3.2 ab | 256 ± 17.7 bc | 94.0 ± 4.2 a | 98.7 ± 5.2 bc | 220.2 ± 21.1 a | 340.2 ± 12.4 bc | 244.3 ± 6.8 ab | ||
| T+C+E+F | 69.8 ± 5.1 a | 178.6 ± 5.8 bc | 798.3 ± 12.7 f | 69.8 ± 6.5 a | 163 ± 17.7 a | 52.1 ± 4.0 a | 50.9 ± 6.1 a | 231.5 ± 26.3 a | 308.5 ± 13.2 b | 230.4 ± 11.5 a | ||
| Significance | ||||||||||||
| A | 0.446 | 0.855 | 0.001 *** | 0.011 * | 0.001 ** | 0.001 *** | 0.001 *** | 0.181 | 0.001 *** | 0.497 | ||
| T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | ||
| A × T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.0287 * | 0.001 *** | 0.0287 * | ||
| Applications (A) X | Tyrosine | Cystine | Valine | Methionine | Tryptophan | Phenylalanine | Isoleucine | Leucine | Lysine | Hydroxy Proline | Sarcosine | Proline |
| Preventive | 268.0 ± 5.2 | 120.6 ± 3.2 b | 77.0 ± 2.3 b | 118.0 ± 3.4 b | 140.4 ± 7.2 b | 153.1 ± 3.3 b | 180 ± 311.3 b | 116.4 ± 2.3 ns | 186.4 ± 3.3 b | 105.2 ± 2.4 ns | 184.3 ± 4.1 ns | 39.0 ± 1.3 a |
| Curative | 272.1 ± 5.9 | 89.6 ± 3.5 a | 54.8 ± 2.6 a | 103.1 ± 3.3 a | 99.8 ± 7.8 a | 128.1 ± 3.9 a | 146.2 ± 11.7 a | 120.1 ± 2.6 | 162.1 ± 3.4 a | 101.2 ± 2.4 | 180.1 ± 4.3 | 28.4 ± 1.1 b |
| Treatments (T) Y | ||||||||||||
| C | 162.2 ± 9.6 a | 186.6 ± 7.6 b | 121.4 ± 5.6 b | 139.2 ± 7.2 b | 221.1 ± 11.6 b | 220.1 ± 3.3 b | 347.9 ± 21.8 b | 110.1 ± 5.1 ab | 204.0 ± 7.3 c | 147.6 ± 4.1 c | 141.8 ± 9.1 a | 76.2 ± 2.2 bc |
| F | 226.1 ± 2.3 b | 290.2 ± 7.6 c | 165.7 ± 2.2 c | 196.9 ± 6.2 c | 278.1 ± 13.3 b | 243.9 ± 3.1 b | 515.7 ± 21.1 c | 142.7 ± 4.6 c | 269.1 ± 3.3 d | 194.5 ± 5.3 d | 189.3 ± 6.2 bc | 86.6 ± 1.8 f |
| T+F | 393.3 ± 12.5 d | 210.5 ± 7.6 b | 144.4 ± 3.6 bc | 197.2 ± 7.1 c | 266.3 ± 12.1 b | 250.3 ± 8.1 b | 245.5 ± 23.8 b | 177.2 ± 5.5 d | 321.1 ± 5.1 e | 146.2 ± 6.5 c | 291.2 ± 6.3 d | 66.9 ± 2.8 b |
| C+F | 264.2 ± 11.6 b | 35.5 ± 7.6 a | 23.3 ± 3.1 a | 74.3 ± 5.2 a | 46.5 ± 14.6 a | 88.1 ± 7.3 a | 56.3 ± 22.4 a | 110.1 ± 4.2 a | 107.2 ± 5.6 a | 69.3 ± 3.1 ab | 152.0 ± 7.2 ab | 11.8 ± 3.5 a |
| E+F | 331.4 ± 9.1 c | 61.4 ± 7.6 a | 37.2 ± 5.6 a | 93.8 ± 5.6 a | 70.9 ± 14.5 a | 108.0 ± 6.1 a | 70.8 ± 22.2 a | 130.3 ± 6.6 bc | 187.0 ± 6.3 c | 90.0 ± 1.4 b | 226.8 ± 4.3 c | 15.3 ± 2.5 a |
| C+E+F | 280.3 ± 8.6 bc | 53.6 ± 7.6 a | 28.0 ± 3.6 a | 75.3 ± 4.2 a | 58.9 ± 11.0 a | 101.0 ± 7.3 a | 60.4 ± 21.3 a | 104.9 ± 4.3 ab | 142.3 ± 5.2 b | 52.8 ± 3.1 a | 181.1 ± 9.1 ab | 10.8 ± 2.2 a |
| T+E+F | 274.7 ± 11.2 bc | 37.9 ± 7.6 a | 25.6 ± 4.2 a | 80.3 ± 7.4 a | 49.4 ± 14.2 a | 89.1 ± 8.4 a | 58.7 ± 19.8 a | 106.0 ± 5.1 ab | 119.4 ± 7.3 ab | 78.7 ± 5.1 b | 164.3 ± 7.2 ab | 12.8 ± 2.1 a |
| T+C+F | 253.3 ± 12.6 b | 34.4 ± 7.6 a | 22.8 ± 7.1 a | 71.6 ± 5.5 a | 44.9 ± 6.6 a | 83.6 ± 6.5 a | 54.0 ± 17.5 a | 92.6 ± 5.4 a | 105.3 ± 2.3 a | 68.7 ± 4.1 ab | 147.3 ± 7.6 ab | 11.5 ± 2.0 a |
| T+C+E+F | 243.3 ± 12.2 b | 35.2 ± 7.6 a | 24.5 ± 3.6 a | 68.3 ± 6.2 a | 44.9 ± 12.5 a | 80.7 ± 7.3 a | 55.3 ± 24.4 a | 101.6 ± 4.6 a | 112.4 ± 4.4 ab | 78.9 ± 5.3 b | 148.2 ± 9.2 ab | 11.5 ± 1.7 a |
| Significance | ||||||||||||
| A | 0.628 | 0.001 *** | 0.001 *** | 0.003 ** | 0.001 *** | 0.001 *** | 0.047 * | 0.293 | 0.001 *** | 0.179 | 0.507 | 0.001 *** |
| T | 0.001 *** | 0.001 *** | 0.01 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| A × T | 0.001 *** | 0.001 *** | 0.01 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.003 ** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| Applications (A) X | Delphinidin-3-glycoside | Cyanidin-3-glycoside | Petunidin-3-glycoside | Peonidin-3-glycoside | Peonidin-3-glycoside_acetyl | Malvidin-3-glycoside_acetyl | Malvidin-3-glycoside | Malvidin-3-glycoside-p-coumaryl |
|---|---|---|---|---|---|---|---|---|
| Preventive | 4.8 ± 0.1 b | 2.6 ± 0.0 b | 9.3 ± 0.2 b | 45.6 ± 0.5 b | 83.5 ± 1.0 b | 55.6 ± 1.3 b | 9.2 ± 0.1 b | 9.4 ± 0.2 b |
| Curative | 2.5 ± 0.0 a | 1.6 ± 0.1 a | 5.4 ± 0.3 a | 19.7 ± 0.3 a | 42.7 ± 1.1 a | 31.5 ± 1.0 a | 4.9 ± 0.2 a | 4.9 ± 0.3 a |
| Treatments (T) Y | ||||||||
| C | 3.8 ± 0.1 c | 2.2 ± 0.0 b | 7.9 ± 0.2 cd | 37.9 ± 1.2 d | 65.7 ± 2.3 bc | 52.0 ± 2.6 de | 6.1 ± 0.2 bc | 5.2 ± 0.2 abc |
| F | 2.6 ± 0.2 b | 1.7 ± 0.4 a | 6.1 ± 0.3 b | 23.0 ± 1.1 bc | 45.3 ± 2.1 a | 40.2 ± 2.3 bc | 17.2 ± 0.1 e | 20.5 ± 0.6 e |
| T+F | 3.7 ± 0.0 c | 2.1 ± 0.0 b | 7.3 ± 0.2 c | 26.4 ± 1.1 c | 63.3 ± 2.2 b | 38.6 ± 2.2 bc | 6.2 ± 0.2 bc | 5.6 ± 0.5 bc |
| C+F | 5.1 ± 0.3 e | 2.6 ± 0.5 c | 9.0 ± 0.4 e | 46.8 ± 1.3 e | 88.2 ± 2.1 d | 47.8 ± 2.2 cd | 5.8 ± 0.0 b | 5.0 ± 0.6 abc |
| E+F | 3.5 ± 0.2 c | 2.1 ± 0.3 b | 7.4 ± 0.3 c | 38.6 ± 1.1 d | 60.8 ± 2.41 b | 47.8 ± 2.3 cd | 7.1 ± 0.1 c | 6.9 ± 0.6 c |
| C+E+F | 5.3 ± 0.0 e | 2.7 ± 0.0 c | 9.6 ± 0.1 e | 44.0 ± 1.4 e | 90.6 ± 2.0 d | 53.3 ± 2.4 de | 9.6 ± 0.2 d | 11.0 ± 0.4 d |
| T+E+F | 4.3 ± 0.1 d | 2.3 ± 0.1 bc | 8.8 ± 0.2 de | 42.5 ± 1.1 de | 73.9 ± 2.1 c | 59.8 ± 2.1 e | 5.7 ± 0.3 b | 4.4 ± 0.6 abc |
| T+C+F | 2.2 ± 0.3 a | 1.7 ± 0.0 a | 4.5 ± 0.3 a | 16.9 ± 1.2 a | 37.9 ± 2.0 a | 23.1 ± 2.2 a | 2.7 ± 0.2 a | 2.7 ± 0.3 a |
| T+C+E+F | 2.4 ± 0.1 ab | 1.6 ± 0.2 a | 5.4 ± 0.2 ab | 18.0 ± 1.1 ab | 42.2 ± 2.1 a | 29.6 ± 2.0 ab | 3.3 ± 0.1 a | 2.9 ± 0.6 ab |
| Significance | ||||||||
| A | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.293 |
| T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| A × T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| Applications (A) X | Oxalic | Propionic | Tartaric | Butyric | Malonic | Malic | Lactic | Citric | Maleic | Fumaric | Succinic |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Preventive | 6.3 ± 0.3 a | 12.5 ± 0.1 a | 20.5 ± 0.6 a | 59.9 ± 2.3 | 55.4 ± 1.2 a | 19.4 ± 0.3 a | 79.8 ± 2.9 a | 71.8 ± 1.4 a | 20.5 ± 0.9 a | 21.5 ± 1.1 | 94.2 ± 2.5 |
| Curative | 7.0 ± 0.1 b | 14.2 ± 0.4 b | 23.1 ± 0.3 b | 64.6 ± 2.1 | 61.6 ± 1.4 a | 22.8 ± 0.7 b | 89.9 ± 2.4 b | 83.0 ± 1.8 b | 24.1 ± 0.4 b | 24.3 ± 1.0 | 92.1 ± 2.6 |
| Treatments (T) Y | |||||||||||
| C | 3.9 ± 0.0 a | 5.3 ± 0.9 a | 2.8 ± 1.3 a | 4.3 ± 4.5 a | 5.1 ± 2.8 a | 2.9 ± 1.4 a | 2.7 ± 6.1 a | 7.5 ± 2.8 a | 2.1 ± 1.9 a | 4.6 ± 3.3 a | 5.4 ± 6.0 a |
| F | 5.8 ± 0.2 b | 8.3 ± 0.9 a | 3.4 ± 1.2 a | 5.9 ± 3.3 b | 6.8 ± 2.2 a | 3.1 ± 1.3 a | 2.9 ± 6.8 a | 9.9 ± 3.1 a | 2.6 ± 1.0 a | 4.7 ± 2.1 a | 8.8 ± 4.6 a |
| T+F | 8.7 ± 0.3 c | 17.4 ± 0.5 c | 33.6 ± 1.4 c | 94.15 ± 2.5 c | 80.7 ± 2.1 c | 26.3 ± 1.2 bc | 127.7 ± 3.2 c | 119.1 ± 4.2 c | 25.9 ± 1.1 b | 28.6 ± 1.1 b | 169.2 ± 3.6 d |
| C+F | 7.1 ± 0.4 bc | 15.3 ± 0.4 bc | 26.22 ± 1.4 b | 77.1 ± 4.3 bc | 74.3 ± 2.0 c | 28.3 ± 1.1 c | 108.3 ± 6.0 bc | 89.9 ± 3.6 b | 30.9 ± 1.9 b | 29.4 ± 2.1 b | 107.6 ± 5.6 bc |
| E+F | 7.4 ± 0.3 bc | 15.7 ± 0.6 bc | 27.2 ± 1.2 bc | 80.4 ± 4.4 bc | 78.9 ± 2.9 c | 26.4 ± 1.5 bc | 112.7 ± 6.4 bc | 102.0 ± 4.8 bc | 22.7 ± 1.4 b | 30.3 ± 2.4 b | 125.3 ± 4.6 c |
| C+E+F | 6.4 ± 0.3 b | 12.9 ± 0.3 b | 25.6 ± 1.3 b | 72.2 ± 4.5 b | 59.4 ± 2.2 b | 20.8 ± 1.3 b | 94.8 ± 3.2 b | 89.9 ± 1.4 b | 27.1 ± 1.1 b | 21.7 ± 2.1 b | 113.3 ± 5.6 bc |
| T+E+F | 7.4 ± 0.1 bc | 16.1 ± 0.9 bc | 27.1 ± 1.3 bc | 79.2 ± 4.0 bc | 78.4 ± 2.3 c | 29.7 ± 1.0 c | 114.1 ± 3.4 bc | 96.2 ± 2.8 b | 30.0 ± 1.2 b | 31.2 ± 2.5 b | 109.8 ± 5.1 bc |
| T+C+F | 6.7 ± 0.3 b | 14.6 ± 0.2 bc | 24.8 ± 1.7 b | 73.1 ± 4.1 bc | 71.6 ± 2.9 bc | 27.0 ± 1.2 bc | 103.4 ± 4.1 bc | 86.3 ± 3.3 b | 28.9 ± 1.9 b | 28.4 ± 2.1 b | 102.5 ± 5.6 bc |
| T+C+E+F | 6.6 ± 0.2 b | 14.4 ± 0.9 bc | 25.3 ± 1.4 b | 73.1 ± 4.5 bc | 71.2 ± 2.4 bc | 25.1 ± 1.5 bc | 96.5 ± 6.4 b | 95.3 ± 3.5 b | 30.1 ± 1.4 b | 26.7 ± 4.1 b | 96.0 ± 5.3 b |
| Significance | |||||||||||
| A | 0.010 * | 0.010 * | 0.010 * | 0.125 | 0.003 ** | 0.001 ** | 0.019 * | 0.001 *** | 0.008 ** | 0.060 | 0.583 |
| T | 0.001 *** | 0.001 *** | 0.001 | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| A × T | 0.001 *** | 0.002 ** | 0.004 ** | 0.050 | 40.001 *** | 0.002 ** | 0.001 ** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| Applications (A) X | Gallic-Acid | Vanillic-Acid | Trans-Caffeic-Acid | Trans-p-Coumaric-Acid | Ferulic-Acid | Caftaric-Acid | Catechin | Epicatechin | Quercetin | Routine | Myricetin | Tyrosol |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Preventive | 3.57 ± 0.1 b | 5.34 ± 0.3 b | 3.73 ± 0.1 b | 5.36 ± 0.1 b | 1.71 ± 0.2 | 6.73 ± 0.3 b | 7.10 ± 0.1 b | 4.23 ± 0.1 b | 2.57 ± 0.0 | 3.01 ± 0.1 b | 2.61 ± 0.2 b | 8.4 ± 0.3 b |
| Curative | 2.12 ± 0.4 a | 3.14 ± 0.2 a | 2.04 ± 0.4 a | 3.36 ± 0.2 a | 1.66 ± 0.1 | 3.77 ± 0.2 a | 4.39 ± 0.3 a | 2.25 ± 0.3 a | 2.65 ± 0.2 | 1.77 ± 0.2 a | 1.45 ± 0.1 a | 5.2 ± 0.2 a |
| Treatments (T) Y | ||||||||||||
| C | 2.50 ± 0.3 abc | 3.82 ± 0.3 abc | 2.70 ± 0.1 abc | 4.02 ± 0.3 abc | 1.17 ± 0.1 a | 4.57 ± 0.3 abc | 4.67 ± 0.3 ab | 3.15 ± 0.1 abc | 1.79 ± 0.1 a | 2.09 ± 0.2 | 1.59 ± 0.5 ab | 5.69 ± 0.7 abc |
| F | 2.48 ± 0.2 abc | 3.89 ± 0.4 abc | 2.57 ± 0.2 abc | 4.04 ± 0.4 abc | 1.75 ± 0.0 b | 4.82 ± 0.6 abc | 5.21 ± 0.7 ab | 2.86 ± 0.2 abc | 2.60 ± 0.3 b | 2.11 ± 0.6 | 1.69 ± 0.2 abc | 6.01 ± 0.3 abc |
| T+F | 3.33 ± 1.3 bc | 4.93 ± 0.4 bc | 3.28 ± 0.3 bc | 4.92 ± 0.4 bc | 2.26 ± 0.0 c | 6.30 ± 0.4 bc | 7.08 ± 0.6 ab | 3.54 ± 0.5 abc | 3.44 ± 0.1 c | 2.84 ± 0.7 | 2.64 ± 0.4 bc | 8.25 ± 0.4 bc |
| C+F | 3.52 ± 3.3 bc | 5.11 ± 0.1 bc | 3.58 ± 0.3 c | 5.07 ± 0.3 bc | 1.74 ± 0.1 b | 6.46 ± 0.5 bc | 6.98 ± 0.3 ab | 3.97 ± 0.3 bc | 2.70 ± 0.4 b | 3.01 ± 0.3 | 2.76 ± 0.6 bc | 8.42 ± 0.5 bc |
| E+F | 3.22 ± 1.2 abc | 4.74 ± 0.1 bc | 3.17 ± 0.2 bc | 5.04 ± 0.2 bc | 1.54 ± 0.2 ab | 5.80 ± 0.6 abc | 6.48 ± 0.1 ab | 3.72 ± 0.4 bc | 2.38 ± 0.5 ab | 2.58 ± 0.2 | 2.04 ± 0.3 abc | 7.47 ± 0.7 abc |
| C+E+F | 3.89 ± 0.4 c | 5.71 ± 0.2 c | 4.05 ± 0.3 c | 5.66 ± 0.1 c | 1.60 ± 0.1 ab | 7.25 ± 0.1 c | 7.65 ± 0.2 b | 4.57 ± 0.2 c | 2.43 ± 0.3 b | 3.30 ± 0.1 | 2.95 ± 0.2 c | 9.12 ± 0.6 c |
| T+E+F | 2.75 ± 0.2 abc | 4.23 ± 0.3 abc | 2.96 ± 0.2 abc | 4.39 ± 0.4 abc | 1.56 ± 0.3 ab | 5.19 ± 0.0 abc | 5.38 ± 0.7 ab | 3.42 ± 0.4 abc | 2.40 ± 0.1 ab | 2.27 ± 0.2 | 1.81 ± 0.1 abc | 6.39 ± 0.2 abc |
| T+C+F | 1.75 ± 0.5 a | 2.56 ± 0.4 a | 1.65 ± 0.1 a | 2.69 ± 0.3 a | 1.71 ± 0.1 b | 3.10 ± 0.3 a | 3.68 ± 0.5 a | 1.74 ± 0.5 a | 2.75 ± 0.6 b | 1.50 ± 0.3 | 1.32 ± 0.2 a | 4.44 ± 0.7 a |
| T+C+E+F | 2.17 ± 0.3 ab | 3.16 ± 0.3 ab | 2.02 ± 0.3 ab | 3.42 ± 0.2 ab | 1.86 ± 0.1 bc | 3.78 ± 0.6 ab | 4.56 ± 0.7 ab | 2.19 ± 0.3 ab | 3.00 ± 0.1 bc | 1.80 ± 0.5 | 1.51 ± 0.4 ab | 5.40 ± 0.1 ab |
| Significance | ||||||||||||
| A | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.509 | 0.001 *** | 0.001 *** | 0.001 *** | 0.430 | 0.001 *** | 0.001 *** | 0.001 *** |
| T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.005 ** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| A × T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.898 | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| Applications (A) X | IAA | ABA | GA | SA | Cytokinin | Zeatin | Jasmonic_Acid |
|---|---|---|---|---|---|---|---|
| Preventive | 2.9 ± 0.1 b | 196.0 ± 9.4 a | 4.6 ± 0.1 a | 2.1 ± 0.3 b | 3.1 ± 0.0 b | 1.8 ± 0.1 a | 0.8 ± 0.0 a |
| Curative | 2.2 ± 0.0 a | 230.2 ± 10.4 b | 12.3 ± 0.2 b | 1.8 ± 0.0 a | 2.0 ± 0.2 a | 2.1 ± 0.0 b | 1.0 ± 0.3 b |
| Treatments (T) Y | |||||||
| C | 3.6 ± 0.1 c | 110.0 ± 21.2 a | 2.9 ± 0.1 a | 6.0 ± 0.1 d | 7.2 ± 0.4 c | 1.9 ± 0.1 b | 0.9 ± 0.0 b |
| F | 5.3 ± 0.2 d | 232.3 ± 11.4 b | 4.5 ± 0.2 b | 4.0 ± 0.6 e | 9.7 ± 0.1 d | 2.5 ± 0.3 c | 1.7 ± 0.1 c |
| T+F | 3.9 ± 0.1 c | 145.4 ± 15.8 ab | 9.8 ± 0.3 c | 3.3 ± 0.4 c | 5.4 ± 0.5 b | 1.6 ± 0.0 a | 1.0 ± 0.0 b |
| C+F | 1.7 ± 0.1 ab | 362.4 ± 21.2 c | 8.8 ± 0.6 c | 1.1 ± 0.0 b | 0.2 ± 0.1 a | 2.3 ± 0.1 c | 0.9 ± 0.0 b |
| E+F | 1.8 ± 0.2 ab | 483.0 ± 20.9 d | 2.8 ± 0.3 a | 1.1 ± 0.3 b | 0.2 ± 0.5 a | 2.6 ± 0.0 c | 0.9 ± 0.2 b |
| C+E+F | 1.8 ± 0.1 ab | 141.2 ± 21.6 ab | 4.0 ± 0.3 ab | 0.6 ± 0.0 a | 0.1 ± 0.2 a | 1.9 ± 0.2 b | 0.6 ± 0.0 a |
| T+E+F | 1.6 ± 0.3 ab | 168.3 ± 22.8 ab | 12.1 ± 0.4 d | 0.6 ± 0.1 a | 0.1 ± 0.1 a | 1.6 ± 0.0 a | 0.7 ± 0.3 a |
| T+C+F | 1.3 ± 0.1 a | 118.4 ± 21.4 a | 22.8 ± 0.3 e | 0.6 ± 0.0 a | 0.1 ± 0.7 a | 1.6 ± 0.4 a | 0.6 ± 0.0 a |
| T+C+E+F | 2.1 ± 0.4 b | 161.5 ± 23.1 ab | 8.7 ± 0.1 c | 0.5 ± 0.3 a | 0.1 ± 0.1 a | 1.5 ± 0.0 a | 0.6 ± 0.1 a |
| Significance | |||||||
| A | 0.001 *** | 0.026 * | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
| A × T | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** | 0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karakus, S. Enhancing Post-Harvest Resilience: Investigating the Synergistic Effects of Essential Oil Combinations on Biochemical Profiles in Botrytis cinerea-Infected Apples. Horticulturae 2024, 10, 341. https://doi.org/10.3390/horticulturae10040341
Karakus S. Enhancing Post-Harvest Resilience: Investigating the Synergistic Effects of Essential Oil Combinations on Biochemical Profiles in Botrytis cinerea-Infected Apples. Horticulturae. 2024; 10(4):341. https://doi.org/10.3390/horticulturae10040341
Chicago/Turabian StyleKarakus, Sinem. 2024. "Enhancing Post-Harvest Resilience: Investigating the Synergistic Effects of Essential Oil Combinations on Biochemical Profiles in Botrytis cinerea-Infected Apples" Horticulturae 10, no. 4: 341. https://doi.org/10.3390/horticulturae10040341
APA StyleKarakus, S. (2024). Enhancing Post-Harvest Resilience: Investigating the Synergistic Effects of Essential Oil Combinations on Biochemical Profiles in Botrytis cinerea-Infected Apples. Horticulturae, 10(4), 341. https://doi.org/10.3390/horticulturae10040341