
Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of the Hsp70 Family Genes in Cucumber
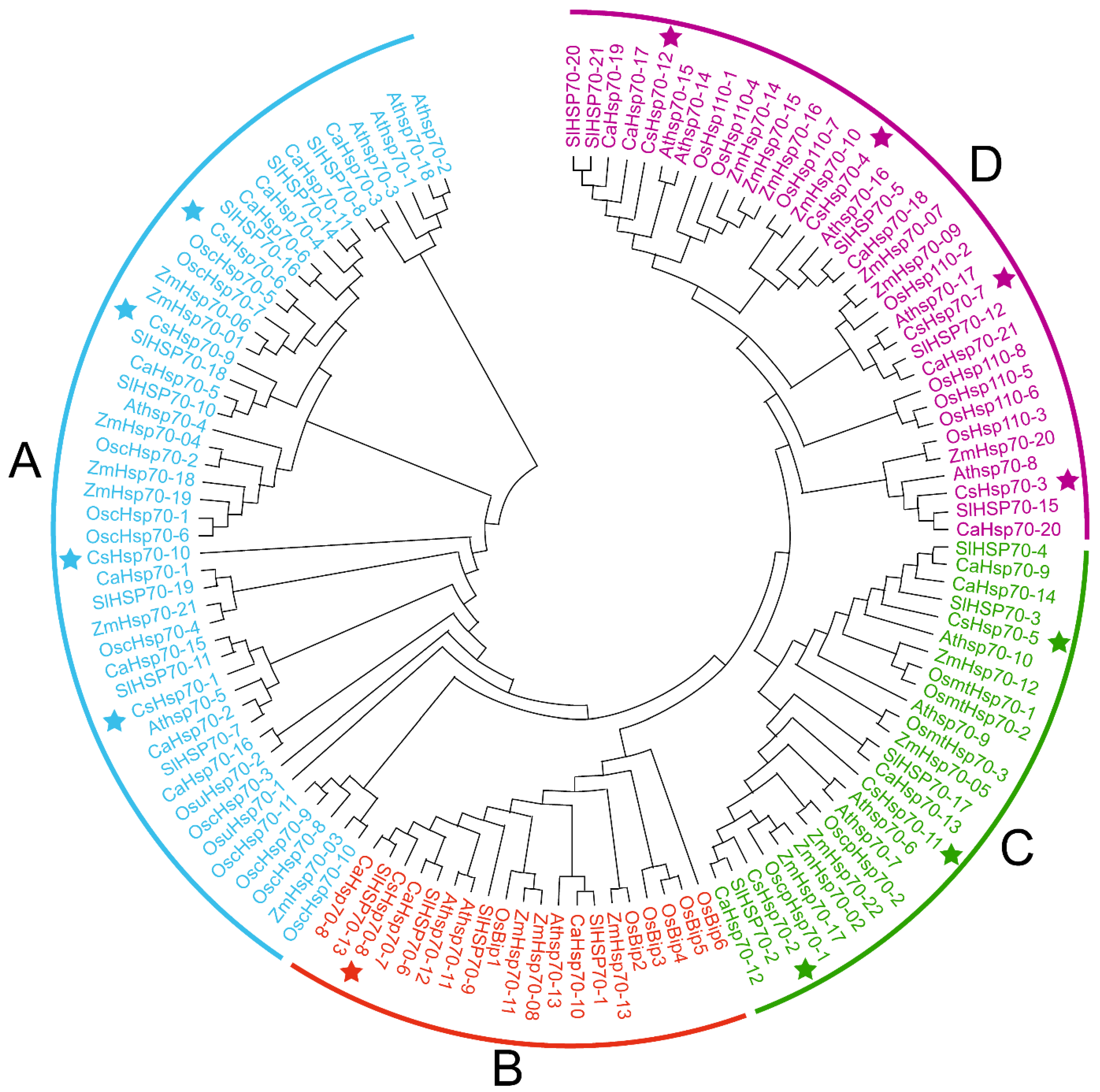
2.2. Evolutionary Relationships of CsHsp70 Proteins
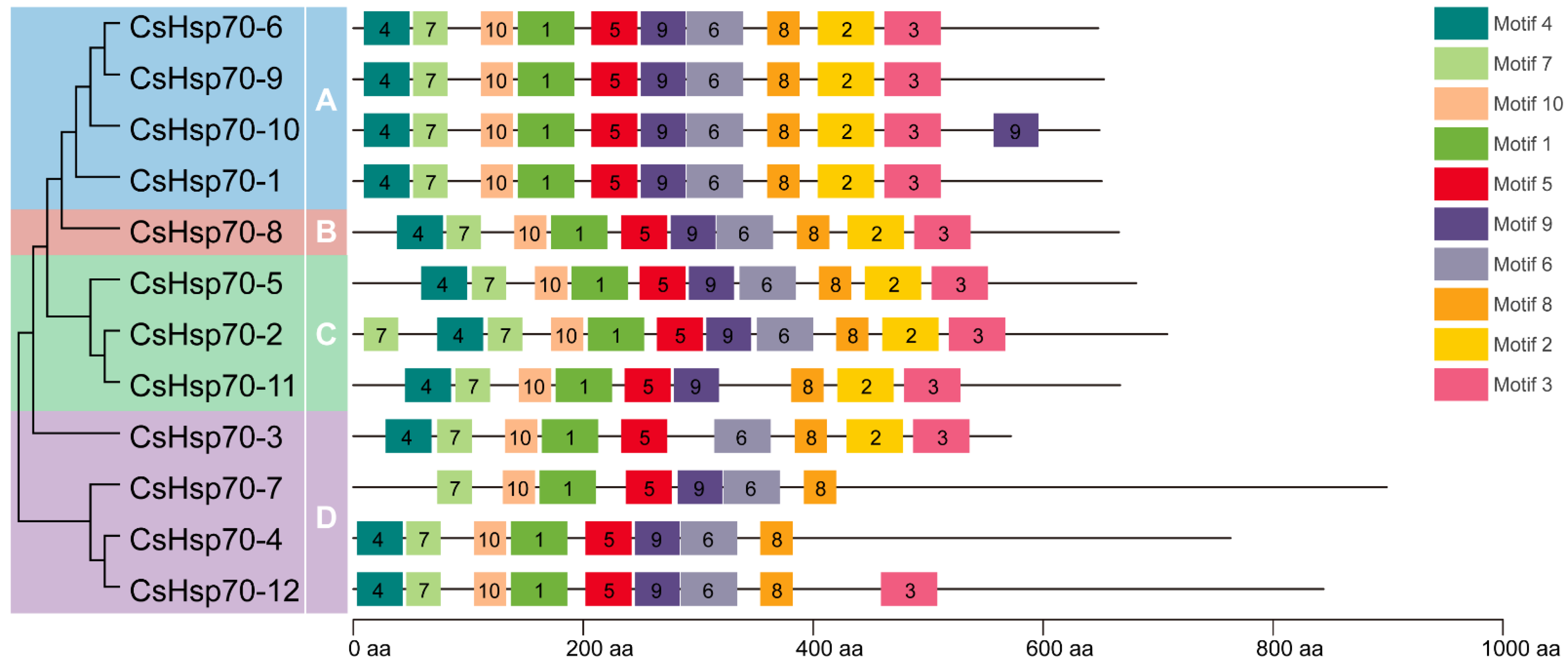
2.3. Conserved Motif Analysis of CsHsp70 Proteins
2.4. Chromosomal Location and Gene Duplication Analysis of the CsHsp70 Genes
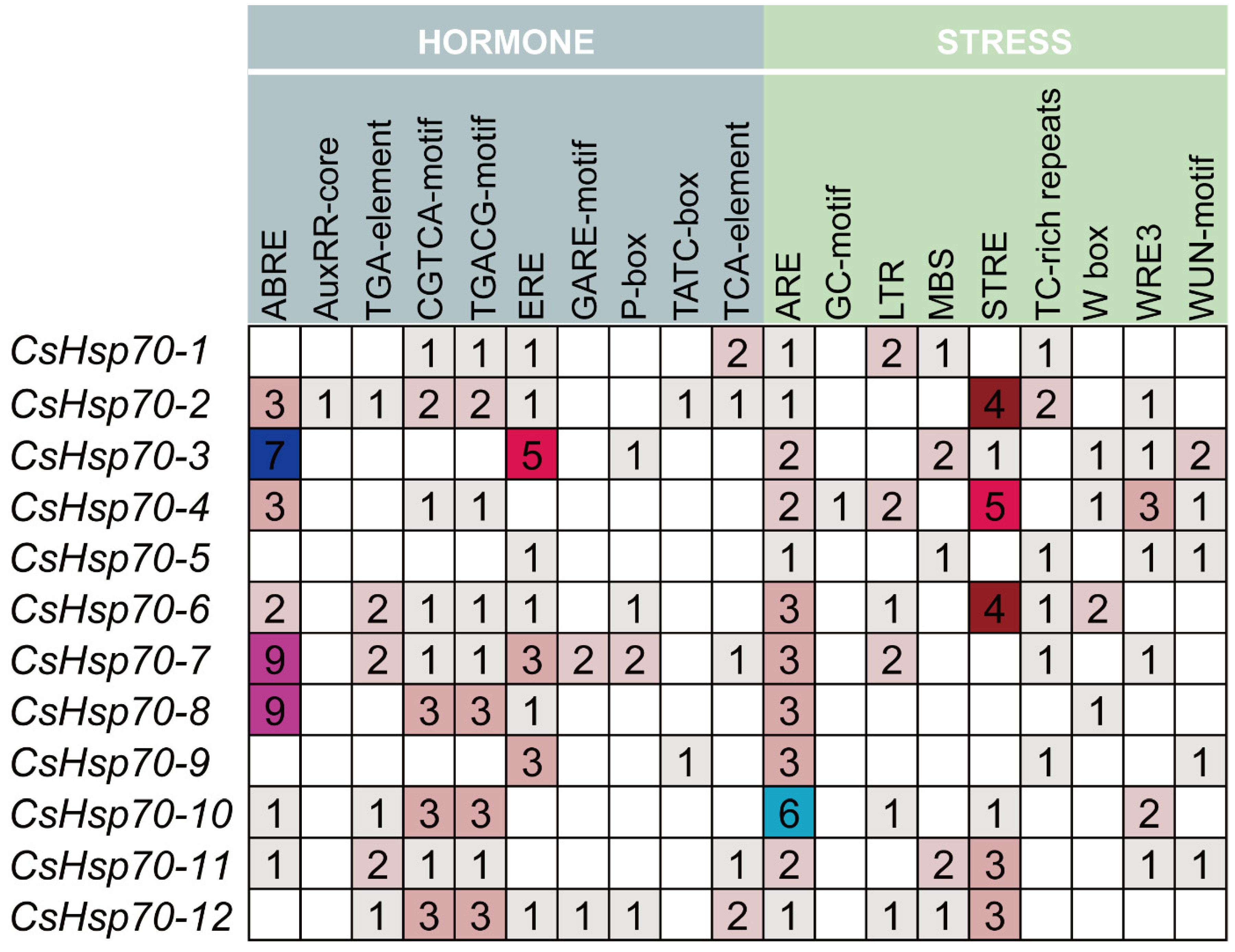
2.5. Cis-Acting Regulatory Element Analysis of the CsHsp70 Genes
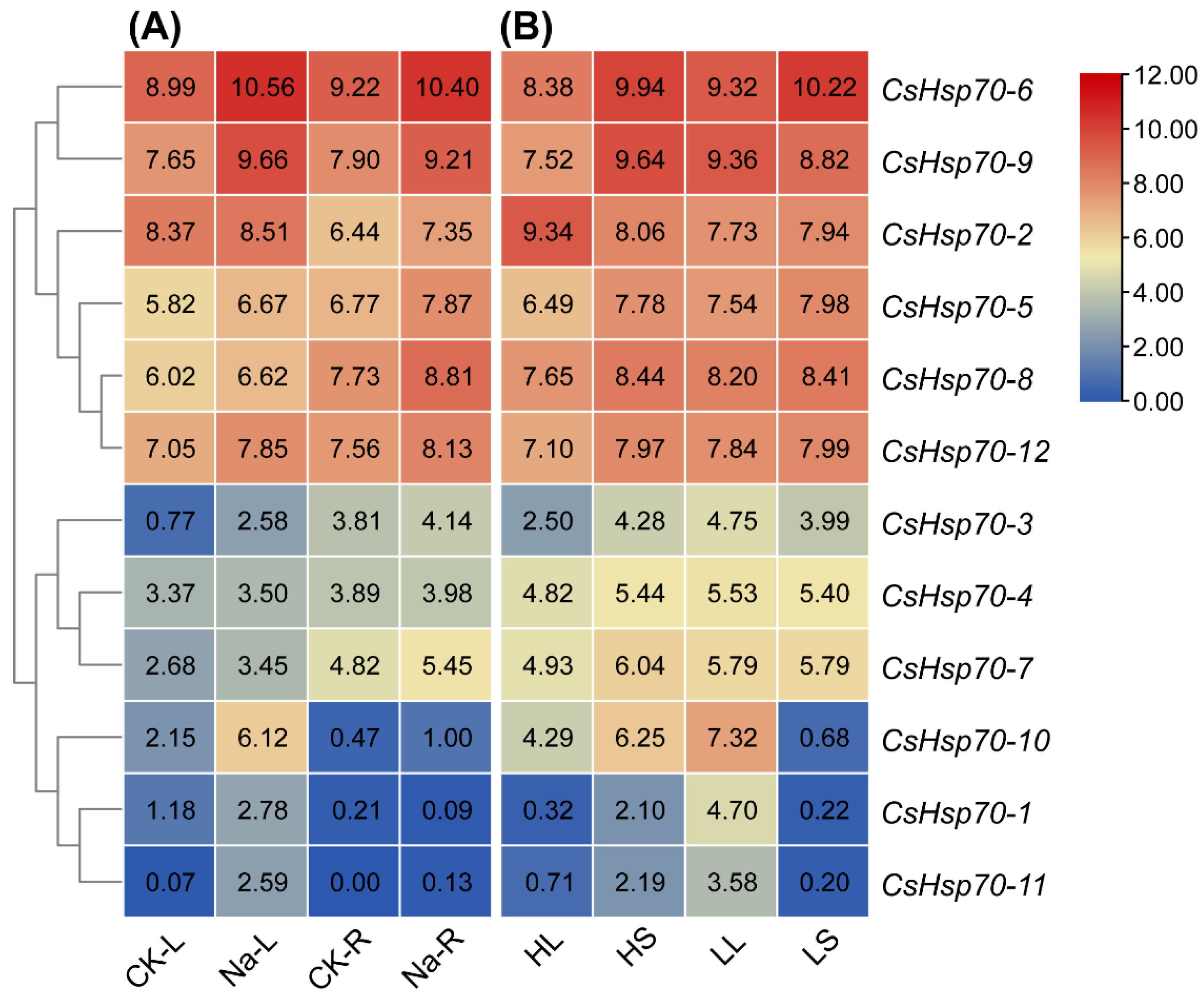
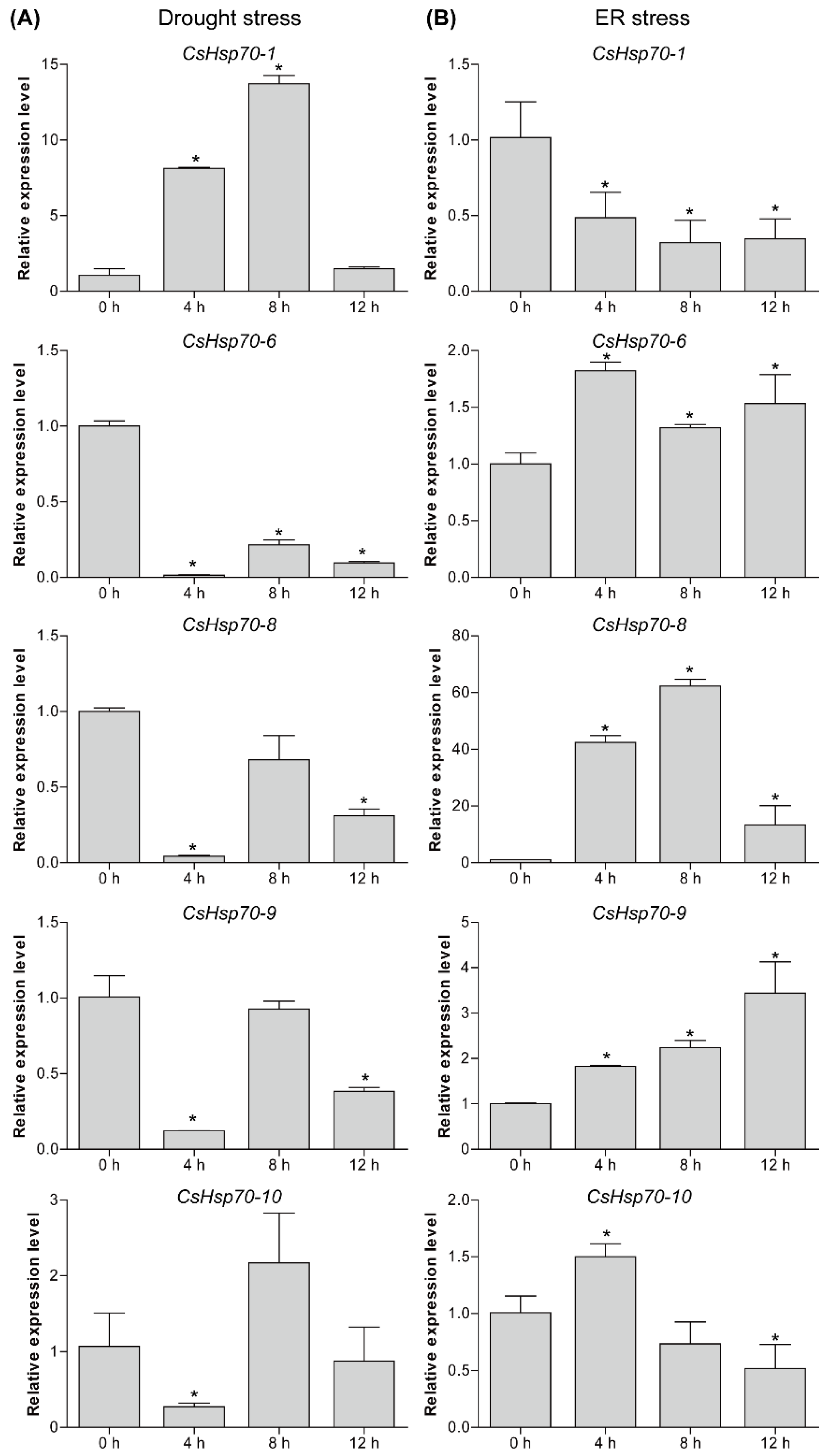
2.6. Expression Analysis of CsHsp70 Genes under Different Abiotic Stresses
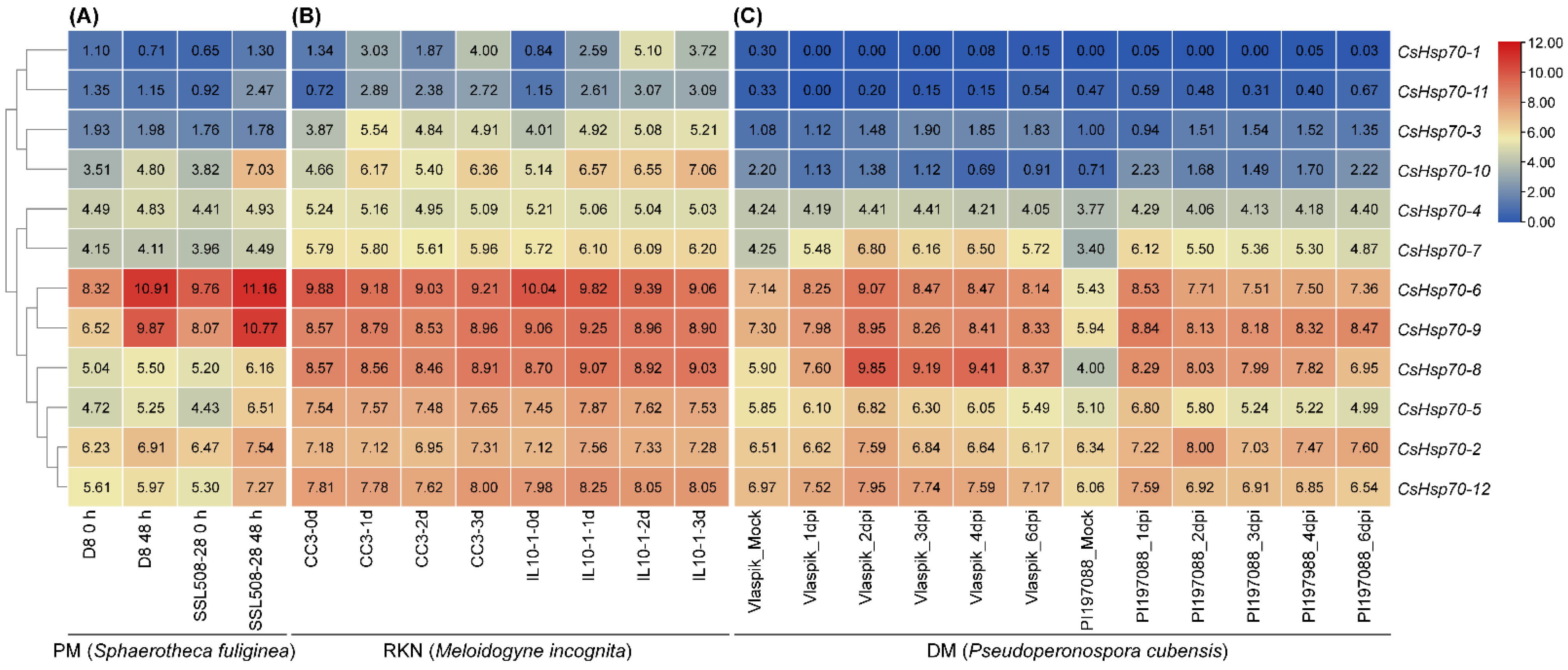
2.7. Expression Analysis of CsHsp70 Genes in Response to Different Biotic Stresses
3. Discussion
4. Materials and Methods
4.1. Database Searches and Annotation of the Hsp70 Members in Cucumber
4.2. Phylogenetic Analysis and Conserved Motif Identification
4.3. Prediction of Chromosomal Locations, Gene Duplication, and Cis-Acting Regulatory Elements of CsHsp70 Genes
4.4. Expression Analysis of the CsHsp70 Genes via RNA-Seq Data
4.5. Plant Materials and Stress Treatments
4.6. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR) Analysis of the CsHsp70 Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yu, A.; Li, P.; Tang, T.; Wang, J.; Chen, Y.; Liu, L. Roles of Hsp70s in stress responses of microorganisms, plants, and animals. Biomed. Res. Int. 2015, 2015, 510319. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- De Maio, A.; Hightower, L.E. Heat shock proteins and the biogenesis of cellular membranes. Cell Stress Chaperones 2021, 26, 15–18. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular analysis of Hsp70 mechanisms in plants and their function in response to stress. Biotechnol. Genet. Eng. Rev. 2017, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef]
- Cho, E.K.; Choi, Y.J. A nuclear-localized HSP70 confers thermoprotective activity and drought-stress tolerance on plants. Biotechnol. Lett. 2009, 31, 597–606. [Google Scholar] [CrossRef]
- Jung, K.-H.; Gho, H.-J.; Nguyen, M.X.; Kim, S.-R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef]
- Sarkar, N.K.; Kundnani, P.; Grover, A. Functional analysis of Hsp70 superfamily proteins of rice (Oryza sativa). Cell Stress Chaperones 2013, 18, 427–437. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Zhai, Y.F.; Gong, Z.H.; Lu, M.H. Genome-wide analysis of the Hsp70 family genes in pepper (Capsicum annuum L.) and functional identification of CaHsp70-2 involvement in heat stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef]
- Liu, J.; Pang, X.; Cheng, Y.; Yin, Y.; Zhang, Q.; Su, W.; Hu, B.; Guo, Q.; Ha, S.; Zhang, J.; et al. The Hsp70 gene family in Solanum tuberosum: Genome-wide identification, phylogeny, and expression patterns. Sci. Rep. 2018, 8, 16628. [Google Scholar] [CrossRef]
- Song, Z.; Pan, F.; Lou, X.; Wang, D.; Yang, C.; Zhang, B.; Zhang, H. Genome-wide identification and characterization of Hsp70 gene family in Nicotiana tabacum. Mol. Biol. Rep. 2019, 46, 1941–1954. [Google Scholar] [CrossRef]
- Su, H.; Xing, M.; Liu, X.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Lv, H. Genome-wide analysis of HSP70 family genes in cabbage (Brassica oleracea var. capitata) reveals their involvement in floral development. BMC Genom. 2019, 20, 369. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Shah, Z.; Ullah, A.; Ahmed, S.; Ahmad, B.; Khan, A. Genome wide and evolutionary analysis of heat shock protein 70 proteins in tomato and their role in response to heat and drought stress. Mol. Biol. Rep. 2022, 49, 11229–11241. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Li, Y.; Elbaiomy, R.G.; Yan, K.; Ragauskas, A.J.; Yadav, V.; Soaud, S.A.; Islam, M.M.; Saleem, N.; Noor, Z.; et al. Genome-wide analysis and expression profiling of SlHsp70 gene family in Solanum lycopersicum revealed higher expression of SlHsp70-11 in roots under Cd2+ stress. Front. Biosci. 2022, 27, 186. [Google Scholar] [CrossRef] [PubMed]
- Davoudi, M.; Chen, J.; Lou, Q. Genome-wide identification and expression analysis of heat shock protein 70 (HSP70) gene family in pumpkin (Cucurbita moschata) rootstock under drought stress suggested the potential role of these chaperones in stress tolerance. Int. J. Mol. Sci. 2022, 23, 1918. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Hu, R.; Song, A.; Guan, Z.; Chen, F.; Jiang, J. Genome-wide identification and expression analysis of HSP70 gene family in Chrysanthemum lavandulifolium under heat stress. Horticulturae 2023, 9, 238. [Google Scholar] [CrossRef]
- He, Q.; Zhang, X.; He, M.; Zhang, X.; Ma, Y.; Zhu, Y.; Dong, J.; Ying, J.; Wang, Y.; Liu, L.; et al. Genome-wide characterization of RsHSP70 gene family reveals positive role of RsHSP70-20 gene in heat stress response in radish (Raphanus sativus L.). Plant Physiol. Biochem. 2023, 199, 107710. [Google Scholar] [CrossRef]
- Basile, A.; Sorbo, S.; Cardi, M.; Lentini, M.; Castiglia, D.; Cianciullo, P.; Conte, B.; Loppi, S.; Esposito, S. Effects of heavy metals on ultrastructure and Hsp70 induction in Lemna minor L. exposed to water along the Sarno River, Italy. Ecotoxicol. Environ. Saf. 2015, 114, 93–101. [Google Scholar] [CrossRef]
- Büyük, İ.; Inal, B.; Ilhan, E.; Tanriseven, M.; Aras, S.; Erayman, M. Genome-wide identification of salinity responsive HSP70s in common bean. Mol. Biol. Rep. 2016, 43, 1251–1266. [Google Scholar] [CrossRef]
- Liu, J.; Wang, R.; Liu, W.; Zhang, H.; Guo, Y.; Wen, R. Genome-wide characterization of heat-shock protein 70s from Chenopodium quinoa and expression analyses of Cqhsp70s in response to drought stress. Genes 2018, 9, 35. [Google Scholar] [CrossRef]
- Coutinho, F.S.; dos Santos, D.S.; Lima, L.L.; Vital, C.E.; Santos, L.A.; Pimenta, M.R.; da Silva, J.C.; Ramos, J.R.L.S.; Mehta, A.; Fontes, E.P.B.; et al. Mechanism of the drought tolerance of a transgenic soybean overexpressing the molecular chaperone BiP. Physiol. Mol. Biol. Plants 2019, 25, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Samraj, S.; Jiménez, J.; Gómez, C.; Liu, T.; Begcy, K. Genome-wide identification of heat shock factors and heat shock proteins in response to UV and high intensity light stress in lettuce. BMC Plant Biol. 2021, 21, 185. [Google Scholar] [CrossRef] [PubMed]
- Jungkunz, I.; Link, K.; Vogel, F.; Voll, L.M.; Sonnewald, S.; Sonnewald, U. AtHsp70-15-deficient Arabidopsis plants are characterized by reduced growth, a constitutive cytosolic protein response and enhanced resistance to TuMV. Plant J. 2011, 66, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, F.; Thomas, C.L.; Lederer, C.; Niu, Y.; Wang, D.; Maule, A.J. Virus induction of heat shock protein 70 reflects a general response to protein accumulation in the plant cytosol. Plant Physiol. 2005, 138, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Liu, N.; Yu, Y.; Bi, C.; Chen, Q.; Qu, Y. The cotton 70-kDa heat shock protein GhHSP70-26 plays a positive role in the drought stress response. Environ. Exp. Bot. 2021, 191, 104628. [Google Scholar] [CrossRef]
- Park, C.J.; Park, J.M. Endoplasmic reticulum plays a critical role in integrating signals generated by both biotic and abiotic stress in plants. Front. Plant Sci. 2019, 10, 399. [Google Scholar] [CrossRef]
- Wang, H.; Niu, H.; Zhai, Y.; Lu, M. Characterization of BiP genes from pepper (Capsicum annuum L.) and the role of CaBiP1 in response to endoplasmic reticulum and multiple abiotic stresses. Front. Plant Sci. 2017, 8, 1122. [Google Scholar] [CrossRef]
- Zhu, X.; Teng, X.; Wang, Y.; Hao, Y.; Jing, R.; Wang, Y.; Liu, Y.; Zhu, J.; Wu, M.; Zhong, M.; et al. FLOURY ENDOSPERM11 encoding a plastid heat shock protein 70 is essential for amyloplast development in rice. Plant Sci. 2018, 277, 89–99. [Google Scholar] [CrossRef]
- Zhu, C.; Xiao, L.; Hu, Y.; Liu, L.; Liu, H.; Hu, Z.; Liu, S.; Zhou, Y. Genome-wide survey and expression analysis of B-Box family genes in cucumber reveal their potential roles in response to diverse abiotic and biotic stresses. Agriculture 2022, 12, 827. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Tang, R.; Wang, L.; Chen, C.; Ren, Z. Genome-wide identification and expression analysis of Hsf and Hsp gene families in cucumber (Cucumis sativus L.). Plant Growth Regul. 2021, 95, 223–239. [Google Scholar] [CrossRef]
- Lin, B.L.; Wang, J.S.; Liu, H.C.; Chen, R.W.; Meyer, Y.; Barakat, A.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lai, Y.; Zhang, W.; Ahmad, J.; Qiu, Y.; Zhang, X.; Duan, M.; Liu, T.; Song, J.; Wang, H.; et al. MicroRNAs and their targets in cucumber shoot apices in response to temperature and photoperiod. Can. J. Plant Sci. 2018, 19, 819. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, J.; Liang, Y.; Liu, J.; Jia, J.; Huo, H.; Wu, Z.; Yang, R.; Gong, H. Transcriptomic dynamics provide an insight into the mechanism for silicon-mediated alleviation of salt stress in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 174, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Yang, L.; Liu, N.; Yang, J.; Zhou, X.K.; Xia, Y.C.; He, Y.; He, Y.Q.; Gong, H.J.; Ma, D.F.; et al. Genome-wide identification, structure characterization, and expression pattern profiling of aquaporin gene family in cucumber. BMC Plant Biol. 2019, 19, 345. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, A.; Day, B. Transcriptome and small RNAome dynamics during a resistant and susceptible interaction between cucumber and downy mildew. Plant Genome 2016, 9, 1–19. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Shi, Y.; Qi, X.; Chen, X. Elucidation of the molecular responses of a cucumber segment substitution line carrying Pm5.1 and its recurrent parent triggered by powdery mildew by comparative transcriptome profiling. BMC Genom. 2017, 18, 21. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, C.; Zhang, K.; Tian, Z.; Xu, J.; Yang, S.; Lou, Q.; Li, J.; Chen, J.F. Comparative transcriptomics reveals suppressed expression of genes related to auxin and the cell cycle contributes to the resistance of cucumber against Meloidogyne incognita. BMC Genom. 2018, 19, 583. [Google Scholar] [CrossRef]
- Augustine, S.M.; Narayan, J.A.; Syamaladevi, D.P.; Appunu, C.; Chakravarthi, M.; Ravichandran, V.; Subramonian, N. Erianthus arundinaceus HSP70 (EaHSP70) overexpression increases drought and salinity tolerance in sugarcane (Saccharum spp. hybrid). Plant Sci. 2015, 232, 23–34. [Google Scholar] [CrossRef]
- Rathor, P.; Borza, T.; Bahmani, R.; Stone, S.; Tonon, T.; Yurgel, S.; Potin, P.; Prithiviraj, B. Expression of a Heat Shock Protein 70 from the brown alga Ectocarpus sp. imparts salinity stress tolerance in Arabidopsis thaliana. J. Appl. Phycol. 2023, 35, 803–819. [Google Scholar] [CrossRef]
- Wakasa, Y.; Hayashi, S.; Takaiwa, F. Expression of OsBiP4 and OsBiP5 is highly correlated with the endoplasmic reticulum stress response in rice. Planta 2012, 236, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Berka, M.; Kopecká, R.; Berková, V.; Brzobohatý, B.; Černý, M. Regulation of heat shock proteins 70 and their role in plant immunity. J. Exp. Bot. 2022, 73, 1894–1909. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Bart, R.; Chern, M.; Canlas, P.E.; Bai, W.; Ronald, P.C. Overexpression of the endoplasmic reticulum chaperone BiP3 regulates XA21-mediated innate immunity in rice. PLoS ONE 2010, 5, e9262. [Google Scholar] [CrossRef]
- Yang, S.; Zhu, C.; Chen, J.; Zhao, J.; Hu, Z.; Liu, S.; Zhou, Y. Identification and expression profile analysis of the OSCA gene family related to abiotic and biotic stress response in cucumber. Biology 2022, 11, 1134. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. (v2) | Accession No. (v3) | Chromosome: Location | CDS/bp | Physicochemical Parameters of Proteins | |||
|---|---|---|---|---|---|---|---|---|
| AA | pI | MW/kDa | GRAVY | |||||
| CsHsp70-1 | Csa2G070310.1 | CsaV3_2G009170 | Chr2: 5,427,208 .. 5,429,443 (+) | 1953 | 650 | 5.18 | 71.27995 | −0.422 |
| CsHsp70-2 | Csa2G122520.1 | CsaV3_2G011210 | Chr2: 8,176,614 .. 8,180,834 (+) | 2124 | 707 | 5.18 | 75.39607 | −0.295 |
| CsHsp70-3 | Csa3G147740.1 | CsaV3_3G013380 | Chr3: 9,928,649 .. 9,930,364 (−) | 1716 | 571 | 5.52 | 61.98548 | 0.040 |
| CsHsp70-4 | Csa3G391900.1 | CsaV3_3G022450 | Chr3: 19,117,817 .. 19,123,998 (−) | 2289 | 762 | 5.63 | 85.39136 | −0.422 |
| CsHsp70-5 | Csa4G179170.1 | CsaV3_4G014660 | Chr4: 8,965,693 .. 8,969,625 (−) | 2043 | 680 | 5.69 | 73.1238 | −0.323 |
| CsHsp70-6 | Csa4G295440.1 | CsaV3_4G024840 | Chr4: 11,823,184 .. 11,826,273 (+) | 1944 | 647 | 5.16 | 70.86926 | −0.403 |
| CsHsp70-7 | Csa4G617390.1 | CsaV3_4G032260 | Chr4: 19,669,174 .. 19,679,176 (+) | 2697 | 898 | 5.27 | 100.13606 | −0.498 |
| CsHsp70-8 | Csa5G149330.1 | CsaV3_5G001960 | Chr5: 4,469,887 .. 4,473,757 (+) | 1998 | 665 | 5.10 | 73.42913 | −0.474 |
| CsHsp70-9 | Csa5G512930.1 | CsaV3_5G026450 | Chr5: 17,912,303 .. 17,915,006 (+) | 1959 | 652 | 5.10 | 71.45378 | −0.442 |
| CsHsp70-10 | Csa5G514500.1 | CsaV3_5G026520 | Chr5: 17,969,849 .. 17,972,430 (−) | 1947 | 648 | 5.21 | 71.04657 | −0.415 |
| CsHsp70-11 | Csa7G312930.1 | CsaV3_7G024760 | Chr7: 10,868,096 .. 10,872,494 (+) | 2001 | 666 | 5.09 | 71.51079 | −0.250 |
| CsHsp70-12 | Csa7G446710.1 | CsaV3_7G033150 | Chr7: 17,685,920 .. 17,691,540 (−) | 2532 | 843 | 5.40 | 92.89913 | −0.430 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Xiao, L.; Zhao, J.; Hu, Z.; Zhou, Y.; Liu, S.; Wu, H.; Zhou, Y. Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber. Horticulturae 2023, 9, 1057. https://doi.org/10.3390/horticulturae9091057
Zhou Z, Xiao L, Zhao J, Hu Z, Zhou Y, Liu S, Wu H, Zhou Y. Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber. Horticulturae. 2023; 9(9):1057. https://doi.org/10.3390/horticulturae9091057
Chicago/Turabian StyleZhou, Zixian, Lingdi Xiao, Jindong Zhao, Zhaoyang Hu, Yuelong Zhou, Shiqiang Liu, Hao Wu, and Yong Zhou. 2023. "Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber" Horticulturae 9, no. 9: 1057. https://doi.org/10.3390/horticulturae9091057
APA StyleZhou, Z., Xiao, L., Zhao, J., Hu, Z., Zhou, Y., Liu, S., Wu, H., & Zhou, Y. (2023). Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber. Horticulturae, 9(9), 1057. https://doi.org/10.3390/horticulturae9091057