
Somatic Embryogenesis and Plant Regeneration from Stem Explants of Pomegranate

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Tissue Culture Plantlets
2.2. Screening Optimum Combination of Media and Plant Growth Regulators for Callus Induction
2.3. Screening Optimum Concentrations of Plant Growth Regulators for Embryogenic Callus Induction
2.4. Evaluation of Different Explants on Embryogenic Callus Induction
2.5. Evaluation of Different Subculture Cycles on Proliferation of Embryogenic Calli
2.6. Screening Optimum Concentrations of Plant Growth Regulators for Somatic Embryogenesis
2.7. Rooting and Regeneration of Somatic Embryo Seedlings
3. Results
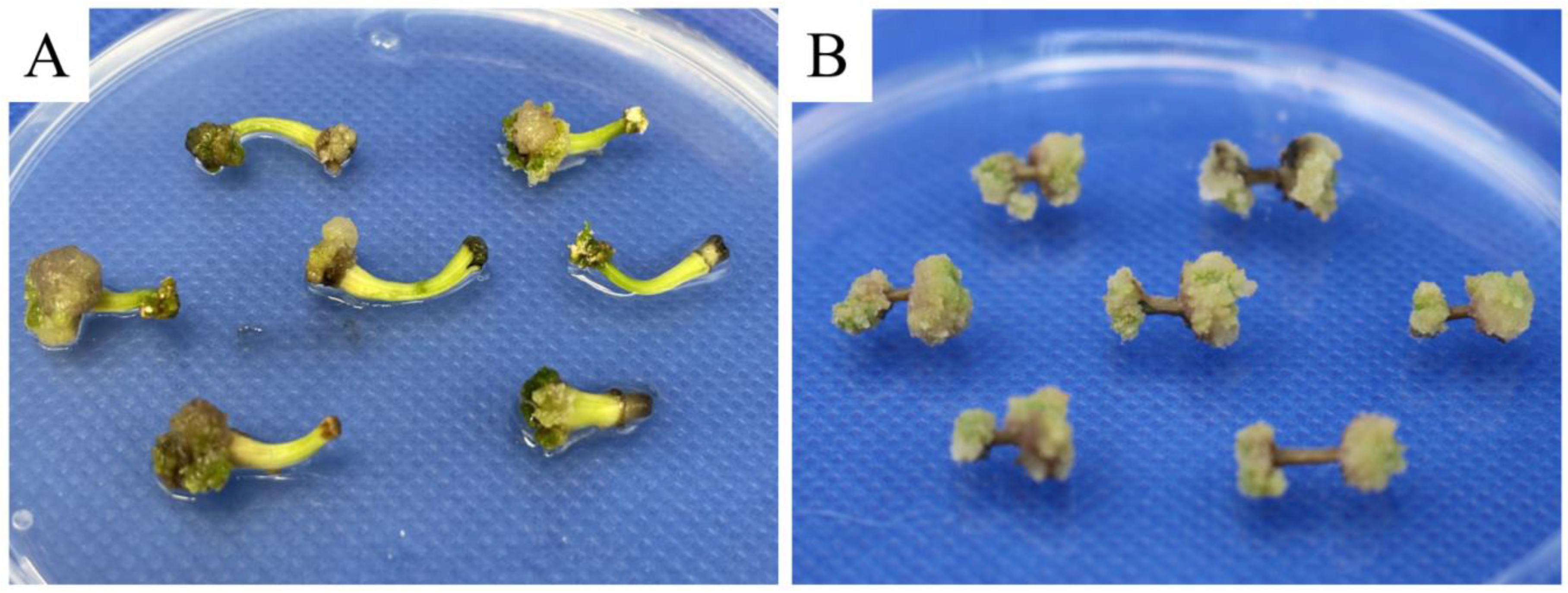
3.1. Effects of Different Combinations of Media and Plant Growth Regulators on Callus Induction
3.2. Effects of Different Concentrations of Plant Growth Regulators on Embryogenic Callus Induction
3.3. Effect of Different Explants on Embryogenic Callus Induction
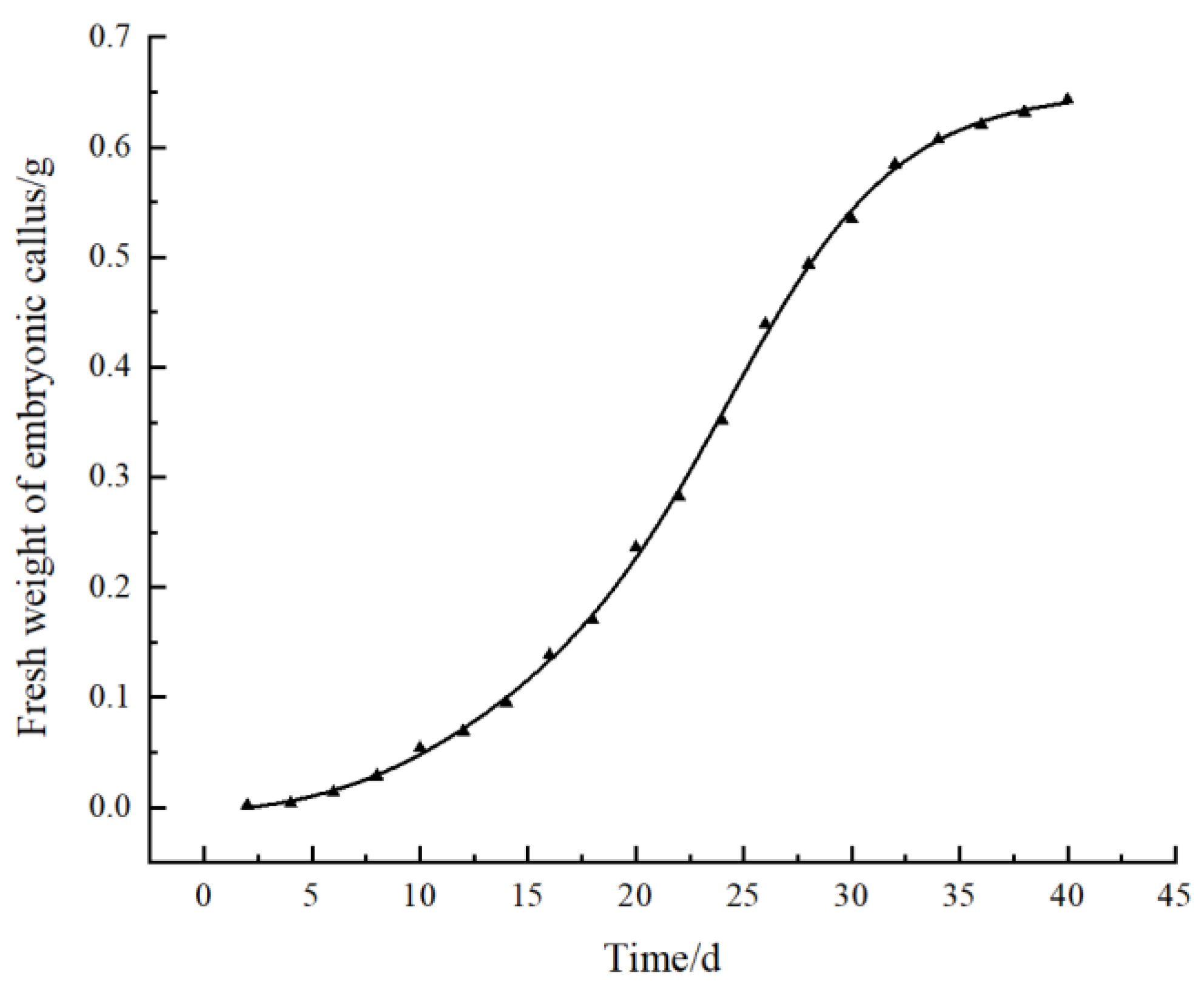
3.4. Effect of Subculture Cycle on Embryogenic Callus Proliferation
3.5. Effects of Different Concentrations of 6-BA and NAA on Somatic Embryo Induction
3.6. Rooting and Regeneration of Somatic Embryo Seedlings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Caruso, A.; Barbarossa, A.; Tassone, A.; Ceramella, J.; Carocci, A.; Catalano, A.; Basile, G.; Fazio, A.; Iacopetta, D.; Franchini, C.; et al. Pomegranate: Nutraceutical with Promising Benefits on Human Health. Appl. Sci. 2020, 10, 6915. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, X.; Ni, H.; Yuan, F.; Zhang, S. Identification and Functional Analysis of CAD Gene Family in Pomegranate (Punica granatum). Genes 2023, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, J.; Qin, G. Genome-wide distribution of simple sequence repeats in pomegranate and their application to the analysis of genetic diversity. Tree Genet. Genom. 2020, 16, 36. [Google Scholar] [CrossRef]
- Xia, X.; Fan, M.; Liu, Y.; Chang, X.; Wang, J.; Qian, J.; Yang, Y. Genome-wide alternative polyadenylation dynamics underlying plant growth retardant-induced dwarfing of pomegranate. Front. Plant Sci. 2023, 14, 1189456. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, S.; Qin, C.; Li, Q.; Cai, J.; Sun, C.; Wang, W.; Weng, Y. Somatic embryogenesis in mature zygotic embryos of Picea pungens. Sci. Rep. 2021, 11, 19072. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romero, C. Somatic embryogenesis in Olea spp. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 138, 403–426. [Google Scholar] [CrossRef]
- Park, Y.-S. Conifer Somatic Embryogenesis and Multi-Varietal Forestry. In Challenges and Opportunities for the World’s Forests in the 21st Century; Fenning, T., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 425–439. [Google Scholar] [CrossRef]
- Corredoira, E.; Ballester, A.; Ibarra, M.; Vieitez, A.M. Induction of somatic embryogenesis in explants of shoot cultures established from adult Eucalyptus globulus and E. saligna × E. maidenii trees. Tree Physiol. 2015, 35, 678–690. [Google Scholar] [CrossRef]
- Pais, M.S. Somatic Embryogenesis Induction in Woody Species: The Future After OMICs Data Assessment. Front. Plant Sci. 2019, 10, 240. [Google Scholar] [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, C.; Wang, H.; Ma, X.; Shen, H.; Yang, L. Indirect somatic embryogenesis and regeneration of Fraxinus mandshurica plants via callus tissue. J. For. Res. 2021, 32, 1613–1625. [Google Scholar] [CrossRef]
- Yan, R.; Sun, Y.; Sun, H. Current status and future perspectives of somatic embryogenesis in Lilium. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 143, 229–240. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.-G.; Fan, X.-F.; Su, Z.-H. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef]
- Baskaran, P.; Van Staden, J. Plant regeneration via somatic embryogenesis in Drimia robusta. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 119, 281–288. [Google Scholar] [CrossRef]
- Khan, T.; Reddy, V.S.; Leelavathi, S. High-frequency regeneration via somatic embryogenesis of an elite recalcitrant cotton genotype (Gossypium hirsutum L.) and efficient Agrobacterium-mediated transformation. Plant Cell Tissue Organ Cult. (PCTOC) 2010, 101, 323–330. [Google Scholar] [CrossRef]
- Lu, L.; Holt, A.; Chen, X.; Liu, Y.; Knauer, S.; Tucker, E.J.; Sarkar, A.K.; Hao, Z.; Roodbarkelari, F.; Shi, J.; et al. miR394 enhances WUSCHEL-induced somatic embryogenesis in Arabidopsis thaliana. New Phytol. 2023, 238, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Syeed, R.; Mujib, A.; Malik, M.Q.; Gulzar, B.; Zafar, N.; Mamgain, J.; Ejaz, B. Direct somatic embryogenesis and flow cytometric assessment of ploidy stability in regenerants of Caladium × hortulanum ‘Fancy’. J. Appl. Genet. 2022, 63, 199–211. [Google Scholar] [CrossRef]
- Xiong, Y.; Wei, Z.; Yu, X.; Pang, J.; Zhang, T.; Wu, K.; Ren, H.; Jian, S.; Teixeira da Silva, J.A.; Ma, G. Shoot proliferation, embryogenic callus induction, and plant regeneration in Lepturus repens (G. Forst.) R. Br. Vitr. Cell. Dev. Biol. Plant 2021, 57, 1031–1039. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, F.; Wang, Y.; Li, X.; Liang, H. Efficient Somatic Embryogenesis and Plant Regeneration from Immature Embryos of Tapiscia sinensis Oliv., an Endemic and Endangered Species in China. HortScience Horts 2014, 49, 1558–1562. [Google Scholar] [CrossRef]
- Bernula, D.; Benkő, P.; Kaszler, N.; Domonkos, I.; Valkai, I.; Szőllősi, R.; Ferenc, G.; Ayaydin, F.; Fehér, A.; Gémes, K. Timely removal of exogenous cytokinin and the prevention of auxin transport from the shoot to the root affect the regeneration potential of Arabidopsis roots. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 327–339. [Google Scholar] [CrossRef]
- Binte Mostafiz, S.; Wagiran, A. Efficient Callus Induction and Regeneration in Selected Indica Rice. Agronomy 2018, 8, 77. [Google Scholar] [CrossRef]
- Ming, N.J.; Binte Mostafiz, S.; Johon, N.S.; Abdullah Zulkifli, N.S.; Wagiran, A. Combination of Plant Growth Regulators, Maltose, and Partial Desiccation Treatment Enhance Somatic Embryogenesis in Selected Malaysian Rice Cultivar. Plants 2019, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.-W.; Yan, Y.-Y.; Liu, Y.-M.; Liu, Y.-M.; Deng, Y.-W.; Yao, H.-Y. The regeneration of Acer rubrum L. “October Glory” through embryonic callus. BMC Plant Biol. 2020, 20, 309. [Google Scholar] [CrossRef] [PubMed]
- Teixeira da Silva, J.A.; Rana, T.S.; Narzary, D.; Verma, N.; Meshram, D.T.; Ranade, S.A. Pomegranate biology and biotechnology: A review. Sci. Hortic. 2013, 160, 85–107. [Google Scholar] [CrossRef]
- Qian, J.; Wang, N.; Ren, W.; Zhang, R.; Hong, X.; Chen, L.; Zhang, K.; Shu, Y.; Hu, N.; Yang, Y. Molecular Dissection Unveiling Dwarfing Effects of Plant Growth Retardants on Pomegranate. Front. Plant Sci. 2022, 13, 866193. [Google Scholar] [CrossRef] [PubMed]
- Shin, U.; Chandra, R.; Kang, H. In vitro and Ex vitro Propagations of Astilboides tabularis (Hemsl.) Engl. as a Rare and Endangered Species. Hortic. J. 2019, 6, 261. [Google Scholar]
- Hazrati, R.; Zare, N.; Asghari-Zakaria, R.; Sheikhzadeh, P.; Johari-Ahar, M. Factors affecting the growth, antioxidant potential, and secondary metabolites production in hazel callus cultures. AMB Express 2022, 12, 109. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors Influencing Somatic Embryogenesis Induction and Plant Regeneration with Particular Reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Fehér, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, Q.; Kita, D.; Huang, J.-B.; Liu, G.; Wu, X.; Zhu, X.; Cheung, A.Y.; Wu, H.-M.; Tao, L.-Z. RopGEF1 Plays a Critical Role in Polar Auxin Transport in Early Development. Plant Physiol. 2017, 175, 157–171. [Google Scholar] [CrossRef]
- Kantharajah, A.; Dewitz, I.; Jabbari, S. The effect of media, plant growth regulators and source of explants on in vitro culture of pomegranate (Punica granatum L.). Erwerbsobstbau 1998, 40, 54–58. [Google Scholar]
- Vinterhalter, B.; Mitić, N.; Vinterhalter, D.; Uzelac, B.; Krstić-Milošević, D. Somatic embryogenesis and in vitro shoot propagation of Gentianautriculosa. Biologia 2016, 71, 139–148. [Google Scholar] [CrossRef]
- Jaidka, K.; Mehra, P.N. Morphogenesis in Punica granatum (pomegranate). Can. J. Bot. 1986, 64, 1644–1653. [Google Scholar] [CrossRef]
- Yang, Z.; Ludders, P. Organogenesis of Punica granatum L. var. nana. Angew. Bot. 1993, 67, 151–156. [Google Scholar]
- Ren, Y.; Yu, X.; Xing, H.; Tretyakova, I.N.; Nosov, A.M.; Yang, L.; Shen, H. Interaction of Subculture Cycle, Hormone Ratio, and Carbon Source Regulates Embryonic Differentiation of Somatic Cells in Pinus koraiensis. Forests 2022, 13, 1557. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Basal Medium | Plant Growth Regulators (mg/L) | Callus Induction Percentage (%) | Callus Growth Status | ||
|---|---|---|---|---|---|---|
| 6-BA | NAA | IBA | ||||
| A1 | WPM | 1.0 | 1.0 | 0.0 | 67.78 ± 6.94 b | Milky white, not dense |
| A2 | WPM | 1.0 | 0.0 | 0.5 | 81.11 ± 5.09 a | Light brown, dense |
| A3 | WPM | 0.0 | 1.0 | 0.5 | 65.56 ± 3.85 b | Grayish white, not dense |
| A4 | MS | 1.0 | 1.0 | 0.0 | 72.22 ± 5.09 ab | Light yellow, slightly dense |
| A5 | MS | 1.0 | 0.0 | 0.5 | 66.67 ± 3.33 b | White, dense |
| A6 | MS | 0.0 | 1.0 | 0.5 | 68.89 ± 6.94 b | Milky white, not dense |
| Medium | Plant Growth Regulator (mg/L) | Embryogenic Callus Induction Rate (%) | Callus Growth State | |
|---|---|---|---|---|
| 6-BA | NAA | |||
| B1 | 0.5 | 0.5 | 14.44 ± 1.92 e | First yellow, slowly brown, grew slowly, not dense |
| B2 | 0.5 | 1.0 | 74.44 ± 5.09 a | Light yellow to light green, grew fast, not dense |
| B3 | 0.5 | 1.5 | 0 ± 0 f | Brown, did not grow well |
| B4 | 1.0 | 0.5 | 35.56 ± 5.09 c | Light yellow, partially brown, modest growth, modestly dense |
| B5 | 1.0 | 1.0 | 61.11 ± 8.39 b | White, first grew fast and then grew slowly, dense |
| B6 | 1.0 | 1.5 | 25.56 ± 5.09 d | Yellow, slow grew, not dense |
| B7 | 1.5 | 0.5 | 7.78 ± 1.92 ef | White, grew slowly, dense |
| B8 | 1.5 | 1.0 | 43.33 ± 6.67 c | Milky white, grew modestly, not dense |
| B9 | 1.5 | 1.5 | 2.22 ± 3.85 f | First white, slowly brown, almost did not grow |
| Medium | Plant Growth Regulators (mg/L) | Somatic Embryo Induction Percentage (%) | Number of Somatic Embryoids Formed from One Callus Explant | |
|---|---|---|---|---|
| 6-BA | NAA | |||
| C1 | 0.1 | 0.1 | 0 ± 0 e | 0 ± 0 e |
| C2 | 0.1 | 0.5 | 9.33 ± 2.31 e | 1.17 ± 0.29 d |
| C3 | 0.1 | 1.0 | 9.33 ± 4.62 e | 1.67 ± 0.33 c |
| C4 | 0.5 | 0.1 | 20.00 ± 4.00 d | 1.85 ± 0.13 c |
| C5 | 0.5 | 0.5 | 78.67 ± 6.11 a | 2.83 ± 0.15 a |
| C6 | 0.5 | 1.0 | 57.33 ± 6.11 b | 2.26 ± 0.41 b |
| C7 | 1.0 | 0.1 | 22.67 ± 4.62 d | 1.74 ± 0.05 c |
| C8 | 1.0 | 0.5 | 50.67 ± 6.11 b | 1.80 ± 0.17 c |
| C9 | 1.0 | 1.0 | 40.00 ± 10.58 c | 1.45 ± 0.17 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Xia, X.; Qin, G.; Tang, J.; Wang, J.; Zhu, W.; Qian, M.; Li, J.; Cui, G.; Yang, Y.; et al. Somatic Embryogenesis and Plant Regeneration from Stem Explants of Pomegranate. Horticulturae 2023, 9, 1038. https://doi.org/10.3390/horticulturae9091038
Wang J, Xia X, Qin G, Tang J, Wang J, Zhu W, Qian M, Li J, Cui G, Yang Y, et al. Somatic Embryogenesis and Plant Regeneration from Stem Explants of Pomegranate. Horticulturae. 2023; 9(9):1038. https://doi.org/10.3390/horticulturae9091038
Chicago/Turabian StyleWang, Jingting, Xinhui Xia, Gaihua Qin, Jingwen Tang, Jun Wang, Wenhao Zhu, Ming Qian, Jiyu Li, Guangrong Cui, Yuchen Yang, and et al. 2023. "Somatic Embryogenesis and Plant Regeneration from Stem Explants of Pomegranate" Horticulturae 9, no. 9: 1038. https://doi.org/10.3390/horticulturae9091038
APA StyleWang, J., Xia, X., Qin, G., Tang, J., Wang, J., Zhu, W., Qian, M., Li, J., Cui, G., Yang, Y., & Qian, J. (2023). Somatic Embryogenesis and Plant Regeneration from Stem Explants of Pomegranate. Horticulturae, 9(9), 1038. https://doi.org/10.3390/horticulturae9091038