Abstract
Kohlrabi (Brassica oleracea var. gongylodes) is a vegetable cultivated for its edible stem tuber. Although valued for its nutritional properties and tolerance to abiotic stress, kohlrabi is one of the least studied brassicas. In this review, we summarize the results of our decade-long research on in vitro morphogenesis of kohlrabi, starting from 2013. Protocols for efficient in vitro regeneration with minimal requirements for external application of plant growth regulators (PGRs) have been developed, both by somatic embryogenesis (SE) and by de novo shoot organogenesis (DNSO). Efficient regeneration by direct SE was achieved using immature zygotic embryos as explants incubated on PGR-free media, and the production process was maintained in culture thanks to highly efficient secondary SE. Conversely, efficient regeneration by indirect DNSO was achieved using entire seedlings as explants and adding only cytokinin (CK) without the need for exogenous auxin. Comprehensive phytohormone analyses revealed that different exogenously applied CKs differentially affected the composition of endogenous phytohormones and induced changes in the expression of cell cycle-related genes and other genes involved in the organogenic response. The addition of high sucrose concentrations to the nutrient media failed to induce the formation of stem tubers in in vitro culture, but revealed a complex interaction with exogenously applied CKs, interfering with both the endogenous phytohormonome and the expression of organogenesis-related genes. Our work has provided substantial biotechnological advances in the field of in vitro regeneration of kohlrabi, as well as in understanding the underlying phytohormonal regulation. The review aims to introduce kohlrabi to the scientific community as a model system for both basic and applied research, while we continue seeking answers to the outstanding questions and trying to pave the way for the development of more resistant varieties.
1. Kohlrabi: Brassica’s Hidden Gem
Kohlrabi (Brassica oleracea var. gongylodes), a low, biennial plant in the Brassicaceae family, is grown for the edible tuber that develops on the lower part of the stem. The interior of the tuberous flesh is always white, regardless of the white, purple, or green exterior color of the plant stem [1].
Similarly to most brassicas, kohlrabi is characterized by high nutritive and medicinal value due to its high content of fibers, vitamin C, and other vitamins (A, B1, B2, B5, B6, and E), minerals (K, Ca, Mg, Zn, and Fe), and antioxidants with anticancer activity [2,3]. However, unlike some other brassica crops distinguished by a bitter taste, kohlrabi has a luscious and mellow taste, presumably due to the higher content of hydrophilic amino acids [4]. Also, kohlrabi flesh is rich in starch, sucrose, and reducing sugars, primarily fructose and glucose [1,4,5].
With a short vegetation period and greater resistance to low temperatures (especially in purple varieties) and drought compared to other crucifers, kohlrabi is able to grow in less favorable areas, which makes it a suitable alternative to more economically popular family members [6,7]. A comparative analysis of the nutritive, structural, and functional features of kohlrabi and radish (Raphanus sativus) confirmed a higher total level of free amino acids as well as essential amino acids in kohlrabi, pointing to the better nutritional value of this crop [4]. Additionally, kohlrabi stems were shown to contain the most glucoiberverin and glucoiberin among 16 different types of identified glucosinolates, which is significant from a nutritional perspective [8]. Compared to examined edible parts of other Brassica oleracea subspecies, kohlrabi had remarkable amounts of glucoraphanin, which can be further transformed to isocyanate sulforaphane [8,9], known for its anticarcinogenic properties [10]. Shetty and co-authors [11] analyzed the content of kohlrabi peroxidases, and the results indicated that kohlrabi could serve as a possible alternative source of these enzymes that have great commercial importance nowadays.
All these features are considered highly beneficial for preserving human health. Being more favorable in taste than most other brassicas and comprising a large variety of functional compounds, it is not surprising that kohlrabi has been recently receiving more and more attention both as food and feed [3]. In this context, the number of studies on ex vitro grown kohlrabi has increased, including studies on the metabolism of phenolic compounds and glucosinolates [8,12], their antioxidant and antimicrobial potential [13], biosynthesis of anthocyanin [14,15], effects of plant growth regulators and various means of biofortification on growth, yield, and quality traits of kohlrabi [16,17,18], as well as reports on transcriptomic [19,20] and hormonomic [21] analysis of stem tuber formation.
However, several problems are encountered when it comes to the cultivation of kohlrabi. The most significant are weight loss, excessive hardening during winter storage, and disease development [1]. Kohlrabi is susceptible to numerous pests and pathogens such as bacteria, insects, and fungi, including Rhizoctonia solani and Plasmodiophora brassicae, causing the bottom rot disease [22] and the clubroot disease [23], respectively. Thus, as a complement to conventional breeding, biotechnological means could be the solution to improve existing traits in dealing with different pests and pathogens to develop resistant varieties. The use of such methods, like genetic engineering, genuinely requires an efficient and reproducible system for in vitro growth and regeneration as a prerequisite.
2. In Vitro Growth and Regeneration as a Biotechnological Approach for Studying and Improvement of Kohlrabi
The key characteristic of plant cells for their regeneration potential is totipotency [24,25,26]. This provides the possibility for the development of complete plants from isolated organs or their smaller fragments, even from single cells, under specific conditions. This means that plant cells can regain totipotency, but they are not totipotent per se. Or, to put it another way, “not all plant cells are totipotent, but under appropriate conditions, certain cells may become totipotent” [25].
Regeneration in plant tissue culture in vitro provides the best evidence for the existence of totipotency. By culturing plant cells, tissues, organs, and their parts (explants) under controlled physical and chemical conditions in vitro, a large number of vegetative clones of the mother plant can be obtained. Identification of the conditions for the expression of totipotency of plant cells in vitro is difficult and still is mostly of empirical character.
The in vitro development and regeneration of plants depend on various factors, such as plant species, genotype, age and developmental stage of explants, and physiological state of the mother plant serving as the explant source [27]. Very important factors are also the composition of the nutrient medium and the culture conditions, such as light and temperature. The interplay of all these factors leads to the induction and expression of a specific pathway of dedifferentiation/transdifferentiation of cells and tissues, or in other words, cellular reprogramming, resulting in the development of a new plant [28,29,30]. Plant regeneration in vitro proceeds through two main morphogenetic pathways: somatic embryogenesis (SE) and de novo shoot organogenesis (DNSO). Both pathways play important roles in the development of whole regenerated plants in in vitro culture, as well as in protocols for plant transformation, mutagenesis, polyploidization, and germplasm storage, and can also serve as useful experimental systems for studying the regulatory mechanisms of plant development [31].
The conditions for plant regeneration in vitro in numerous members of the genus Brassica have been intensively studied in recent decades (reviewed in [32,33,34]). Somatic embryogenesis was described in B. napus [35,36], B. oleracea [37,38,39,40], B. rapa [41], B. carinata [42,43], B. repanda [44], and B. juncea [45] using immature zygotic embryos, hypocotyls, cotyledons, leaves or roots as source plant material.
In many Brassica species, DNSO is considered the fundamental tool for plant regeneration in vitro [32]. Several protocols have been developed using different starting plant materials such as hypocotyls [38,46,47,48], leaves [49,50], roots [51], cotyledons [52,53], protoplasts [54], petioles [55], and floral segments [56].
For a long time, the only successful protocol for kohlrabi regeneration in vitro was published in 1988 by Glendening and Sjölund [49] and required the application of extremely high concentrations of cytokinin N6-benzyladenine (BA; 10 mg L−1). However, the application of high concentrations of PGRs often leads to numerous side effects such as inhibition of rhizogenesis, acclimatization problems, or induction of morphological abnormalities [57]. Additional studies were carried out with microspore culture, but the procedure was complicated and associated with a lower regeneration rate [58].
In the last decade, our team succeeded for the first time in establishing and optimizing protocols for efficient regeneration of kohlrabi in vitro, using both somatic embryogenesis and de novo shoot organogenesis [59,60] (Figure 1). Since then, this system has been used productively as a model system for studying plant morphogenesis in vitro [61,62,63,64].
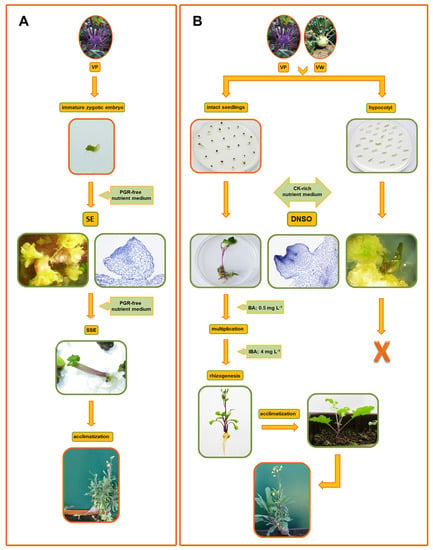
Figure 1.
Schematic representation of protocols for efficient regeneration of kohlrabi in vitro, via somatic embryogenesis (SE) and de novo shoot organogenesis (DNSO). (A) Direct SE from immature zygotic embryos of cv. Vienna Purple (VP), where the highest regeneration frequency was obtained on a nutrient medium without plant growth regulators (PGRs). Secondary somatic embryogenesis (SSE) occurred on primary embryos used as explants without the need for PGRs. The embryos developed into normal diploid plantlets that were subsequently successfully acclimatized. (B) Indirect DNSO from hypocotyl explants and intact seedlings of VP and cv. Vienna White (VW) with successful regeneration in both tissue types on cytokinin (CK)-rich nutrient medium. Shoots that regenerated from intact seedlings were viable and multiplied well on a medium containing N6-benzyladenine (BA; 0.5 mg L−1). They were further effectively rooted using indole-3-butyric acid (IBA; 4 mg L−1) and acclimatized. Histological analyses confirmed two individual ways of kohlrabi regeneration. The red frames indicate the distinct starting material that successfully followed the path to develop a whole plant.
3. Direct Somatic Embryogenesis from Kohlrabi Immature Zygotic Embryos
A prerequisite for establishing a successful system for somatic embryogenesis implies a good selection of plant material, that is, explants need to be a source of competent cells. On the other hand, the determination of the physical and chemical factors that induce the embryogenic path of development is also very important [65,66,67].
Somatic embryogenesis can be initiated from different types of explants, such as whole seedlings and their fragments: petioles, leaves, roots, cotyledons, and shoot meristems, as well as seeds or zygotic embryos. The different types of explants used to induce SE show the greatest response at a certain age [35,68]. As with zygotic embryos, the higher embryogenic potential is most commonly observed in younger zygotic embryos [69]. However, cases have also been observed in which embryogenic potential increases as zygotic embryos mature [65,70]. It has also been shown that the maturity of the zygotic embryo affects the type of somatic embryogenesis, i.e., more mature zygotic embryos enter the direct SE, while in younger ones, callus is first formed, from which somatic embryos subsequently develop [65,71,72].
Immature zygotic embryos represent the most commonly used source of embryogenic cells. Their use has allowed the induction of SE in plant species where it could not be obtained by other means [73,74]. In kohlrabi, cv. Vienna Purple, immature zygotic embryos showed high regeneration potential via direct and repetitive embryogenesis when grown at a temperature of 25 ± 2 °C and 16/8 h (light/dark) photoperiod [59] (Figure 1A).
The developmental stage of immature kohlrabi zygotic embryos significantly affected embryogenic competence, as shown for other members of the genus Brassica [28,35,65,75,76]. Zygotic embryos at the torpedo and late cotyledonary stages of development were less responsive, in contrast to explants at the early cotyledonary stage, for which the highest frequency of SE was recorded [59]. The observed discrepancy could be the consequence of a different physiological status of explant tissue at different developmental stages [76].
A high potential for SE in kohlrabi was recorded for hypocotyls of germinated immature zygotic explants [59]. This is consistent with results from other plant species, such as Rosa hybrida [77], Brassica napus [75], and Brassica carinata [43], suggesting that young hypocotyls are favorable targets for the induction of SE. Konar and co-authors [78] attributed this potential to epidermal hypocotyl cells that have retained meristem characteristics, i.e., regeneration capacity. On the other hand, similarly to DNSO [79], pericycle-like stem cells could be possibly responsible for this high regeneration capacity; however, this is still a matter of debate [25].
The key factors for the acquisition of embryogenic competence are auxins and cytokinins (CKs), which is related to their involvement in the cell cycle and control of cell division [80]. Hence, their exogenous application, particularly of auxin, is generally considered a prerequisite for successful SE induction in vitro [27,81,82].
The most commonly used auxin in SE induction is 2,4-dichlorophenoxyacetic acid (2,4-D) [73], which has been shown to be particularly effective in numerous plant species [28,39,65,83]. However, 2,4-D negatively affected the SE of kohlrabi [59], resulting in a decrease in SE efficiency in immature zygotic embryos, while also having no stimulatory effect on further somatic embryo development. Both processes were more effective on a medium without growth regulators (Figure 1A). Moreover, the presence of 2,4-D induced irregular growth and deformation of explants, regardless of their developmental stage, and the degree of deformity increased with increasing 2,4-D concentration. A compact callus was observed only in a smaller number of explants grown at higher 2,4-D concentrations, and in other cases, somatic embryos developed directly on the explant, without an intermediate callus phase [59].
2,4-D also acts as a strong stressor [68]. Hence, its use in the medium for SE induction may have deleterious effects, such as the development of malformed embryos [84,85]. This is thought to be a consequence of the disruption of the polar transport of endogenous auxins as a result of exogenous auxin application [86]. Application of auxin may also result in inhibition of the development of embryogenic masses into somatic embryos, by either disrupting cell polarity or preventing the formation of an auxin gradient [80,87].
Somatic embryogenesis on media without PGRs is a rare event [28,68], thus, a SE frequency of more than 80% on a PGR-free medium obtained in kohlrabi can be considered exceptional [59]. In most species that do not require the addition of PGRs for the induction of SE, endogenous levels of auxins and CKs were crucial factors in determining the embryogenic response [28,80,88].
Secondary somatic embryogenesis (SSE) occurs when new (secondary) somatic embryos arise from somatic embryos as explants [73]. This phenomenon provides a potential long-term source of somatic embryos and is one of the most advantageous techniques for in vitro multiplication, regeneration, and genetic transformation in certain species [89,90,91]. The presence of auxin is required for the induction of SSE in most cases. Some systems require a combination of auxin with CKs [91,92] or SSE occurs in the absence of PGRs [35,93]. In kohlrabi, secondary embryos also appeared spontaneously on primary embryos during cultivation on a substrate lacking growth regulators (Figure 1A). Exogenously applied CKs generally did not significantly increase the frequency of secondary embryogenesis [59].
The majority of plant species show a higher frequency of SSE compared to the induction of primary SE [73,91,94]. Furthermore, some cultures are able to maintain competence for secondary embryogenesis for many years [90,95,96]. Comparable results were obtained in kohlrabi research, where cultures retained high embryogenic potential after one year of repetitive SE on a nutrient medium without PGRs [59]. The highest efficiency of SSE was recorded in embryogenic lines derived from zygotic explants at the early cotyledonary stage. This efficiency was almost three times higher than in the primary SE from the same explants, presumably due to the use of young tissue as starting material. Overall, the results indicate the possibility of generating a large number of embryos in a relatively short time. All tested secondary embryos germinated into normal diploid plants (Figure 1A), indicating that the risk of somaclonal variation occurring during this regenerative procedure is very low.
The early concept of somatic embryogenesis assumed that a somatic embryo originates from a single cell [97], which has been shown in various plants such as carrot [98]. However, the unicellular origin of somatic embryos is not the rule, and there are examples of the simultaneous occurrence of embryos with unicellular and multicellular origins in one regenerative system [99]. Regenerative systems that can guarantee a unicellular origin of somatic embryos are thought to be very rare [25]. Somatic embryos formed from the callus, that is, through indirect SE, have a higher probability of a multicellular origin, whereas direct SE is more likely to result in somatic embryos from a single cell [28]. In most cases where PGRs were not used for embryogenic induction, somatic embryos are formed directly on the explant surface without callus formation [80]. Histological analysis indicated that the kohlrabi somatic embryos were presumably of unicellular origin and formed directly from the surface cells, that is, explant layers, without an intermediate callus phase [59].
Somatic embryos usually develop into small plants, similar to seedlings, on a nutrient medium without growth regulators. In many SE induction systems, a large number of embryos is formed, but many of them do not subsequently develop into plants. For the development of somatic embryos into plants, gibberellic acid [100,101], glutamine, or casein hydrolysate [87] are usually added to the medium. Indole-3-butyric acid (IBA), abscisic acid (ABA), or CKs are also used for these purposes [70,102,103]. In kohlrabi, 96% of somatic embryos spontaneously developed into young plants, and after one month of acclimatization in the greenhouse, the survival rate was 72.5% with no ploidy variation in acclimatized plants [59] (Figure 1A). The efficient conversion of embryos into plants is an important prerequisite for the application of this protocol in potential biotechnological kohlrabi breeding programs.
Establishing the efficient kohlrabi regeneration system as one of the model systems in which the addition of PGRs is not necessary for the induction of somatic embryogenesis may also help to elucidate early regulatory events in embryo development and provide insights that could apply to other plant groups.
4. One-Step De Novo Shoot Organogenesis in Kohlrabi
A balance of appropriate PGRs, namely auxin and cytokinin, in the nutrient medium is necessary for the initiation of the process of de novo organogenesis in plants. Back in the middle of the 20th century, Skoog and Miller demonstrated the regulatory effect of these hormones on the process of organogenesis in vitro, whereby their presence at certain concentrations in nutrient medium plays a role in determining the morphological path of the explant and the formation of the various organs [104,105]. A high ratio of auxin to low CKs promotes root formation, whereas the reverse situation, in which CKs prevail, leads to shoot formation. Moreover, not only the ratio but also the concentration of exogenously administered hormones plays an important role in directing cell differentiation during organogenesis toward the formation of shoots or roots [106]. Although the importance of Skoog and Miller’s research to plant regeneration is enormous, it has been shown that not all plants are equally sensitive to growth regulators [107]. A number of processes take place prior to the actual regeneration of the organ, so different treatments are required depending on the plant species [108,109].
In 1988, Valvekens and co-authors [110] developed a simple and efficient protocol for shoot regeneration from the root of A. thaliana by applying a series of callus induction medium containing high auxin and low CK concentrations, and, conversely, shoot induction medium with a ratio of high CK and low auxin. Since then, this two-step shoot regeneration practice has become standard and is widely used for the regeneration of numerous plant species [111].
However, when establishing and optimizing kohlrabi shoot regeneration in vitro, it was found that kohlrabi tissue was able to develop callus and subsequently de novo shoots when grown on medium only supplemented with CK (Figure 1B) and skipping preincubation with auxin [60]. This indirect one-step DNSO protocol was accomplished for both investigated cultivars—Vienna Purple (VP) and Vienna White (VW), at the same growth conditions as previously described for somatic embryogenesis induction. Exceptions to the basic two-step shoot regeneration methodology have been demonstrated in other plant species among conifers, dicots, and monocots, both as direct and indirect shoot organogenesis [112,113,114]. The advantage of such methodology, which excludes the exogenous application of auxin, is the shorter duration of the procedure itself, lower costs, and, most importantly, the avoidance of the negative effects that auxin may have in plant tissue culture [24]. Regardless of the presence or absence of auxin in regenerative media, it has been shown that the same processes are very likely to occur at the molecular level during DNSO stages in plant tissue competent for regeneration (reviewed in [30,111]).
The success of in vitro plant regeneration depends on the choice of plant tissue, that is, explant type [113]. Cotyledons, roots, and hypocotyls have been shown to be good starting materials for regeneration in many brassicas, with the hypocotyl generally considered the most competent [32,115,116,117,118]. Indeed, kohlrabi hypocotyl explants have been shown to be more successful than roots and cotyledons in shoot regeneration [60]. However, compared to hypocotyls, intact seedlings showed almost ten times higher regeneration frequency in both studied kohlrabi cultivars, especially when 5 mg L−1 BA or 2 mg L−1 thidiazuron (TDZ) were applied [60].
Shoots regenerated from kohlrabi seedlings appeared to be more viable, but interestingly, the average number of regenerated shoots per explant was lower in seedlings compared to those obtained from hypocotyl explants. Most studies have shown that explants derived from seedlings several days old have optimal regeneration frequencies. In B. rapa ssp. oleifera, three-day-old seedlings provided better material for regeneration than four-day-old ones [119,120]. Explants from four-day-old B. napus seedlings had a regeneration frequency of 90% [121], and in B. juncea, the highest shoot regeneration efficiency was obtained with explants from 3 to 5-day-old seedlings [122]. Moreover, the optimal age of hypocotyls for shoot production via DNSO has been shown to be up to seven days [48,118,123,124]. However, hypocotyl sections were excised from two-week-old seedlings in kohlrabi experiments because the younger seedlings were not sufficiently developed [60]. For similar reasons, hypocotyl explants isolated from nine-day-old [53] or two-week-old seedlings [125] have also been used for shoot regeneration in cabbage.
In many plant species, including members of the genus Brassica, BA and TDZ are considered the most efficient PGRs when it comes to de novo organogenesis of shoots, and for this reason, they are also the most commonly used CKs [47,50,126,127,128]. Synthetic CK-like growth regulators, such as TDZ, are thought to be able to influence plant morphogenesis differently than isoprenoid and aromatic CKs [129]. TDZ appears to be less sensitive to enzymatic degradation in vivo compared to natural CKs, so it may elicit a stronger response at low concentrations. In addition, TDZ is known to inhibit the activity of the CK catabolic enzyme, CK oxidase/dehydrogenase [130,131,132]. However, this growth regulator may also cause some adverse effects and abnormalities in plant tissues grown in vitro, such as inhibition of shoot elongation or hyperhydricity [133,134]. The high efficiency of both BA and TDZ was recorded in inducing DNSO in kohlrabi seedlings without significant side effects [60].
Similarly, trans-zeatin (transZ) induced high regeneration frequency in seedlings of both cultivars as well as in VW hypocotyls and was the most successful among exogenously applied CKs in terms of the number of de novo induced shoots from the VP hypocotyls. Other studies also showed a strong effect of transZ on shoot regeneration compared to other CKs [135,136,137,138].
For a long time, the physiological activity of zeatin was believed to be limited to transZ only, which is typically considered the most active of all CK nucleobases [139,140,141], whereas cis-zeatin (cisZ) was considered to have little or no activity at all [142,143]. Meanwhile, some recent discoveries indicated that the cis-isomers of zeatin may have unique physiological functions in plants after all [144,145,146,147]. Accordingly, regeneration of kohlrabi shoots, both from hypocotyls and intact seedlings, was also achieved on a medium containing cisZ, but cisZ proved to be significantly less active than transZ in inducing shoot regeneration in the case of kohlrabi seedlings [60].
Nutrient media supplemented with BA are considered the best media for in vitro propagation of shoots in numerous plant species [55,122,148,149,150]. Cultivation of regenerated kohlrabi shoots on 0.5 mg L−1 BA (Figure 1B) provided an additional opportunity to study differences in multiplication potential between shoots regenerated from hypocotyl explants and seedlings, as well as between pretreatments with different CKs [60]. The effect of CK pretreatment on the multiplication of shoots regenerated from seedlings differed between the two tested cultivars, with a noticeable impact in VP in contrast to VW, in which the effect of pretreatment was not pronounced. This difference between genotypes may be due to the differences in their metabolism. It is possible that the upper limit of tissue sensitivity to growth regulators in VW shoots was exceeded, which is why the expected stimulation of shoot multiplication was absent [60].
The ultimate goal for a successful regeneration protocol should comprise the efficient development of regenerated shoots into viable plants that are subsequently acclimatized to ex vitro conditions. To this end, the development of a robust root system is necessary, for which auxins are routinely used following shoot regeneration [151,152]. Application of IBA at a concentration of 4 mg L−1 (Figure 1B) increased the frequency of rhizogenesis as well as the number of roots formed per shoot in both kohlrabi cultivars, compared to 2 mg L−1 [60]. The average root length was also increased, but Pavlović and co-authors [48] found that the higher IBA concentration (4 mg L−1) had negative effects on parameters of rhizogenesis in different varieties of Brassica oleracea L. Abbasi and co-authors [50] showed the maximum response when 3 mg L−1 IBA was applied, while higher concentration had an inhibitory effect in B. rapa L. var. turnip as well. Hence, kohlrabi appears to be less sensitive to IBA than other Brassica species in which rhizogenesis has been studied. A well-developed root system enabled the successful acclimatization of kohlrabi plants (Figure 1B). All acclimatized plants continued to develop normally, flowered, and formed pods without any change in ploidy. All these indicate the success and efficiency of the regeneration protocol for kohlrabi through DNSO.
5. Hormonal Input during Growth and Regeneration of Kohlrabi In Vitro
Plant hormones act as integrating metabolites essential for the normal growth and functioning of plants. Their homeostasis and dynamics fluctuate in response to various external and internal stimuli and depend on the stage of plant development, as has been shown for the initial life phases of in vitro grown kohlrabi.
A comprehensive phytohormone analysis of VP kohlrabi seedlings during growth and shoot regeneration in vitro was performed for the first time by our group [60,63,64]. Depending on the type of investigated phytohormone and corresponding metabolite, individual changes were observed on a time scale (four developmental stages T1–T4, defined arbitrarily in [63]), showing specific hormone profiles (Figure 2A).
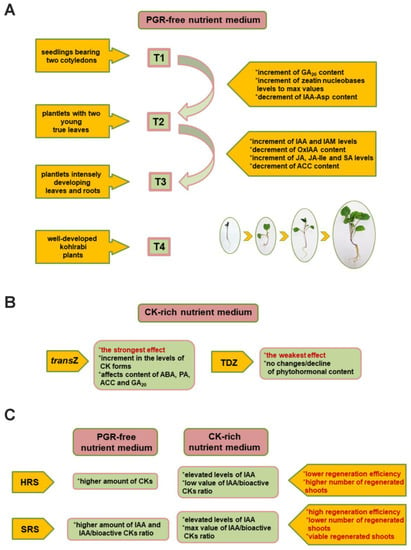
Figure 2.
Phytohormonal input during growth and de novo shoot organogenesis (DNSO) of kohlrabi in vitro. (A) Significant changes in phytohormonome during the four (T1–T4) developmental stages of kohlrabi seedlings grown on a nutrient medium without plant growth regulators (PGRs). (B) Main effects of exogenous application of cytokinins (CKs) on hormone levels. (C) Differences between two types of kohlrabi tissues used in the DNSO process along with regenerated shoots; additionally, the correlation between endogenous levels of CKs and auxins, the type of nutrient medium, and the regenerative potential of the explants used are indicated. GA20: gibberellin GA20; IAA: indole-3-acetic acid; IAA-Asp: IAA aspartate; OxIAA: 2-oxindole-3-acetic acid; JA: jasmonic acid; JA-Ile: (+)-7-iso-jasmonoyl-L-isoleucine; SA: salicylic acid; ACC: 1-aminocyclopropane-1-carboxylic acid; transZ: trans-zeatin; TDZ: thidiazuron; HRS: hypocotyl with regenerated shoots; SRS: seedling with regenerated shoots (terminology according to [60]).
Growth on PGR-free medium from the T1 stage represented by kohlrabi seedlings with two cotyledons to plantlets with two young and developing true leaves at the T2 stage resulted in a significant increase of the precursor of bioactive GAs, gibberellin GA20. Another significant change at T2 involved a decrease in the content of one of the most abundant indole-3-acetic acid (IAA) conjugates, IAA aspartate (IAA-Asp) [64]. In contrast, the content of zeatin nucleobases, comprising transZ, cisZ, and dihydrozeatin (DHZ), increased sharply, reaching the maximum value at T2. CK nucleobases are considered the bioactive CK forms in plants [153]. They are the least abundant of all CK groups [154,155], with kohlrabi seedlings being no exception [60,63], and with transZ mostly contributing throughout all developmental stages studied [64]. The strong increase of these bioactive CK forms argues for their specific role in controlling important processes in young developing kohlrabi seedlings, together with shaping the shoot apical meristem [156].
During the T3 developmental stage, kohlrabi plantlets continued to grow intensely developing leaves and roots. This growth stage was characterized by an increase in IAA levels (Figure 2A). Presumably, this was a consequence of the hydrolysis of IAA conjugates, which was consistent with lowered IAA-Asp levels at the previous stage [64]. As key growth regulators, auxins are involved in cell division and expansion, stimulation of lateral root initiation, and adventitious root formation, among others. Importantly, auxin also plays a critical role in regulating leaf growth and development [157,158]. Additionally, a significant drop in concentrations of the physiologically inactive IAA catabolite, 2-oxindole-3-acetic acid (OxIAA), argues for increased auxin action, since oxidation of IAA moderates IAA signaling [159].
The content of indole-3-acetamide (IAM), an inactive precursor of IAA, increased even more than IAA at this time. Several plant species have been shown to contain IAM [160], and large amounts of IAA have been detected in transgenic lines overexpressing the iaaM gene coding for tryptophan 2-monooxygenase, which catalyzes the conversion of Trp to IAM [161]. Furthermore, IAM hydrolases that convert IAM to IAA have been isolated from Arabidopsis and Nicotiana tabacum [162,163], suggesting that the IAM pathway is an alternative route of IAA biosynthesis in plants, possibly including kohlrabi.
At the same time, reaching the T3 stage, kohlrabi plantlets develop extensively, consuming nutrients and releasing exudates, resulting in a stressful setting. One of the key regulators of adaptation to these types of stresses and plant survival under such conditions is abscisic acid (ABA), which rapidly accumulates in plants exposed to environmental stress [164,165]. However, the expected response of ABA did not occur in kohlrabi [64]. Levels of ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC) declined, indicating possible elevated synthesis of ethylene as well as its expected release into the kohlrabi surroundings. On the other hand, kohlrabi plantlets at the T3 stage greatly accumulated active jasmonates ˗ jasmonic acid (JA) and (+)-7-iso-jasmonoyl-L-isoleucine (JA-Ile), as well as salicylic acid (SA) (Figure 2A). JA signaling in plants is closely associated with other hormones and can interact with ABA, ethylene, and SA as well as GAs in controlling both plant responses to stress and physiological processes. Depending on the context, these interactions can be either synergistic or antagonistic [166], thus the observed changes in kohlrabi are likely complex.
The content of endogenous plant hormones depends on the plant species and genotype, as well as on the type of explant and developmental stage [167], but is also strongly affected by the composition of PGRs in the nutrient medium [168,169,170,171]. CKs, applied in the regeneration media, initiated callogenesis and de novo shoot formation (corresponding to stages T3 and T4, respectively), while at the same time strongly affecting the kohlrabi phytohormonome, particularly endogenous CKs [60,63,64] (Figure 2B).
Exogenous transZ led to a significant increase in endogenous levels of all CK forms, including CK nucleobases, ribosides, O-glucosides, N-glucosides, and phosphates, throughout kohlrabi development, ultimately contributing to the enrichment of the total CK pool [60,63]. It can be assumed that this was predominantly due to uptake from the media, since the application of both naturally occurring CKs, transZ or cisZ, particularly contributed to an increase in the content of the same type of CKs as added to the nutrition medium (transZ- or cisZ-type, respectively). In addition, transZ induced a less pronounced increment of the endogenous content of cisZ-type CKs and vice-versa [60]. However, it is not clear whether this process is due to the possibility of interconversion between cis- and transZ, which has already been shown in several other studies [170,172,173].
Significant effect of transZ was also observed for ABA and its metabolite phaseic acid (PA), as well as for SA, 1-aminocyclopropane-1-carboxylic acid (ACC), and GA20 [64]. The observed relations suggest a complex network between growth regulators involved in the induction of shoot organogenesis and endogenous tissue-specific hormones, as well as a crosstalk among different phytohormone pathways, using a large number of confirmed and potential points of interaction [174].
In contrast to the transZ treatment, the total content of endogenous CKs was lowest when a synthetic diphenylurea-type CK, TDZ, was added to the nutrient medium [60]. The composition of endogenous CKs was similar to the control, and TDZ even led to a decrease in the concentrations of certain CK groups at certain developmental stages compared to the control [63,64]. A significant decline after TDZ treatment was also observed in other phytohormonal groups [64] (Figure 2B).
The principal component analysis confirmed that the difference between transZ and TDZ treatments was substantial at the first three analyzed stages in kohlrabi development, but greatly reduced as proceeding to the last stage [64]. The differences between the effects of transZ and TDZ can be attributed to their different modes of action. TDZ is able to competently bind receptors for CKs [175]; it can inhibit CKX [130,131,132] and consequently affect the metabolism of endogenous CKs [176]. TDZ can also affect the metabolic pathways of other phytohormones, such as auxin, ultimately disturbing their regulation [129].
Hormonal analysis of kohlrabi hypocotyl explants with regenerated shoots (HRSs) and intact seedlings with regenerated shoots (SRSs) after 6 and 8 weeks of in vitro culture, respectively, corresponding to a prolonged T4 stage, helped to clarify the correlation between the endogenous status of hormones, particularly CKs and auxins, and the application of an efficient CK-rich regeneration medium in relation to the regenerative potential of the explants used [60]. Differences were observed between HRSs and SRSs, with HRSs generally characterized by a higher amount of CKs compared to SRSs. However, SRSs contained more IAA and, as a consequence, a higher ratio of IAA to bioactive CKs (Figure 2C).
The exogenous CKs not only affected the endogenous levels of total CKs, but their application also significantly elevated the IAA content in both types of explants with regenerated shoots, regardless of the kohlrabi cultivar [60]. The results obtained for kohlrabi are consistent with the well-known fact that CKs influence both auxin biosynthesis as well as polar auxin transport in plants, although this influence is quite variable [177,178]. Several examples of the complex relationship between these two groups of hormones pointed to the possibility that the increase in endogenous IAA content in kohlrabi exposed to treatment with exogenous CKs was actually a result of CK-induced auxin biosynthesis [60].
During de novo organogenesis of shoots, specific changes occur in the expression of genes involved in auxin and cytokinin signaling pathways, as well as corresponding transcription factors [111,179,180]. Using the standard two-step in vitro regeneration protocol, it was shown that exogenous auxin initially mediates processes such as the acquisition of competence for organ formation, cell dedifferentiation, and the formation of callus and organogenic primordia within it. Then, exogenous CK from the nutrient medium to which explants are further transferred mediates the determination of the developmental pathway from organ primordia to the shoot [30,181]. Formation of local gradients of phytohormones is necessary for the activation of organogenesis-related events; CKs affect local changes in auxin levels and vice versa [178,182,183]. The endogenous auxin level in explant tissue has an important impact on regeneration viability. Arabidopsis mutants with high expression of genes involved in auxin biosynthesis exhibited high regenerative capacity even in protocols that exclude the first step of incubation on auxin-containing medium [184]. Thus, elevated endogenous auxin levels in kohlrabi hypocotyl explants and seedlings during cultivation on media to which CKs were added may have been satisfactory for initiating the first phase of organogenesis in these explants without the need to supplement growth media with auxin [60].
It has been shown that a high IAA/bioactive CK ratio is essential for DNSO induction in various plant species [185,186]. This ratio reached the maximum values in seedlings with regenerated shoots of both kohlrabi cultivars after treatments with BA, TDZ, or transZ [60]. The same treatments were those that elicited the best response in terms of regeneration frequency, so it seems that the efficiency of regeneration as well as the viability of regenerated kohlrabi shoots is indeed correlated with the high ratio of endogenous IAA to bioactive CKs (Figure 2C). In contrast, a significantly lower IAA/bioactive CKs ratio was observed in the hypocotyl explants, which regenerated more shoots compared with the seedling explants [60]. Thus, kohlrabi hypocotyls might have a greater potential for later stages of shoot regeneration compared with other plant parts thanks to the different composition of endogenous hormones. This could be the reason why hypocotyl explants on average produced a higher number of shoots than intact seedlings, even though the regeneration from kohlrabi seedlings was more successful overall.
Hence, a specific balance of CKs and auxins is necessary for organogenesis, and their continuous interaction depends on phytohormone signaling pathways that take place in intact plants [187]. Thus, the intact kohlrabi seedlings might represent a better source of efficient shoot regeneration compared with explants obtained from isolated tissues such as hypocotyls, cotyledons, or roots in which the interaction between endogenous phytohormones from different plant parts is excluded.
6. Genetic Insights into De Novo Shoot Organogenesis of Kohlrabi
The peculiar ability of kohlrabi seedlings to regenerate efficiently by one-step de novo shoot organogenesis raised the question of the activity of some important elements of the signaling cascade driving this process. Since the mechanisms of callus development from different tissue types of Arabidopsis, a closely related member of the Brassicaceae family [188], have been thoroughly characterized [79,189,190], it was possible to compare the genetic regulation of the one-step regeneration in kohlrabi with the previously described model system [61].
Coordinated cell divisions are the basis for the development of multicellular tissues, which occurs during the process of DNSO. The cell division cycle in plants is controlled by cyclins and cyclin-dependent kinases, CDKs [191,192]. CDKs play a central role in controlling cell cycle progression, with their activation regulated by binding to the appropriate cyclin at various checkpoints [193,194]. The activity of these complexes is tightly controlled during the cell cycle and organ development, and these complexes may also be involved in the regulation of signal transduction and differentiation [195,196].
Andersen and co-authors [197] showed that endogenous levels of auxin and CKs were altered in the case of CDKB2 loss-of-function mutation; tissue with impaired CDKB2 function failed to translate hormonal signals into proper development, even though the tissue was normal at the time of explant isolation. This led to the conclusion that coordination of the cell cycle and the developmental program by plant hormones is of exceptional importance. Besides, B2-type CDKs were found to be active in the shoot apical meristem and necessary for its functioning; their activity is closely related to the functions of classical meristem regulators [198]. On the other hand, CKs are thought to control the course of the cell cycle by affecting the phase-specific change of cyclin in complex with CDKs through binding to the ATP-binding site in CDKs [199]. They regulate the transition from G2 to the M phase [200,201] and increase the activity of the corresponding CDKs [202].
Under different CK treatments, regenerated kohlrabi shoots appeared with different dynamics, which was followed by the expression of two cell cycle-related genes, CDKB2;1 and CYCB2;4, whose expression change correlated with certain phases of de novo organogenesis [61]. A precise balance between cell divisions and transdifferentiation in different sections of the meristem is necessary to properly maintain the structure and function of this tissue. In this sense, it is essential that signals from meristematic regulators are correctly construed by the cellular regulatory machinery [30,203].
The inclusion of two auxin transporter genes (PIN3 and PIN4), a cytokinin response regulator (ARR5), and two shoot apical meristem-related transcription factors (CUC1 and RGD3) in the expression analysis, in addition to the two cell cycle-related genes analyzed, helped to obtain a broader image of mechanisms involved in DNSO of kohlrabi [61] (Figure 3A). The profiles of relative expression of selected genes were generally similar and exhibited a two-phase pattern in most CK treatments (Figure 3B). The first expression peak was detected typically after 7 days of cultivation on CK-supplemented media. Since all analyzed genes are thought to respond to CKs [30], this first peak was presumed to be a result of direct upregulation by CK from the regeneration medium. The second peak occurred in accordance with callus formation, especially in treatments with one of the two zeatin isomers. This reflects the acquisition of organogenic competence, typically characterized by strong upregulation of most organogenesis-related genes during the conventional two-step protocol for in vitro regeneration [30,109,181,204].
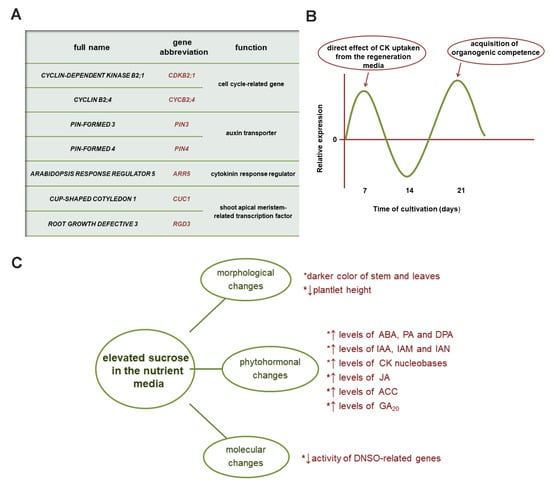
Figure 3.
(A) Analysis of organogenesis-related genes in kohlrabi cv. Vienna Purple (VP) de novo shoot organogenesis (DNSO) obtained by exogenous application of cytokinins (CKs). (B) Arbitrary illustration of the relative expression profile corresponding to selected genes displaying a two-phase pattern for the expression of most genes investigated in the majority of CK treatments. (C) Effects of increased sucrose application during growth and regeneration of kohrabi in vitro at morphological, phytohormonal, and molecular levels. ABA: abscisic acid; PA: phaseic acid; DPA: dihydrophaseic acid; IAA: indole-3-acetic acid; IAM: indole-3-acetamide; IAN: indole-3-acetonitrile; JA: jasmonic acid; ACC: 1-aminocyclopropane-1-carboxylic acid; GA20: gibberellin GA20.
Interestingly, the second upregulation of most organogenesis-related genes in BA and TDZ treatments followed the same time pattern as in cis- or transZ, despite the fact that calli formed earlier in BA- and TDZ-treated kohlrabi plantlets than in zeatin-treated ones [61]. Thus, it can be assumed that BA, similarly to TDZ has a different mode of action and affects the process of kohlrabi organogenesis differently than the isoprenoid CKs used in kohlrabi studies. These differences could also be the reason for the morphological dissimilarities between kohlrabi seedlings cultivated on different CK treatments [60,61]. The functional difference between TDZ, BA, and zeatin isomers during DNSO of kohlrabi remains a matter of debate until now. However, it is evident that the roles of organogenesis-related genes in the distinct phases of the regeneration process coincide in both one-step and classical two-step regeneration protocols, confirming their importance and universal role, at least in the Brassicaceae family.
7. Sucrose as an Important Component Affecting Multiple Aspects of Kohlrabi Growth and Regeneration In Vitro
Plant growth and development are coordinately controlled by exogenous and endogenous signals. On the one hand, phytohormones transmit and coordinate both external and internal cues to enable proper biological processes at physiological, morphological, and molecular levels. On the other hand, the availability of nutrients, predominately sugars, is necessary for normal plant growth. Sugars act as a carbon source and also as important signaling molecules that interact with other components of the plant regulatory machinery, such as plant hormones, affecting each other to varying extents [205].
In the development of an in vitro system for kohlrabi growth and regeneration, it was proven that the optimal concentration of sucrose in the growth media is 3% [59,60,62]. Since kohlrabi has the distinctive ability to form aboveground stem tubers, our team attempted to induce this process in vitro by applying higher sucrose concentrations (6% and 9%), among other potential regulators—fructose, glucose, xylose, and mannitol [62]. Presence of the latter sugar alcohol in nutrient media induced severe damage to kohlrabi seedling growth, to a much higher extent than the other used carbon sources [62]. Although tuber formation did not occur after the applied treatments, increased sugar availability was shown to affect kohlrabi at morphological, molecular, and phytohormonal levels (Figure 3C), with dynamic changes depending on the level of sucrose applied [62,63,64].
Kohlrabi plantlets (cv. Vienna Purple) cultivated on elevated sucrose tended to be shorter in height and developed stems and even leaves that were darker in color (Figure 3C), indicating anthocyanin accumulation, presumably as a result of sucrose-induced osmotic stress [62]. This effect is particularly pronounced at the later developmental stage, marked as T4 [63]. Carbohydrates are known to affect anthocyanin biosynthesis [206,207], as they act as signaling molecules. Sucrose is considered a key carbohydrate involved in the stimulation of the anthocyanin biosynthetic pathway [208] through the regulation of gene expression [206]. Moreover, a significant increase in JA levels in kohlrabi seedlings at the T4 stage grown at high sucrose concentrations [64] (Figure 3C) was likely involved in high sucrose-induced anthocyanin accumulation, as it has been shown that stress-induced anthocyanin accumulation is mediated by the JA signaling pathway [209].
While auxins, as a major group of phytohormones, interact extensively with other plant hormones, they have also been shown to importantly crosstalk with sugars and together tightly regulate certain processes in plants [205]. Sugars modulate both auxin formation and degradation; for example, sugar affects auxin biosynthesis [210,211,212]. Furthermore, glucose is known to stimulate IAA biosynthesis by affecting the levels of IAA precursors such as IAM and indole-3-acetonitrile (IAN) and consequently promoting the accumulation of IAA conjugates [213]. Consistent with this, endogenous levels of IAA, IAM, and IAN were elevated in kohlrabi grown at higher concentrations of sucrose [64]. Auxin is also related to starch accumulation [214], supporting the possible scenario that overloaded sucrose uptaken from the media is stored in kohlrabi plantlets.
Sucrose application affected other phytohormonal groups in kohlrabi, including the ethylene precursor ACC as well as ABA and its related metabolites PA and dihydrophaseic acid (DPA), increasing their levels, particularly at the last developmental stage studied, T4 [64] (Figure 3C). The previously proposed positive feedback loop between sugars and biosynthesis of ABA [215,216] supports the existence of a specific sugar regulation of ABA biosynthetic gene activity, leading to subsequent ABA accumulation in kohlrabi plantlets. On the other hand, the increased GA20 levels on sucrose treatments at the same stage were in contrast to the negative feedback loop suggested for gibberellin biosynthesis [217].
The presence of increased amounts of sucrose in the growth media additionally affected the CK content of VP kohlrabi [63,64], particularly with a significant increase in the levels of CK nucleobases at all stages studied [63] (Figure 3C). While the simultaneous use of exogenous CKs and high sucrose resulted in different relations in the content of various phytohormones, the most striking increase in CK content was induced when transZ and 9% sucrose were used together. Their pronounced synergistic effect is reflected particularly in the contents of zeatin nucleobases, at stages T3 and T4, which correspond to callus formation and shoot regeneration [63,64]. This suggested, as mentioned above, that the observed increase in CK levels could be the result of uptake from the nutrient medium, which in this case was further stimulated by the high sucrose concentration.
It is controversial whether phytohormone signaling depends entirely on endogenous hormone content in plant tissues, as affirmed in some cases [218]. Later, it was shown that endogenous plant hormones could be related to PGRs from regeneration media, whereby exogenous and endogenous plant hormones could coordinate to trigger callus formation and subsequently shoot organogenesis [219]. On the other hand, sugars are involved in the biosynthesis, transport, and signaling of CKs [220,221,222,223], and sugar-induced CK accumulation could be attributed to the upregulation of some IPT genes [224,225]. What mechanism allows the combined effects of naturally occurring zeatin and high amount of sugar to cause such remarkable accumulation, particularly of endogenous zeatin as the most bioactive CK form, is yet to be fully elucidated. It can be attributed to several potential factors, including osmotic stress, the activity of CK transporters, and impairment of cell membranes or other specifically induced signals.
Organogenesis-related genes selected for analysis during kohlrabi DNSO and with proven CK responsiveness [61] were significantly affected by a high concentration of sucrose solely, expressing only reduced activity [63] (Figure 3C). This reduced expression of organogenesis-related genes at 6% and 9% sucrose was clearly related to the observed delay of morphogenic events related to DNSO at high sucrose concentrations. Apparently, the stress associated with high sucrose concentration was able to significantly affect the efficiency of shoot regeneration [63].
In the case of exogenous transZ application, the combination with a high sugar concentration at the T4 stage led to the opposite effect, inducing the amplified expression of most of the investigated genes. The negative correlation between the content of CK nucleobases and the expression of most of the studied genes at both T3 and T4 stages in this treatment confirms the existence of a negative feedback loop that regulates high levels of endogenous CKs in kohlrabi as a consequence of transZ treatment [63].
The development of an efficient in vitro system for kohlrabi growth and DNSO allowed us the opportunity to study sugar metabolism in young kohlrabi plants, further investigating the effects of elevated sucrose and CKs application in the sense of stem tuber formation elucidation. Preliminary results of expression analysis of selected genes involved in sugar partitioning in plants seem to point to similar effects of both sucrose and CKs, suggesting an altered metabolism and transport of soluble sugars in kohlrabi (data not published).
8. Conclusions and Further Perspectives
Our work led to the development of protocols for the regeneration of kohlrabi (Brassica oleracea var. gongylodes) cultivars Vienna Purple and Vienna White, with minimal use of plant growth regulators and with remarkably high regeneration rates (up to 100% in SSE). We have shown that both secondary somatic embryogenesis from immature zygotic embryos incubated on growth media without PGRs and one-step de novo shoot organogenesis from whole seedlings grown on media containing only cytokinin can be used for efficient shoot regeneration and subsequent recovery of viable and fertile kohlrabi plants (Figure 4A). These regeneration protocols have the potential to be used in future biotechnological approaches to improve the characteristics of kohlrabi cultivars, particularly with respect to resistance to pests and pathogens, which remain the weakest links in commercial kohlrabi production.
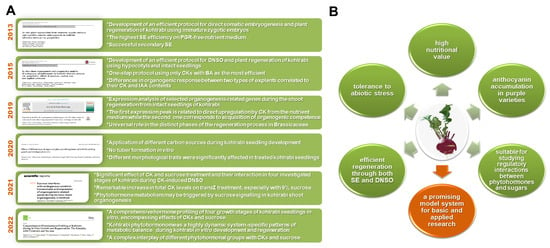
Figure 4.
(A) List of relevant publications [59,60,61,62,63,64] related to kohlrabi in vitro development and regeneration summarized in the present review with highlights from each performed investigation. (B) Schematic representation of significant traits that make kohlrabi an interesting model system for research. SE: somatic embryogenesis; PGR: plant growth regulator; DNSO: de novo shoot organogenesis; CK: cytokinin; BA: N6-benzyladenine; IAA: indole-3-acetic acid.
The physiological regulation of complex plant morphogenic responses to environmental cues such as PGRs and sucrose applied in in vitro culture can be studied in well-defined model systems, such as the one-step DNSO of kohlrabi. The tendency to accumulate carbohydrates in the form of starch in specialized organs—the stem tubers—makes kohlrabi an attractive model system for studying the regulatory circuits involving sugars and the phytohormones CK and auxin, which are involved in both shoot regeneration and assimilate partitioning. In the regeneration media, both CK and sucrose are uptaken by kohlrabi seedlings, leading to marked changes in the composition of their phytohormonome and the expression of CK-responsive genes involved in cell cycle regulation and shoot organogenesis. A particularly surprising outcome of the interaction between CK and sucrose is the dramatic (>150-fold) increase in endogenous transZ-type CKs when transZ is supplemented specifically in combination with 9% sucrose—the mechanism for this is currently unclear.
Moreover, as a “distant cousin” of the model species Arabidopsis thaliana which belongs to a distant clade of the same family Brassicaceae, kohlrabi is a perfect secondary model system to make comparisons with the thoroughly investigated regeneration and metabolism of Arabidopsis. The expression patterns of organogenesis-related genes confirmed that similar genetic mechanisms underlie both the two-step regeneration of Arabidopsis and the one-step regeneration of kohlrabi (Figure 4A). The highly efficient regeneration rates achieved for both SSE and DNSO in kohlrabi with minimal requirement for exogenous supplementation of PGRs could therefore serve as a starting point for understanding the universal factors that determine competence for highly efficient shoot regeneration in plants on a broader scale.
Taken together, kohlrabi, the Brassica’s hidden gem, remains a vegetable with many potentials (Figure 4B). Its high nutritional value, tolerance to abiotic stress, ability to accumulate high levels of anthocyanins in the stem of purple varieties, good suitability for regeneration through both somatic embryogenesis and shoot organogenesis, as well as suitability as a model system for studying regulatory interactions between phytohormones and sugars make it a promising model system for basic and applied research. The exciting journey to learn about kohlrabi has started, and will continue in many directions in the years to come.
Author Contributions
Conceptualization, T.Ć. and S.N.; Writing—original draft preparation, T.Ć. and M.R.; Writing—review and editing, T.Ć., V.M., M.R. and S.N.; Visualization, T.Ć. and A.C.; Supervision, S.N.; Project administration, S.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Ministry of Science, Technological Development and Innovations, Republic of Serbia, Grant No. 451-03-47/2023-01/200007 and by the Ministry of Education, Youth and Sports of the Czech Republic from European Regional Development Fund, Project “Centre for Experimental Plant Biology” (No. CZ.02.1.01/0.0/0.0/16_019/0000738).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Escalona, V.H.; Aguayo, E.; Artés, F. Extending the shelf life of kohlrabi stems by modified atmosphere packaging. J. Food Sci. 2007, 72, 308–313. [Google Scholar] [CrossRef]
- Hassan, E.A.; Hussein, A.; El-Awadi, M.E. Kohlrabi (Brassica caulorapa L): A potent anticancer vegetable processed in baked biscuits. Nat. Sci. 2011, 9, 149–157. Available online: http://www.sciencepub.net/nature/ns0908/27_6344ns0908_149_157.pdf (accessed on 25 April 2023).
- Paśko, P.; Galanty, A.; Tyszka-Czochara, M.; Żmudzki, P.; Zagrodzki, P.; Gdula-Argasińska, J.; Prochownik, E.; Gorinstein, S. Health Promoting vs. Anti-nutritive Aspects of Kohlrabi Sprouts, a Promising Candidate for Novel Functional Food. Plant Foods Hum. Nutr. 2021, 76, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Ryu, D.K.; Park, S.; Ahn, K.G.; Lim, Y.P.; An, G. Composition analysis between kohlrabi (Brassica oleracea var. gongylodes) and Radish (Raphanus sativus). Kor. J. Hort. Sci. Technol. 2010, 28, 469–475. Available online: http://www.koreascience.or.kr/article/JAKO201018651618927.page (accessed on 25 April 2023).
- Selman, I.W.; Kulasegaram, S. Development of the stem tuber in kohlrabi. J. Exp. Bot. 1967, 18, 471–490. [Google Scholar] [CrossRef]
- Uddin, J.; Solaiman, A.H.M.; Hasanuzzaman, M. Plant characters and yield of kohlrabi (Brassica oleracea var. gongylodes) as affected by different organic manures. J. Hort. Sci. Ornam. Plants 2009, 1, 1–4. Available online: https://idosi.org/jhsop/jhsop1(1)09.htm (accessed on 25 April 2023).
- Abd, A.M.; Alalawy, H.H.; Hathal, A.A. Response yield of four cultivar kohlrabi (Brassica oleracea var. caulorapa L.) to plant density and foliar nutrition of seaweed. Plant Arch. 2020, 20, 4069–4076. Available online: http://www.plantarchives.org/SPL%20ISSUE%2020-2/670__4069-4076_.pdf (accessed on 25 April 2023).
- Yi, G.E.; Robin, A.H.K.; Yang, K.; Park, J.I.; Kang, J.G.; Yang, T.J.; Nou, I.S. Identification and expression analysis of glucosinolate biosynthetic genes and estimation of glucosinolate contents in edible organs of Brassica oleracea subspecies. Molecules 2015, 20, 13089–13111. [Google Scholar] [CrossRef]
- Iori, R.; Bernardi, R.; Gueyrard, D.; Rollin, P.; Palmieri, S. Formation of glucoraphanin by chemoselective oxidation of natural glucoerucin: A chemoenzymatic route to sulforaphane. Bioorg. Med. Chem. Lett. 1999, 9, 1047–1048. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Shetty, P.; Banu, U.; Kedilaya, T.; Patheja, M. Characterization of peroxidase from Brassica oleracea gongylodes gives a lead for use of bromocresol purple as a novel substrate for peroxidase assay. J. Biochem. Technol. 2012, 4, 502–507. Available online: https://jbiochemtech.com/article/characterization-of-peroxidase-from-brassica-oleracea-gongylodes-gives-a-lead-for-use-of-bromocresol-purple-as-a-substrate-for-peroxidase-assay (accessed on 25 April 2023).
- Sathasivam, R.; Kim, M.C.; Yeo, H.J.; Nguyen, B.V.; Sohn, S.I.; Park, S.U.; Kim, J. Accumulation of Phenolic Compounds and Glucosinolates in Sprouts of Pale Green and Purple Kohlrabi (Brassica oleracea var. gongylodes) under Light and Dark Conditions. Agronomy 2021, 11, 1939. [Google Scholar] [CrossRef]
- Ben Sassi, A.; Ascrizzi, R.; Chiboub, W.; Mhamed, A.C.; ElAyeb, A.; Skhiri, F.; Saidani, M.T.; Mastouri, M.; Flamini, G. Volatiles, phenolic compounds, antioxidant and antibacterial properties of kohlrabi leaves. Nat. Prod. Res. 2022, 36, 3143–3148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, Z.; Zhu, M.; Zhu, Z.; Wang, Z.; Tian, S.; Chen, G. Antocyanin accumulation and molecular analysis of correlated genes in purple kohlrabi (Brassica oleracea var. gongylodes L.). J. Agric. Food Chem. 2015, 16, 4160–4169. [Google Scholar] [CrossRef]
- Rahim, M.A.; Robin, A.H.K.; Natarajan, S.; Jung, H.J.; Lee, J.; Kim, H.R.; Kim, H.T.; Park, J.I.; Nou, I.S. Identification and characterization of anthocyanin biosynthesis-related genes in kohlrabi. Appl. Biochem. Biotechnol. 2018, 184, 1120–1141. [Google Scholar] [CrossRef] [PubMed]
- Golob, A.; Novak, T.; Kacjan Maršić, N.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Lošák, T.; Válka, T.; Elbl, J.; Kintl, A.; Keutgen, A.; Keutgen, N.; Demková, L.; Árvay, J.; Varga, L.; Hnátková, H.; et al. Fertilization with Magnesium- and Sulfur-Supplemented Digestate Increases the Yield and Quality of Kohlrabi. Sustainability 2020, 12, 5733. [Google Scholar] [CrossRef]
- Bhandari, S.; Bhandari, A.; Shrestha, J. Effect of different doses of triacontanol on growth and yield of kohlrabi (Brassica oleracea L. var. gongylodes). Heliyon 2021, 7, e08242. [Google Scholar] [CrossRef]
- Hearn, D.J.; O’Brien, P.; Poulsen, T.M. Comparative transcriptomics reveals shared gene expression changes during independent evolutionary origins of stem and hypocotyl/root tubers in Brassica (Brassicaceae). PLoS ONE 2018, 13, e0197166. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sato, M.; Okamoto, S.; Hoque, M.; Shea, D.J.; Fujimoto, R.; Shimizu, M.; Fukai, E.; Okazaki, K. Comparative transcriptome analysis during tuberous stem formation in Kohlrabi (B. oleracea var. gongylodes) at early growth periods (seedling stages). Physiol. Plant. 2022, 174, e13770. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Kojima, M.; Sato, M.; Takebayashi, Y.; Hoque, M.; Okamoto, S.; Shea, D.J.; Fujimoto, R.; Fukai, E.; Sakakibara, H.; et al. Comparative anatomical and hormonal analyses between kohlrabi and broccoli seedlings: Relevance to kohlrabi stem tuber intiation. Sci. Hortic. 2023, 316, 112002. [Google Scholar] [CrossRef]
- Grosch, R.; Schneider, J.H.M.; Kofoet, A. Characterisation of Rhizoctonia solani anastomosis groups causing bottom rot in field-grown lettuce in Germany. Eur. J. Plant Pathol. 2004, 110, 53–62. [Google Scholar] [CrossRef]
- Ciaghi, S.; Schwelm, A.; Neuhauser, S. Transcriptomic response in symptomless roots of clubroot infected kohlrabi (Brassica oleracea var. gongylodes) mirrors resistant plants. BMC Plant Biol. 2019, 19, 288. [Google Scholar] [CrossRef]
- Neelakandan, A.K.; Wang, K. Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications. Plant Cell Rep. 2012, 31, 597–620. [Google Scholar] [CrossRef]
- Fehér, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef]
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant cell totipotency: Insights into cellular reprogramming. J. Integr. Plant Biol. 2021, 63, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New Insights Into Tissue Culture Plant-Regeneration Mechanisms. Front. Plant Sci. 2022, 13, 926752. [Google Scholar] [CrossRef] [PubMed]
- Gaj, M.D. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Shibata, M.; Rymen, B.; Iwase, A.; Bågman, A.M.; Watt, L.; Coleman, D.; Favero, D.S.; Takahashi, T.; Ahnert, S.E.; et al. A gene regulatory network for cellular reprogramming in plant regeneration. Plant Cell Physiol. 2018, 59, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Raspor, M.; Motyka, V.; Kaleri, A.R.; Ninković, S.; Tubić, L.; Cingel, A.; Ćosić, T. Integrating the roles for cytokinin and auxin in de novo shoot organogenesis: From hormone uptake to signaling outputs. Int. J. Mol. Sci. 2021, 22, 8554. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Cardoza, V.; Stewart, C.N. Brassica biotechnology: Progress in cellular and molecular biology. Vitr. Cell. Dev. Biol. -Plant 2004, 40, 542–551. [Google Scholar] [CrossRef]
- Vinterhalter, D.; Sretenović-Rajičić, T.; Vinterhalter, B.; Ninković, S. Genetic transformation of Brassica oleracea vegetables. Transgenic Plant J. 2007, 1, 340–355. Available online: http://www.globalsciencebooks.info/Online/GSBOnline/OnlineTPJ_1_2.html (accessed on 25 April 2023).
- Mitra, M.; Gantait, S. Tissue Culture-Mediated Biotechnological Advancements in Genus Brassica. In Brassica Improvement; Wani, S.H., Thakur, A.K., Jeshima Khan, Y., Eds.; Springer: Cham, Switzerland, 2020; pp. 85–107. ISBN 978-3-030-34694-2. [Google Scholar] [CrossRef]
- Koh, W.L.; Loh, C.S. Direct somatic embryogenesis, plant regeneration and in vitro flowering in rapid-cycling Brassica napus. Plant Cell Rep. 2000, 19, 1177–1183. [Google Scholar] [CrossRef]
- Zeynali, M.; Maleki Zanjani, B.; Amiri, M.E.; Noruzian, M.; Aghajari, S.M. Influence of genotype and plant growth regulator on somatic embryogenesis in rapeseed (Brassica napus L.). Afr. J. Biotechnol. 2010, 9, 4050–4055. Available online: https://www.ajol.info/index.php/ajb/article/view/82564 (accessed on 21 April 2023).
- Yang, J.L.; Seong, E.S.; Kim, M.J.; Ghimire, B.K.; Kang, W.H.; Yu, C.Y.; Li, C.H. Direct somatic embryogenesis from pericycle cells of broccoli (Brassica oleracea L. var. italica) root explants. Plant Cell Tiss. Organ Cult. 2010, 100, 49–58. [Google Scholar] [CrossRef]
- Banjac, N.; Vinterhalter, B.; Krstić-Milošević, D.; Milojević, J.; Tubić, L.; Ghalawenji, N.; Zdravković-Korać, S. Somatic embryogenesis and shoot organogenesis from the hypocotyl slices and free radical scavenging activity of regenerants of collard greens (Brassica oleracea L. var. acephala). Plant Cell Tiss. Organ Cult. 2019, 137, 613–626. [Google Scholar] [CrossRef]
- Al-Hardan, G.M.; AL-Shamari, M.; Hanshil, M.A. Impact of culture medium and 2,4-D on direct somatic embryogenesis in red cabbage (Brassica oleracea var. capitata forma rubra). Plant Arch. 2020, 20, 3022–3026. Available online: http://www.plantarchives.org/SPL%20ISSUE%2020-2/501__3022-3026_.pdf (accessed on 25 April 2023).
- Pavlović, S.; Savić, J.; Milojević, J.; Vinterhalter, B.; Girek, Z.; Adžić, S.; Zečević, B.; Banjac, N. Introduction of the Nicotiana protein kinase (NPK1) gene by combining Agrobacterium-mediated transformation and recurrent somatic embryogenesis to enhance salt tolerance in cauliflower. Plant Cell Tiss. Organ Cult. 2020, 143, 635–651. [Google Scholar] [CrossRef]
- Choi, P.S.; Soh, W.Y.; Liu, J.R. Somatic embryogenesis and plant regeneration in cotyledonary explant cultures of Chinese cabbage. Plant Cell Tiss. Organ Cult. 1996, 44, 253–256. [Google Scholar] [CrossRef]
- Barro, F.; Martín, A. Response of different genotypes of Brassica carinata to microspore culture. Plant Breed. 1999, 118, 79–81. [Google Scholar] [CrossRef]
- Shah, R.U.; Munir, I. Efficient in vitro plant regeneration through somatic embryogenesis from callus induction method for Brassica carinata. Sarhad J. Agric. 2019, 35, 314–319. Available online: https://researcherslinks.com/current-issues/Efficient-in-Vitro-Plant-Regeneration-Through-Somatic-Embryogenesis-from-Callus-Induction-Method-for-Brassica-carinata/14/1/2076 (accessed on 25 April 2023). [CrossRef]
- Piovan, A.; Caniato, R.; Filippini, R. Somatic embryogenesis and glucosinolate/myrosinase system in vulnerable Brassica repanda subsp. glabrescens (Poldini) Gómez-Campo. Sci. Hortic. 2014, 172, 317–324. [Google Scholar] [CrossRef]
- Faisal, M.; Abdel-Salam, E.M.; Alatar, A.A.; Qahtan, A.A. Induction of somatic embryogenesis in Brassica juncea L. and analysis of regenerants using ISSR-PCR and flow cytometer. Saudi J. Biol. Sci. 2021, 28, 1147–1153. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Singh, M.B. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat. Protoc. 2008, 3, 181–189. [Google Scholar] [CrossRef]
- Ghnaya, A.B.; Charles, G.; Branchard, M. Rapid shoot regeneration from thin cell layer explants excised from petioles and hypocotyls in four cultivars of Brassica napus L. Plant Cell Tiss. Organ Cult. 2008, 92, 25–30. [Google Scholar] [CrossRef]
- Pavlović, S.; Vinterhalter, B.; Mitić, N.; Adžić, S.; Pavlović, N.; Zdravković, M.; Vinterhalter, D. In vitro shoot regeneration from seedling explants in Brassica vegetables: Red cabbage, broccoli, Savoy cabbage and cauliflower. Arch. Biol. Sci. 2010, 62, 337–345. [Google Scholar] [CrossRef]
- Glendening, T.M.; Sjolund, R. In vitro propagation of kohlrabi from leaf explants. HortScience 1988, 23, 772. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.; Guo, B.; Bokhari, S.A.; Khan, M.A. Efficient regeneration and antioxidative enzyme activities in Brassica rapa var. turnip. Plant Cell Tiss. Organ Cult. 2011, 105, 337–344. [Google Scholar] [CrossRef]
- Lillo, C.; Shahin, E.A. Rapid regeneration of plants from hypocotyl protoplasts and root segments of cabbage. HortScience 1986, 21, 315–317. [Google Scholar] [CrossRef]
- Cogbill, S.; Faulcon, T.; Jones, G.; McDaniel, M.; Harmon, G.; Blackmon, R.; Young, M. Adventitious shoot regeneration from cotyledonary explants of rapid-cycling fast plants of Brassica rapa L. Plant Cell Tiss. Organ Cult. 2010, 101, 127–133. [Google Scholar] [CrossRef]
- Sharma, S.; Gambhir, G.; Srivastava, D.K. High Frequency Organogenesis in Cotyledon and Hypocotyl Explants of Cabbage (Brassica oleracea L. var. capitata). Natl. Acad. Sci. Lett. 2014, 37, 5–12. [Google Scholar] [CrossRef]
- Hu, Q.; Anderson, S.B.; Hansen, L.N. Plant regeneration capacity of mesophyll protoplasts from Brassica napus and related species. Plant Cell Tiss. Organ Cult. 1999, 59, 189–196. [Google Scholar] [CrossRef]
- Kumar, P.; Srivastava, D.K. High frequency organogenesis in hypocotyl, cotyledon, leaf and petiole explants of broccoli (Brassica oleracea L. var. italica), an important vegetable crop. Physiol. Mol. Biol. Plants 2015, 21, 279–285. [Google Scholar] [CrossRef]
- Bhalla, P.L.; de Weerd, N. In vitro propagation of cauliflower, Brassica oleracea var. botrytis for hybrid seed production. Plant Cell Tiss. Organ Cult. 1999, 56, 89–95. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Plant. 1996, 98, 291–297. [Google Scholar] [CrossRef]
- Klíma, M.; Vyvadilová, M.; Kučera, V. Production and utilization of doubled haploids in Brassica oleracea vegetables. Hortic. Sci. 2004, 31, 119–123. [Google Scholar] [CrossRef]
- Ćosić, T.; Vinterhalter, B.; Vinterhalter, D.; Mitić, N.; Cingel, A.; Savić, J.; Bohanec, B.; Ninković, S. In vitro plant regeneration from immature zygotic embryos and repetitive somatic embryogenesis in kohlrabi (Brassica oleracea var. gongylodes). Vitr. Cell. Dev. Biol. Plant 2013, 49, 294–303. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Raspor, M.; Savić, J.; Cingel, A.; Vinterhalter, B.; Vinterhalter, D.; Trávníčková, A.; Dobrev, P.I.; Bohanec, B.; et al. In vitro shoot organogenesis and comparative analysis of endogenous phytohormones in kohlrabi (Brassica oleracea var. gongylodes): Effects of genotype, explant type and applied cytokinins. Plant Cell Tiss. Organ Cult. 2015, 121, 741–760. [Google Scholar] [CrossRef]
- Ćosić, T.; Raspor, M.; Savić, J.; Cingel, A.; Matekalo, D.; Zdravković-Korać, S.; Ninković, S. Expression profiles of organogenesis-related genes over the time course of one-step de novo shoot organogenesis from intact seedlings of kohlrabi. J. Plant Physiol. 2019, 232, 257–269. [Google Scholar] [CrossRef]
- Ćosić, T.; Savić, J.; Raspor, M.; Cingel, A.; Ghalawnji, N.; Vinterhalter, B.; Ninković, S. Effects of different types of sugars and plant growth regulators on kohlrabi seedling growth and development in vitro. Arch. Biol. Sci. 2020, 72, 349–357. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Savić, J.; Raspor, M.; Marković, M.; Dobrev, P.I.; Ninković, S. Sucrose interferes with endogenous cytokinin homeostasis and expression of organogenesis-related genes during de novo shoot organogenesis in kohlrabi. Sci. Rep. 2021, 11, 6494. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Raspor, M.; Sajid, S.; Devrnja, N.; Dobrev, P.I.; Ninković, S. Comprehensive Phytohormone Profiling of Kohlrabi during in vitro Growth and Regeneration: The Interplay with Cytokinin and Sucrose. Life 2022, 12, 1585. [Google Scholar] [CrossRef]
- Gaj, M.D. Direct somatic embryogenesis as a rapid and efficient system for in vitro regeneration of Arabidopsis thaliana. Plant Cell Tiss. Organ Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and molecular control of somatic embryogenesis. Plants 2021, 10, 1467. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Stasolla, C. Transduction of Signals during Somatic Embryogenesis. Plants 2022, 11, 178. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Garin, E.; Isabel, N.; Plourde, A. Screening of large numbers of seed families of Pinus strobus L. for somatic embryogenesis from immature and mature zygotic embryos. Plant Cell Rep. 1998, 18, 37–43. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Park, J.C.; Soh, W.Y.; Choi, K.T. Regenerative ability of somatic single and multiple embryos from cotyledons of Korean ginseng on hormone-free medium. Plant Cell Rep. 1998, 17, 544–551. [Google Scholar] [CrossRef]
- Pillon, E.; Terzi, M.; Baldan, B.; Mariani, P.; Lo Schiavo, F. A protocol for obtaining embryogenic cell lines from Arabidopsis. Plant J. 1996, 9, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Mordhorst, A.P.; Voerman, K.J.; Hartog, M.V.; Meijer, E.A.; van Went, J.; Koornneef, M.; De Vries, S.C. Somatic embryogenesis in Arabidopsis thaliana is facilitated by mutations in genes repressing meristematic cell divisions. Genetics 1998, 149, 549–563. [Google Scholar] [CrossRef]
- Raemakers, C.J.J.M.; Jacobsen, E.; Visser, R.G.F. Secondary somatic embryogenesis and applications in plant breeding. Euphytica 1995, 81, 93–107. [Google Scholar] [CrossRef]
- von Arnold, S.; Clapham, D.; Egertsdotter, U.; Mo, L.H. Somatic embryogenesis in conifers—A case study of induction and development of somatic embryos in Picea abies. Plant Growth Regul. 1996, 20, 3–9. [Google Scholar] [CrossRef]
- Burbulis, N.; Kupriene, R. Induction of somatic embryos on in vitro cultured zygotic embryos of spring Brassica napus. Acta Univ. Latv. Biol. 2005, 691, 137–143. Available online: https://eeb.lu.lv/EEB/2005/Burbulis2.shtml (accessed on 21 April 2023).
- Rai, M.K.; Akhtar, N.; Jaiswal, V.S. Somatic embryogenesis and plant regeneration in Psidium guajava L. cv. Banarasi local. Sci. Hortic. 2007, 113, 129–133. [Google Scholar] [CrossRef]
- Kim, S.W.; Oh, S.C.; Liu, J.R. Control of direct and indirect somatic embryogenesis by exogenous growth regulators in immature zygotic embryo cultures of rose. Plant Cell Tiss. Organ Cult. 2003, 74, 61–66. [Google Scholar] [CrossRef]
- Konar, R.N.; Thomas, E.; Street, H.E. Origin and structure of embryoids arising from epidermal cells of the stem of Ranunculus sceleratus L. J. Cell Sci. 1972, 11, 77–93. [Google Scholar] [CrossRef]
- Sugimoto, K.; Jiao, Y.; Meyerowitz, E.M. Arabidopsis regeneration from multiple tissues occurs via a root development pathway. Dev. Cell 2010, 18, 463–471. [Google Scholar] [CrossRef]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tiss. Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-la-Peña, C.; Loyola-Vargas, V.M. Signaling Overview of Plant Somatic Embryogenesis. Front. Plant Sci. 2019, 10, 77. [Google Scholar] [CrossRef]
- Simonović, A.D.; Trifunović-Momčilov, M.M.; Filipović, B.K.; Marković, M.P.; Bogdanović, M.D.; Subotić, A.R. Somatic Embryogenesis in Centaurium erythraea Rafn—Current Status and Perspectives: A Review. Plants 2021, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.S.; Min, S.R.; Ahn, M.Y.; Soh, W.Y.; Liu, J.R. Somatic embryogenesis and plant regeneration in immature zygotic embryo, ovule, and anther filament cultures of Chinese cabbage. Sci. Hortic. 1998, 72, 151–155. [Google Scholar] [CrossRef]
- Rodriguez, A.P.M.; Wetzstein, H.Y. The effect of auxin type and concentration on pecan (Carya illinoinensis) somatic embryo morphology and subsequent conversion into plants. Plant Cell Rep. 1994, 13, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.P.M.; Wetzstein, H.Y. A morphological and histological comparison of the initiation and development of pecan (Carya illinoinensis) somatic embryogenic cultures induced with naphthaleneacetic acid or 2,4-dichlorophenoxyacetic acid. Protoplasma 1998, 204, 71–83. [Google Scholar] [CrossRef]
- Choi, Y.E.; Kim, H.S.; Soh, W.Y.; Yang, D.C. Developmental and structural aspects of somatic embryos formed on medium containing 2,3,5-triiodobenzoic acid. Plant Cell Rep. 1997, 16, 738–744. [Google Scholar] [CrossRef] [PubMed]
- von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant Cell Tiss. Organ Cult. 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Jiménez, V.M. Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul. 2005, 47, 91–110. [Google Scholar] [CrossRef]
- Ninković, S.; Miljuš-Djukić, J.; Nešković, M. Genetic transformation of alfalfa somatic embryos and their clonal propagation through repetitive somatic embryogenesis. Plant Cell Tiss. Organ Cult. 1995, 42, 255–260. [Google Scholar] [CrossRef]
- Uzelac, B.; Ninković, S.; Smigocki, A.; Budimir, S. Origin and development of secondary somatic embryos in transformed embryogenic cultures of Medicago sativa. Biol. Plant. 2007, 51, 1–6. [Google Scholar] [CrossRef]
- Bogdanović, M.D.; Ćuković, K.B.; Subotić, A.R.; Dragićević, M.B.; Simonović, A.D.; Filipović, B.K.; Todorović, S.I. Secondary Somatic Embryogenesis in Centaurium erythraea Rafn. Plants 2021, 10, 199. [Google Scholar] [CrossRef]
- Agarwal, S.; Kanwar, K.; Sharma, D.R. Factors affecting secondary somatic embryogenesis and embryo maturation in Morus alba L. Sci. Hortic. 2004, 102, 359–368. [Google Scholar] [CrossRef]
- Pavlović, S.; Vinterhalter, B.; Zdravković-Korać, S.; Vinterhalter, D.; Zdravković, J.; Cvikić, D.; Mitić, N. Recurrent somatic embryogenesis and plant regeneration from immature zygotic embryos of cabbage (Brassica oleracea var. capitata) and cauliflower (Brassica oleracea var. botrytis). Plant Cell Tiss. Organ Cult. 2013, 113, 397–406. [Google Scholar] [CrossRef]
- Vasic, D.; Alibert, G.; Skoric, D. Protocols for efficient repetitive and secondary somatic embryogenesis in Helianthus maximiliani (Schrader). Plant Cell Rep. 2001, 20, 121–125. [Google Scholar] [CrossRef]
- Thorpe, T.A. Somatic embryogenesis: Morphogenesis, physiology, biochemistry and molecular biology. Kor. J. Plant Tiss. Cult. 2000, 27, 245–258. [Google Scholar]
- Martinelli, L.; Candioli, E.; Costa, D.; Poletti, V.; Rascio, N. Morphogenic competence of Vitis rupestris S. secondary somatic embryos with a long culture history. Plant Cell Rep. 2001, 20, 279–284. [Google Scholar] [CrossRef]
- Haccius, B. Question of unicellular origin of non-zigotic embryos in callus cultures. Phytomorphology 1978, 28, 74–81. [Google Scholar]
- Toonen, M.A.J.; Hendriks, T.; Schmidt, E.D.L.; Verhoeven, H.A.; van Kammen, A.; de Vries, S.C. Description of somatic-embryo-forming single cells in carrot suspension cultures employing video cell tracking. Planta 1994, 194, 565–572. [Google Scholar] [CrossRef]
- Chan, J.L.; Saénz, L.; Talavera, C.; Hornung, R.; Robert, M.; Oropeza, C. Regeneration of coconut (Cocos nucifera L.) from plumule explants through somatic embryogenesis. Plant Cell Rep. 1998, 17, 515–521. [Google Scholar] [CrossRef]
- Kim, Y.W.; Youn, Y.; Noh, E.R.; Kim, J.C. Somatic embryogenesis and plant regeneration from immature embryos of five families of Quercus acutissima. Plant Cell Rep. 1997, 16, 869–873. [Google Scholar] [CrossRef]
- Yang, D.C.; Choi, Y.E. Production of transgenic plants via Agrobacterium rhizogenes-mediated transformation of Panax ginseng. Plant Cell Rep. 2000, 19, 491–496. [Google Scholar] [CrossRef]
- Ahmed, R.; Gupta, D.S.; De, D.N. Somatic embryogenesis and plant regeneration from leaf derived callus of winged bean [Psophocarpus tetragonolobus (L.) DC.]. Plant Cell Rep. 1996, 15, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Sarasan, V.; Roberts, A.V.; Rout, G.R. Methyl laurate and 6-benzyladenine promote the germination of somatic embryos of a hybrid rose. Plant Cell Rep. 2001, 20, 183–186. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Christianson, M.L.; Warnick, D.A. Temporal requirement for phytohormone balance in the control of organogenesis in vitro. Dev. Biol. 1985, 112, 494–497. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Su, Y.H.; Cheng, Z.J.; Zhang, X.S. Cell fate switch during in vitro plant organogenesis. J. Integr. Plant Biol. 2008, 50, 816–824. [Google Scholar] [CrossRef]
- Oinam, G.; Yeung, E.; Kurepin, L.; Haslam, T.; Lopez-Villalobos, A. Adventitious root formation in ornamental plants: I. General overview and recent successes. Propag. Ornam. Plants 2011, 11, 78–90. Available online: http://www.journal-pop.org/2011_11_2_78-90.html (accessed on 25 April 2023).
- Cary, A.J.; Che, P.; Howell, S.H. Developmental events and shoot apical meristem gene expression patterns during shoot development in Arabidopsis thaliana. Plant J. 2002, 32, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Gingerich, D.J.; Lall, S.; Howell, S.H. Global and hormone-induced gene expression changes during shoot development in Arabidopsis. Plant Cell 2002, 14, 2771–2785. [Google Scholar] [CrossRef]
- Valvekens, D.; Van Montagu, M.; Van Lijsebettens, M. Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. Sci. USA 1988, 85, 5536–5540. [Google Scholar] [CrossRef]
- Ćosić, T.; Raspor, M. The role of auxin and cytokinin signaling components in de novo shoot organogenesis. In Auxins, Cytokinins and Gibberellins Signaling in Plants; Aftab, T., Ed.; Springer: Cham, Switzerland, 2022; pp. 47–75. ISBN 978-3-031-05427-3. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Bueno, N.; Cuesta, C.; Feito, I.; Ordás, R.J. Hormonal and gene dynamics in de novo shoot meristem formation during adventitious caulogenesis in cotyledons of Pinus pinea. Plant Cell Rep. 2020, 39, 527–541. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, J.; Jie, E.Y.; Choi, S.H.; Jiang, L.; Ahn, W.S.; Kim, C.Y.; Kim, S.W. Temporal and spatial expression analysis of shoot-regeneration regulatory genes during the adventitious shoot formation in hypocotyl and cotyledon explants of tomato (cv. Micro-Tom). Int. J. Mol. Sci. 2020, 21, 5309. [Google Scholar] [CrossRef]
- Subrahmanyeswari, T.; Gantait, S.; Sarkar, S.; Bhattacharyya, S. Accelerated mono-phasic in vitro mass production of banana propagules and their morpho-cyto-genetic stability assessment. South Afr. J. Bot. 2022, 146, 794–806. [Google Scholar] [CrossRef]
- Sharma, K.K.; Thorpe, T.A. In vitro regeneration of shoot buds and plantlets from seedling root segments of Brassica napus L. Plant Cell Tiss. Organ Cult. 1989, 18, 129–141. [Google Scholar] [CrossRef]
- Mollika, S.R.; Sarker, R.H.; Hoque, M.I. In vitro plant regeneration in Brassica spp. Plant Tissue Cult. Biotech. 2011, 21, 127–134. [Google Scholar] [CrossRef]
- Ravanfar, S.A.; Aziz, M.A.; Kadir, M.A.; Rashid, A.A.; Haddadi, F. In vitro adventitious shoot regeneration and acclimatisation of Brassica oleracea subsp. italica cv. Green Marvel. Afr. J. Biotechnol. 2011, 10, 5614–5619. Available online: https://www.ajol.info/index.php/ajb/article/view/94349 (accessed on 30 March 2023).
- Ravanfar, S.A.; Aziz, M.A.; Rashid, A.A.; Salim, S. In vitro adventitious shoot regeneration from cotyledon explant of Brassica oleracea subsp. italica and Brassica oleracea subsp. capitata using TDZ and NAA. Pak. J. Bot. 2014, 46, 329–335. Available online: http://www.pakbs.org/pjbot/archives2.php?vol=46&iss=1&yea=2014 (accessed on 25 April 2023).
- Burnett, L.; Arnoldo, M.; Yarrow, S.; Huang, B. Enhancement of shoot regeneration from cotyledon explants of Brassica rapa ssp. oleifera through pretreatment with auxin and cytokinin and use of ethylene inhibitors. Plant Cell Tiss. Organ Cult. 1994, 37, 253–256. [Google Scholar] [CrossRef]
- Teo, W.; Lakshmanan, P.; Kumar, P.; Goh, C.J.; Swarup, S. Direct shoot formation and plant regeneration from cotyledon explants of rapid-cycling Brassica rapa. Vitr. Cell. Dev. Biol. Plant 1997, 33, 288–292. [Google Scholar] [CrossRef]
- Ono, Y.; Takahata, Y.; Kaizuma, N. Effect of genotype on shoot regeneration from cotyledonary explants of rapeseed (Brassica napus L.). Plant Cell Rep. 1994, 14, 13–17. [Google Scholar] [CrossRef]
- Sharma, K.K.; Bhojwani, S.S.; Thorpe, T.A. Factors affecting high frequency differentiation of shoots and roots from cotyledon explants of Brassica juncea (L.) Czern. Plant Sci. 1990, 66, 247–253. [Google Scholar] [CrossRef]
- Fuller, M.P.; Strullu, D.G.; Schlesser, A. The regeneration of shoots from seedlings explants of cauliflower. Crucif. Newslett. 1994, 16, 53–54. Available online: https://www.brassica.info/info/publications/cruciferae-newsletter.php (accessed on 25 April 2023).
- Kamal, G.B.; Illich, K.G.; Asadollah, A. Effects of genotype, explant type and nutrient medium components on canola (Brassica napus L.) shoot in vitro organogenesis. Afr. J. Biotechnol. 2007, 6, 861–867. Available online: https://www.ajol.info/index.php/ajb/article/view/56918 (accessed on 21 April 2023).
- Jin, R.G.; Liu, Y.B.; Tabashnik, B.E.; Borthakur, D. Development of transgenic cabbage (Brassica oleracea var. capitata) for insect resistance by Agrobacterium tumefaciens-mediated transformation. Vitr. Cell. Dev. Biol. Plant 2000, 36, 231–237. [Google Scholar] [CrossRef]
- Khan, M.R.; Rashid, H.; Ansar, M.; Chaudry, Z. High frequency shoot regeneration and Agrobacterium-mediated DNA transfer in Canola (Brassica napus). Plant Cell Tiss. Organ Cult. 2003, 75, 223–231. [Google Scholar] [CrossRef]
- Bedir, H.; Ari, E.; Vural, G.E.; Seguí-Simarro, J.M. Effect of the genotype, explant source and culture medium in somatic embryogenesis and organogenesis in Vaccaria hispanica (Mill.) Rauschert. Plant Cell Tiss. Organ Cult. 2022, 150, 329–343. [Google Scholar] [CrossRef]
- Yang, H.; Yuan, H.; Du, C.; Liang, L.; Chen, M.; Zou, L. Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants. Plants 2022, 11, 2456. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.L.; Wei, Y.H. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar] [CrossRef]
- Motyka, V.; Kamínek, M. Characterization of cytokinin oxidase from tobacco and poplar callus cultures. In Physiology and Biochemistry of Cytokinins in Plants; Kamínek, M., Mok, D.W.S., Zažímalová, E., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1992; pp. 33–39. ISBN 978-9051030662. [Google Scholar]
- Motyka, V.; Kamínek, M. Cytokinin oxidase from auxin- and cytokinin-dependent callus cultures of tobacco (Nicotiana tabacum L.). J. Plant Growth Regul. 1994, 13, 1–9. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Končitíková, R.; Zatloukal, M.; Bazgier, V.; Berka, K.; Zalabák, D.; Briozzo, P.; Strnad, M.; Spíchal, L. Novel thidiazuron-derived inhibitors of cytokinin oxidase/dehydrogenase. Plant Mol. Biol. 2016, 92, 235–248. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszki, J.; Teixeira da Silva, J.A.; Bulley, S.M.; Hudák, I. The role of cytokinins in shoot organogenesis in apple. Plant Cell Tiss. Organ Cult. 2010, 101, 251–267. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Nurmansyah; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef]
- Coleman, G.D.; Ernst, S.G. In vitro shoot regeneration of Populus deltoides: Effect of cytokinin and genotype. Plant Cell Rep. 1989, 8, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.M.; Kraus, J.E.; Endres, L.; Mercier, H. Relationships between endogenous hormonal levels and axillary bud development of Ananas comosus nodal segments. Plant Physiol. Biochem. 2003, 41, 733–739. [Google Scholar] [CrossRef]
- Suri, S.S.; Saini, A.R.K.; Ramawat, K.G. High frequency regeneration and Agrobacterium tumefaciens-mediated transformation of broccoli (Brassica oleracea var. italica). Eur. J. Hortic. Sci. 2005, 70, 71–78. Available online: https://www.pubhort.org/ejhs/2005/29079.htm (accessed on 30 March 2023).
- Nikolić, R.; Mitić, N.; Miletić, R.; Nešković, M. Effects of cytokinins on in vitro seed germination and early seedling morphogenesis in Lotus corniculatus L. J. Plant Growth Regul. 2006, 25, 187–194. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [PubMed]
- Miyawaki, K.; Tarkowski, P.; Matsumoto-Kitano, M.; Kato, T.; Sato, S.; Tarkowska, D.; Tabata, S.; Sandberg, G.; Kakimoto, T. Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 16598–16603. [Google Scholar] [CrossRef]
- Schmitz, R.Y.; Skoog, F.; Playtis, A.J.; Leonard, N.J. Cytokinins: Synthesis and biological activity of geometric and position isomers of zeatin. Plant Physiol. 1972, 50, 702–705. [Google Scholar] [CrossRef]
- Kamínek, M.; Pačes, V.; Corse, J.; Challice, J.S. Effect of stereospecific hydroxylation of N6-(Δ2-isopentenyl)adenosine on cytokinin activity. Planta 1979, 145, 239–243. [Google Scholar] [CrossRef]
- Gajdošová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef]
- Kudo, T.; Makita, N.; Kojima, M.; Tokunaga, H.; Sakakibara, H. Cytokinin Activity of cis-Zeatin and Phenotypic Alterations Induced by Overexpression of Putative cis-Zeatin-O-glucosyltransferase in Rice. Plant Physiol. 2012, 160, 319–331. [Google Scholar] [CrossRef]
- Stirk, W.A.; Václavíková, K.; Novák, O.; Gajdošová, S.; Kotland, O.; Motyka, V.; Strnad, M.; van Staden, J. Involvement of cis-zeatin, dihydrozeatin, and aromatic cytokinins in germination and seedling establishment of maize, oats, and lucerne. J. Plant Growth Regul. 2012, 31, 392–405. [Google Scholar] [CrossRef]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Grosskinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef]
- Ružić, D.V.; Vujović, T.I. The effects of cytokinin types and their concentration on in vitro multiplication of sweet cherry cv. Lapins (Prunus avium L.). Hortic. Sci. 2008, 35, 12–21. [Google Scholar] [CrossRef]
- Moura, M.; Candeias, M.I.; Silva, L. In vitro propagation of Viburnum treleasei Gand., an Azorean endemic with high ornamental interest. HortScience 2009, 44, 1668–1671. [Google Scholar] [CrossRef]
- Yunus, M.F.; Aziz, M.A.; Kadir, M.A.; Rashid, A.A. In vitro propagation of Etlingera elatior (Jack) (torch ginger). Sci. Hortic. 2012, 135, 145–150. [Google Scholar] [CrossRef]
- Kulus, D. Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. Vitr. Cell. Dev. Biol. Plant 2020, 56, 447–457. [Google Scholar] [CrossRef]
- Dang, S.; Gao, R.; Zhang, Y.; Feng, Y. In vitro regeneration and its histological characteristics of Dioscorea nipponica Makino. Sci. Rep. 2022, 12, 18436. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmülling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Bot. 2015, 66, 1851–1863. [Google Scholar] [CrossRef]
- Trifunović-Momčilov, M.; Motyka, V.; Dragićević, I.Č.; Petrić, M.; Jevremović, S.; Malbeck, J.; Holík, J.; Dobrev, P.I.; Subotić, A. Endogenous phytohormones in spontaneously regenerated Centaurium erythraea Rafn. plants grown in vitro. J. Plant Growth Regul. 2016, 35, 543–552. [Google Scholar] [CrossRef]
- Raspor, M.; Motyka, V.; Ninković, S.; Dobrev, P.I.; Malbeck, J.; Ćosić, T.; Cingel, A.; Savić, J.; Tadić, V.; Dragićević, I.Č. Endogenous levels of cytokinins, indole-3-acetic acid and abscisic acid in in vitro grown potato: A contribution to potato hormonomics. Sci. Rep. 2020, 10, 3437. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Guo, X.; Wang, Y.; Xiong, Y.; Wang, J.; Hayashi, K.I.; Lei, J.; Zhang, L.; Jiao, Y. Feedback from lateral organs controls shoot apical meristem growth by modulating auxin transport. Dev. Cell 2018, 44, 204–216. [Google Scholar] [CrossRef]
- Scarpella, E.; Barkoulas, M.; Tsiantis, M. Control of leaf and vein development by auxin. Cold Spring Harb. Perspect. Biol. 2010, 2, a001511. [Google Scholar] [CrossRef]
- Byrne, M.E. Making leaves. Curr. Opin. Plant Biol. 2012, 15, 24–30. [Google Scholar] [CrossRef]
- Peer, W.A.; Cheng, Y.; Murphy, A.S. Evidence of oxidative attenuation of auxin signalling. J. Exp. Bot. 2013, 64, 2629–2639. [Google Scholar] [CrossRef]
- Novák, O.; Hényková, E.; Sairanen, I.; Kowalczyk, M.; Pospíšil, T.; Ljung, K. Tissue-specific profiling of the Arabidopsis thaliana auxin metabolome. Plant J. 2012, 72, 523–536. [Google Scholar] [CrossRef]
- Kasahara, H. Current aspects of auxin biosynthesis in plants. Biosci. Biotechnol. Biochem. 2016, 80, 34–42. [Google Scholar] [CrossRef]
- Nemoto, K.; Hara, M.; Suzuki, M.; Seki, H.; Muranaka, T.; Mano, Y. The NtAMI1 gene functions in cell division of tobacco BY-2 cells in the presence of indole-3-acetamide. FEBS Lett. 2009, 583, 487–492. [Google Scholar] [CrossRef]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.K. Regulation of abscisic acid biosynthesis. Plant Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N. Abscisic acid and abiotic stress signalling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Borrego, E.; Kolomiets, M.V. Jasmonate biosynthesis, perception and function in plant development and stress responses. In Lipid Metabolism; Baez, R.V., Ed.; IntechOpen: London, UK, 2013; pp. 393–442. ISBN 978-953-51-0944-0. [Google Scholar] [CrossRef]
- Cuesta, C.; Novák, O.; Ordás, R.J.; Fernández, B.; Strnad, M.; Doležal, K.; Rodríguez, A. Endogenous cytokinin profiles and their relationships to between-family differences during adventitious caulogenesis in Pinus pinea cotyledons. J. Plant Physiol. 2012, 169, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Kamínek, M.; Motyka, V.; Vaňková, R. Regulation of cytokinin content in plant cells. Physiol. Plant. 1997, 101, 689–700. [Google Scholar] [CrossRef]
- Klemš, M.; Slámová, Z.; Motyka, V.; Malbeck, J.; Trávníčková, A.; Macháčková, I.; Holík, J.; Procházka, S. Changes in cytokinin levels and metabolism in tobacco (Nicotiana tabacum L.) explants during in vitro shoot organogenesis induced by trans-zeatin and dihydrozeatin. Plant Growth Regul. 2011, 65, 427–437. [Google Scholar] [CrossRef]
- Montalbán, I.A.; Novák, O.; Rolčik, J.; Strnad, M.; Moncaleán, P. Endogenous cytokinin and auxin profiles during in vitro organogenesis from vegetative buds of Pinus radiata adult trees. Physiol. Plant. 2013, 148, 214–231. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Bairu, M.W.; Novák, O.; Plíhalová, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. How does exogenously applied cytokinin type affect growth and endogenous cytokinins in micropropagated Merwilla plumbea? Plant Cell Tiss. Organ Cult. 2014, 118, 245–256. [Google Scholar] [CrossRef]
- Bassil, N.V.; Mok, D.W.S.; Mok, M.C. Partial purification of a cis-trans-isomerase of zeatin from immature seed of Phaseolus vulgaris L. Plant Physiol. 1993, 102, 867–872. [Google Scholar] [CrossRef]
- Kasahara, H.; Takei, K.; Ueda, N.; Hishiyama, S.; Yamaya, T.; Kamiya, Y.; Yamaguchi, S.; Sakakibara, H. Distinct isoprenoid origins of cis- and trans-zeatin biosyntheses in Arabidopsis. J. Biol. Chem. 2004, 279, 14049–14054. [Google Scholar] [CrossRef]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Vergara, L.E.; Palme, J.; Marín-de la Rosa, N.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A.; et al. Extensive signal integration by the phytohormone protein network. Nature 2020, 583, 271–276. [Google Scholar] [CrossRef]
- Romanov, G.A.; Lomin, S.N.; Schmülling, T. Biochemical characteristics and ligand-binding properties of Arabidopsis cytokinin receptor AHK3 compared to CRE1/AHK4 as revealed by a direct binding assay. J. Exp. Bot. 2006, 57, 4051–4058. [Google Scholar] [CrossRef]
- Zhang, C.G.; Li, W.; Mao, Y.F.; Zhao, D.L.; Dong, W.; Guo, G.Q. Endogenous hormonal levels in Scutellaria baicalensis calli induced by thidiazuron. Russ. J. Plant Physiol. 2005, 52, 345–351. [Google Scholar] [CrossRef]
- Eklöf, S.; Ȧstot, C.; Blackwell, J.; Moritz, T.; Olsson, O.; Sandberg, G. Auxin-cytokinin interactions in wild-type and transgenic tobacco. Plant Cell Physiol. 1997, 38, 225–235. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Wang, L.; Sun, W.; Zhang, Y.; Zhou, C.; Su, Y.H.; Li, W.; Sun, T.T.; Zhao, X.Y.; Li, X.G.; et al. Pattern of auxin and cytokinin responses for shoot meristem induction results from the regulation of cytokinin biosynthesis by AUXIN RESPONSE FACTOR3. Plant Physiol. 2013, 161, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Lall, S.; Nettleton, D.; Howell, S.H. Gene expression programs during shoot, root, and callus development in Arabidopsis tissue culture. Plant Physiol. 2006, 141, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Lall, S.; Howell, S.H. Developmental steps in acquiring competence for shoot development in Arabidopsis tissue culture. Planta 2007, 226, 1183–1194. [Google Scholar] [CrossRef]
- Motte, H.; Vereecke, D.; Geelen, D.; Werbrouck, S. The molecular path to in vitro shoot regeneration. Biotechnol. Adv. 2014, 32, 107–121. [Google Scholar] [CrossRef]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef]
- Pernisová, M.; Klíma, P.; Horák, J.; Válková, M.; Malbeck, J.; Souček, P.; Reichman, P.; Hoyerová, K.; Dubová, J.; Friml, J.; et al. Cytokinins modulate auxin-induced organogenesis in plants via regulation of the auxin efflux. Proc. Natl. Acad. Sci. USA 2009, 106, 3609–3614. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Su, Y.H.; Zhang, C.L.; Wang, L.; Li, X.G.; Zhang, X.S. Differences in capacities of in vitro organ regeneration between two Arabidopsis ecotypes Wassilewskija and Columbia. Plant Cell Tiss. Organ Cult. 2013, 112, 65–74. [Google Scholar] [CrossRef]
- Kakani, A.; Li, G.; Peng, Z. Role of AUX1 in the control of organ identity during in vitro organogenesis and in mediating tissue specific auxin and cytokinin interaction in Arabidopsis. Planta 2009, 229, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Koike, I.; Taniguchi, K.; Shimomura, K.; Umehara, M. Dynamics of endogenous indole-3-acetic acid and cytokinins during adventitious shoot formation in ipecac. J. Plant Growth Regul. 2017, 36, 805–813. [Google Scholar] [CrossRef]
- Salvi, E.; Di Mambro, R.; Pacifici, E.; Dello Ioio, R.; Costantino, P.; Moubayidin, L.; Sabatini, S. SCARECROW and SHORTROOT control the auxin/cytokinin balance necessary for embryonic stem cell niche specification. Plant Signal. Behav. 2018, 13, e1507402. [Google Scholar]
- Shao, D.; Ma, Y.; Li, X.; Ga, S.; Ren, Y. The sequence structure and phylogenetic analysis by complete mitochondrial genome of kohlrabi (Brassica oleracea var. gongylodes L.). Mitochondrial DNA B 2021, 6, 2714–2716. [Google Scholar] [CrossRef]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J. 2009, 57, 626–644. [Google Scholar] [CrossRef]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct conversion of root primordium into shoot meristem relies on timing of stem cell niche development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Berckmans, B.; De Veylder, L. Transcriptional control of the cell cycle. Curr. Opin. Plant Biol. 2009, 12, 599–605. [Google Scholar] [CrossRef]
- Dong, Q.; Zhao, Y.; Jiang, H.; He, H.; Zhu, S.; Cheng, B.; Xiang, Y. Genome-wide identification and characterization of the cyclin gene family in Populus trichocarpa. Plant Cell Tiss. Organ Cult. 2011, 107, 55–67. [Google Scholar] [CrossRef]
- Dewitte, W.; Murray, J.A.H. The plant cell cycle. Annu. Rev. Plant Biol 2003, 54, 235–264. [Google Scholar] [CrossRef]
- Nieuwland, J.; Maughan, S.; Dewitte, W.; Scofield, S.; Sanz, L.; Murray, J.A.H. The D-type cyclin CYCD4;1 modulates lateral root density in Arabidopsis by affecting the basal meristem region. Proc. Natl. Acad. Sci. USA 2009, 106, 22528–22533. [Google Scholar] [CrossRef]
- Barrôco, R.M.; Van Poucke, K.; Bergervoet, J.H.W.; De Veylder, L.; Groot, S.P.C.; Inzé, D.; Engler, G. The role of the cell cycle machinery in resumption of postembryonic development. Plant Physiol. 2005, 137, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Montero-Cortés, M.; Rodríguez-Paredes, F.; Burgeff, C.; Pérez-Nuñez, T.; Córdova, I.; Oropeza, C.; Verdeil, J.L.; Saénz, L. Characterisation of a cyclin-dependent kinase (CDKA) gene expressed during somatic embryogenesis of coconut palm. Plant Cell Tiss. Organ Cult. 2010, 102, 251–258. [Google Scholar] [CrossRef]
- Andersen, S.U.; Buechel, S.; Zhao, Z.; Ljung, K.; Novák, O.; Busch, W.; Schuster, C.; Lohmann, J.U. Requirement of B2-type Cyclin-Dependent Kinases for meristem integrity in Arabidopsis thaliana. Plant Cell 2008, 20, 88–100. [Google Scholar] [CrossRef]
- Gutierrez, C. Coupling cell proliferation and development in plants. Nat. Cell Biol. 2005, 7, 535–541. [Google Scholar] [CrossRef]
- Planchais, S.; Glab, N.; Tréhin, C.; Perennes, C.; Bureau, J.M.; Meijer, L.; Bergounioux, C. Roscovitine, a novel cyclin-dependent kinase inhibitor, characterizes restriction point and G2/M transition in tobacco BY-2 cell suspension. Plant J. 1997, 12, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Redig, P.; Shaul, O.; Inzé, D.; Van Montagu, M.; Van Onckelen, H. Levels of endogenous cytokinins, indole-3-acetic acid and abscisic acid during the cell cycle of synchronized tobacco BY-2 cells. FEBS Lett. 1996, 391, 175–180. [Google Scholar] [CrossRef]
- Laureys, F.; Dewitte, W.; Witters, E.; Van Montagu, M.; Inzé, D.; Van Onckelen, H. Zeatin is indispensable for the G2-M transition in tobacco BY-2 cells. FEBS Lett. 1998, 426, 29–32. [Google Scholar] [CrossRef]
- Zhang, K.; Diederich, L.; John, P.C.L. The cytokinin requirement for cell division in cultured Nicotiana plumbaginifolia cells can be satisfied by yeast Cdc25 protein tyrosine phosphatase. Implications for mechanisms of cytokinin response and plant development. Plant Physiol. 2005, 137, 308–316. [Google Scholar] [CrossRef]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the cell cycle. Curr. Opin. Plant Biol. 2014, 21, 7–15. [Google Scholar] [CrossRef]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef]
- Mishra, B.S.; Sharma, M.; Laxmi, A. Role of sugar and auxin crosstalk in plant growth and development. Physiol. Plant. 2022, 174, e13546. [Google Scholar] [CrossRef]
- Hara, M.; Oki, K.; Hoshino, K.; Kuboi, T. Enhancement of anthocyanin biosynthesis by sugar in radish (Raphanus sativus) hypocotyl. Plant Sci. 2003, 164, 259–265. [Google Scholar] [CrossRef]
- Song, X.; Guo, H.; Liu, Y.; Wan, F.; Zhang, J.; Chang, X. Effects of salicylic acid and sucrose on pigment content in Pistacia chinensis leaves. Sci. Hortic. 2020, 259, 108783. [Google Scholar] [CrossRef]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2019, 48, 1–11. [Google Scholar] [CrossRef]
- LeClere, S.; Schmelz, E.A.; Chourey, P.S. Sugar levels regulate tryptophan-dependent auxin biosynthesis in developing maize kernels. Plant Physiol. 2010, 153, 306–318. [Google Scholar] [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed]
- Meitzel, T.; Radchuk, R.; McAdam, E.L.; Thormählen, I.; Feil, R.; Munz, E.; Hilo, A.; Geigenberger, P.; Ross, J.J.; Lunn, J.E.; et al. Trehalose 6-phosphate promotes seed filling by activating auxin biosynthesis. New Phytol. 2021, 229, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Sairanen, I.; Novák, O.; Pěnčík, A.; Ikeda, Y.; Jones, B.; Sandberg, G.; Ljung, K. Soluble Carbohydrates Regulate Auxin Biosynthesis via PIF Proteins in Arabidopsis. Plant Cell 2012, 24, 4907–4916. [Google Scholar] [CrossRef] [PubMed]
- McAdam, E.L.; Meitzel, T.; Quittenden, L.J.; Davidson, S.E.; Dalmais, M.; Bendahmane, A.I.; Thompson, R.; Smith, J.J.; Nichols, D.S.; Urquhart, S.; et al. Evidence that auxin is required for normal seed size and starch synthesis in pea. New Phytol. 2017, 216, 193–204. [Google Scholar] [CrossRef]
- Cheng, W.H.; Endo, A.; Zhou, L.; Penney, J.; Chen, H.C.; Arroyo, A.; Leon, P.; Nambara, E.; Asami, T.; Seo, M.; et al. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 2002, 14, 2723–2743. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Kamiya, Y. Gibberellin biosynthesis: Its regulation by endogenous and environmental signals. Plant Cell Physiol. 2000, 41, 251–257. [Google Scholar] [CrossRef]
- Gordon, S.P.; Heisler, M.G.; Reddy, G.V.; Ohno, C.; Das, P.; Meyerowitz, E.M. Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development 2007, 134, 3539–3548. [Google Scholar] [CrossRef]
- Huang, W.L.; Lee, C.H.; Chen, Y.R. Levels of endogenous abscisic acid and indole-3-acetic acid influence shoot organogenesis in callus cultures of rice subjected to osmotic stress. Plant Cell Tiss. Organ Cult. 2012, 108, 257–263. [Google Scholar] [CrossRef]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef]
- Kushwah, S.; Laxmi, A. The interaction between glucose and cytokinin signal transduction pathway in Arabidopsis thaliana. Plant Cell Environ. 2014, 37, 235–253. [Google Scholar] [CrossRef]
- Kiba, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H. Sugar-induced de novo cytokinin biosynthesis contributes to Arabidopsis growth under elevated CO2. Sci. Rep. 2019, 9, 7765. [Google Scholar] [CrossRef]
- Wang, M.; Le Gourrierec, J.; Jiao, F.; Demotes-Mainard, S.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Crespel, L.; Bertheloot, J.; Chen, J.; et al. Convergence and divergence of sugar and cytokinin signaling in plant development. Int. J. Mol. Sci. 2021, 22, 1282. [Google Scholar] [CrossRef]
- Barbier, F.; Péron, T.; Lecerf, M.; Perez-Garcia, M.D.; Barrière, Q.; Rolčík, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, S.; Laxmi, A. The interaction between glucose and cytokinin signaling in controlling Arabidopsis thaliana seedling root growth and development. Plant Signal. Behav. 2017, 12, e1312241. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).