

Expression Analysis of Trihelix Transcription Factor Family in Strawberries and Functional Characterization of FvTrihelix6
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Seedlings Treatment
2.2. Identification of Trihelix Genes in the F. vesca Genome
2.3. Phylogenetic Analysis, Gene Structure, Motif Analysis, and Multiple Sequence Alignment
2.4. Chromosomal Distribution and Promoter Analysis
2.5. RNA Extraction and PCR Analysis
2.6. Plasmid Construction and Transformation
2.7. Subcellular Localization of FvTrihelix6
2.8. Cloning of FvTrihelix6 Promoter, GUS Protein Staining, and GUS Activity Assay
2.9. Statistical Analyses
3. Results
3.1. Identification of Strawberry TTF Members
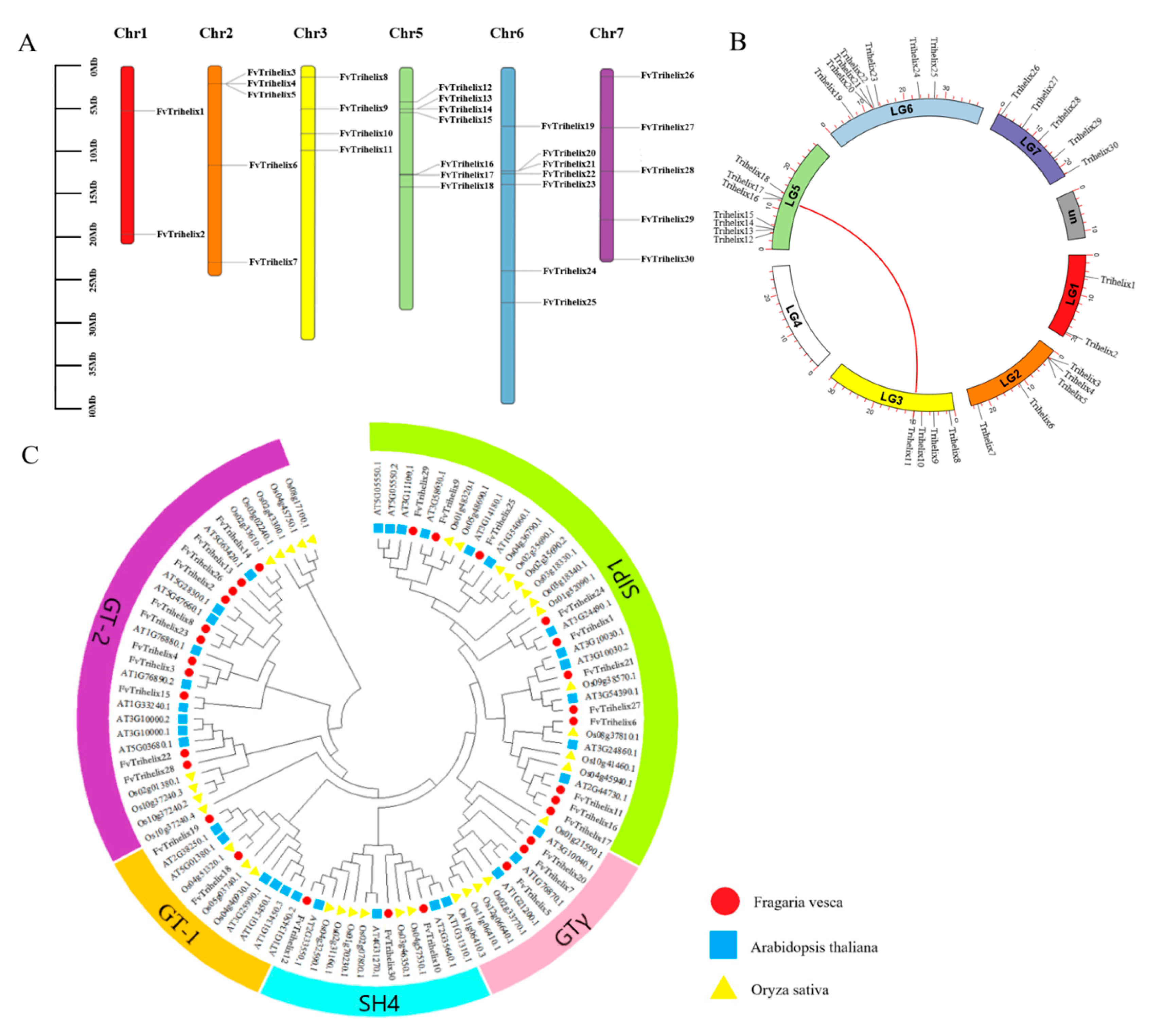
3.2. The Distribution of FvTrihelix Genes across Chromosomes and Evolutionary Relationships
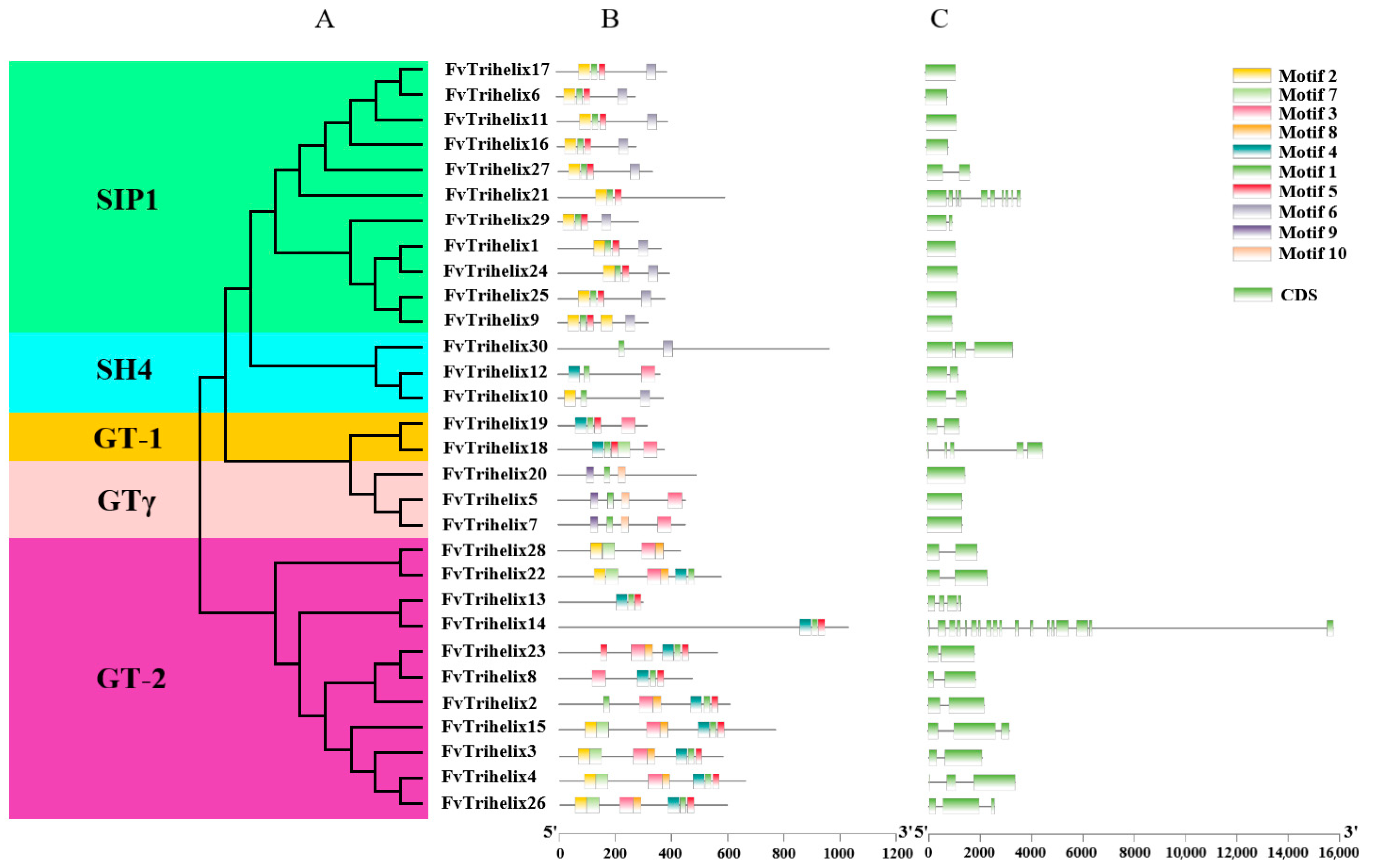
3.3. Phylogenetic and Gene Structure Analysis
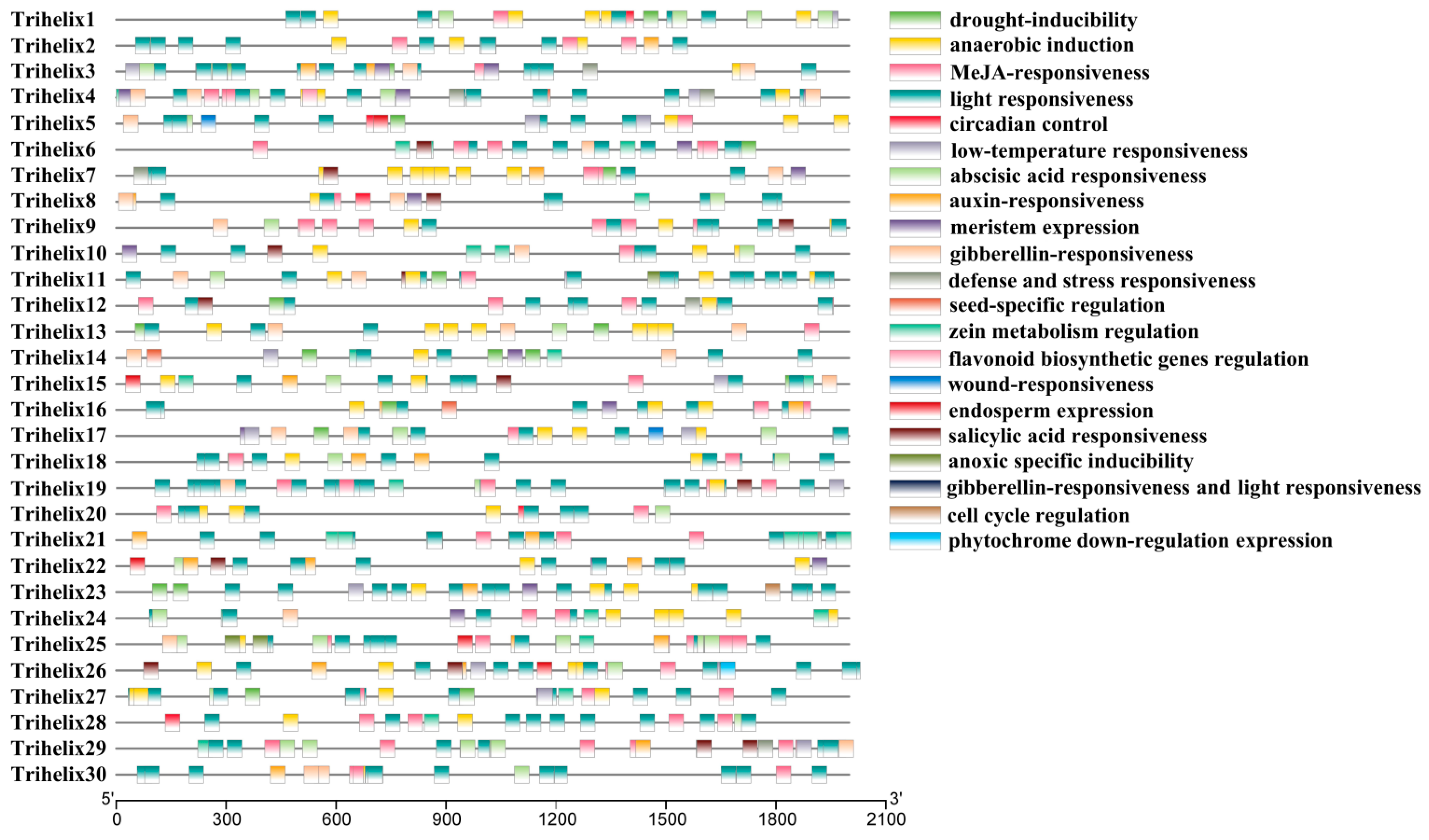
3.4. Cis-Acting Elements Analysis
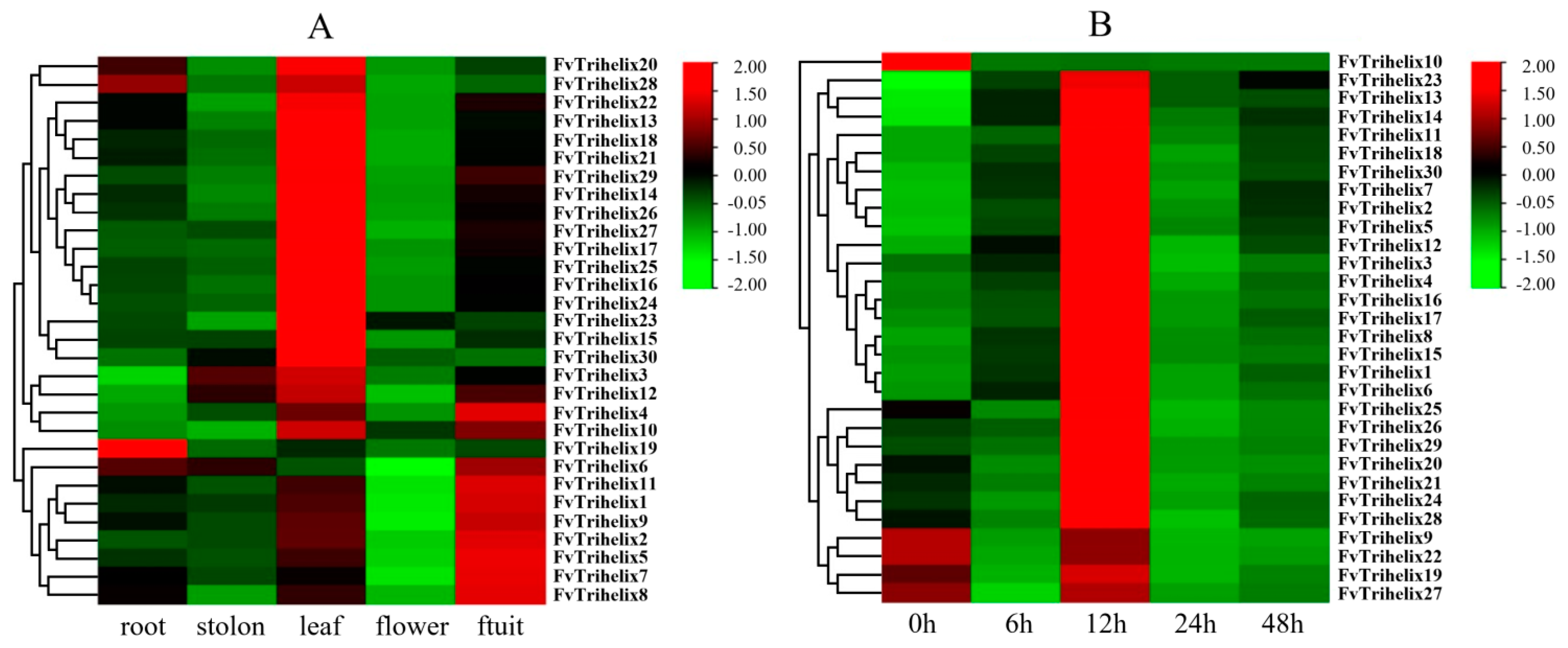
3.5. Expression Analysis of TTF Genes in Various Parts of the Strawberry
3.6. Evaluation of the FvTrihelix Genes’ Expression following C. gloeosporioides Inoculation
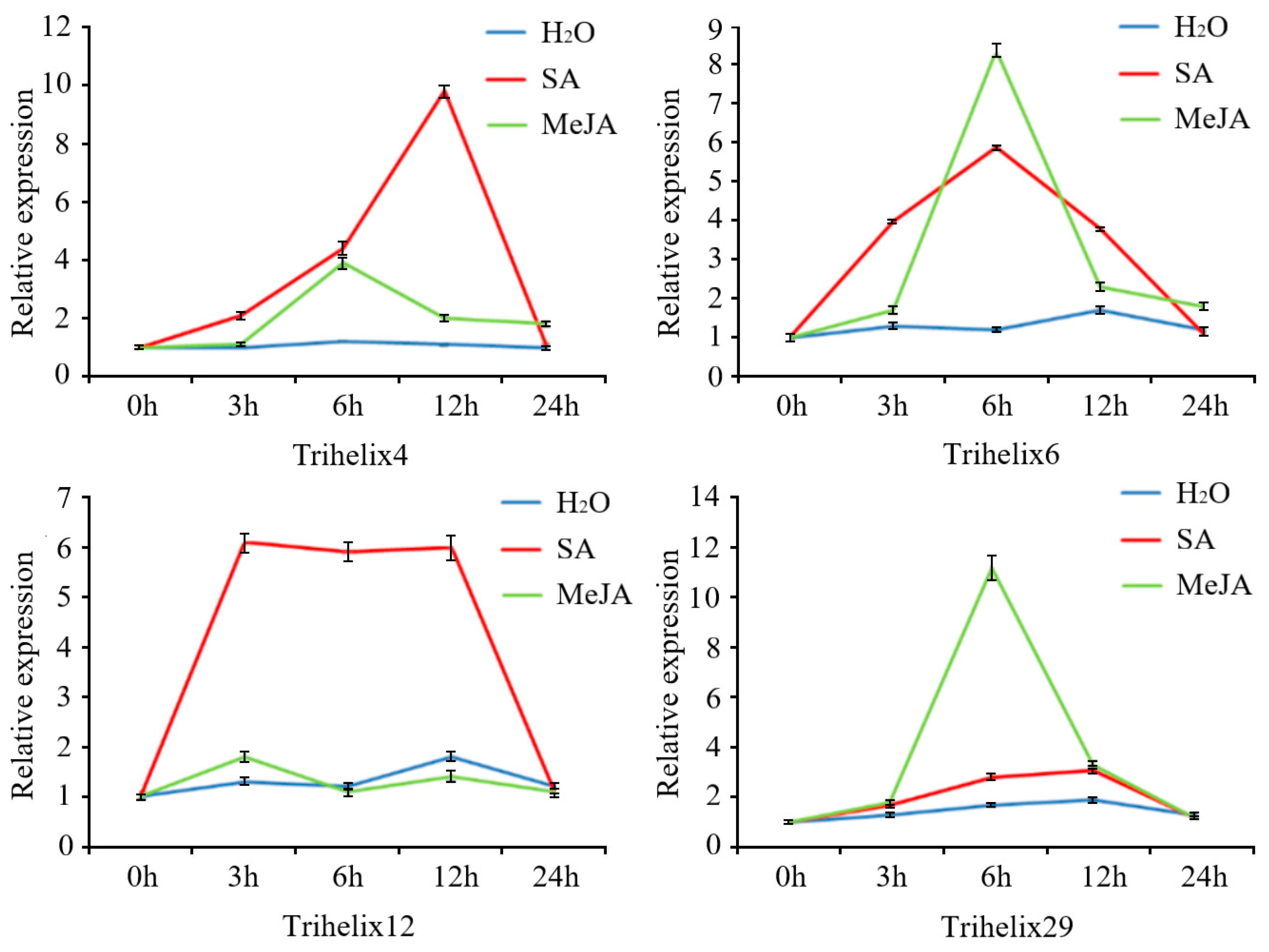
3.7. Expression Profiling of Selected Strawberry TTF Genes in Strawberry Leaves following SA and MeJA Treatments
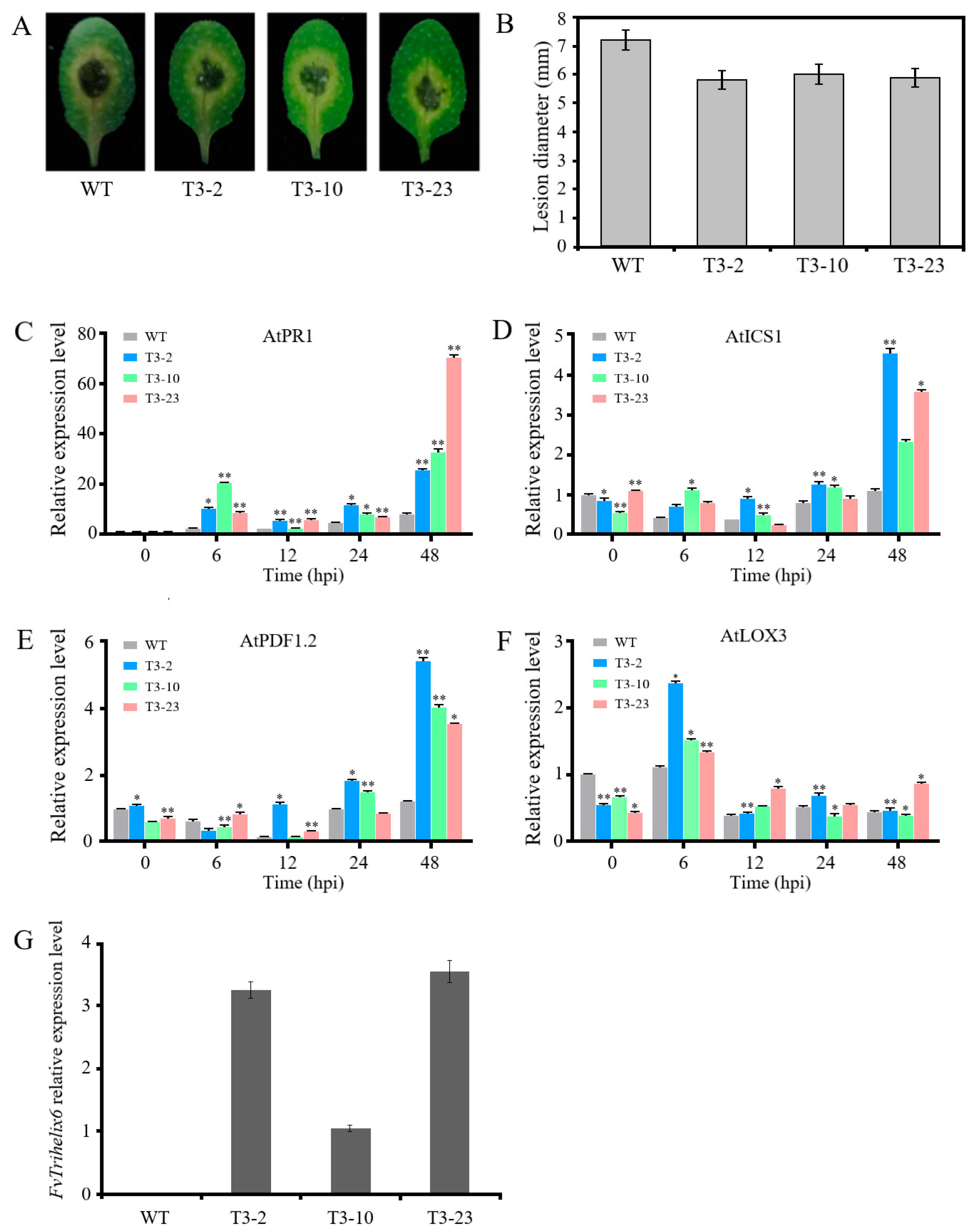
3.8. FvTrihelix6 Ectopic Expression in A. thaliana Enhanced Resistance to C. higginsianum
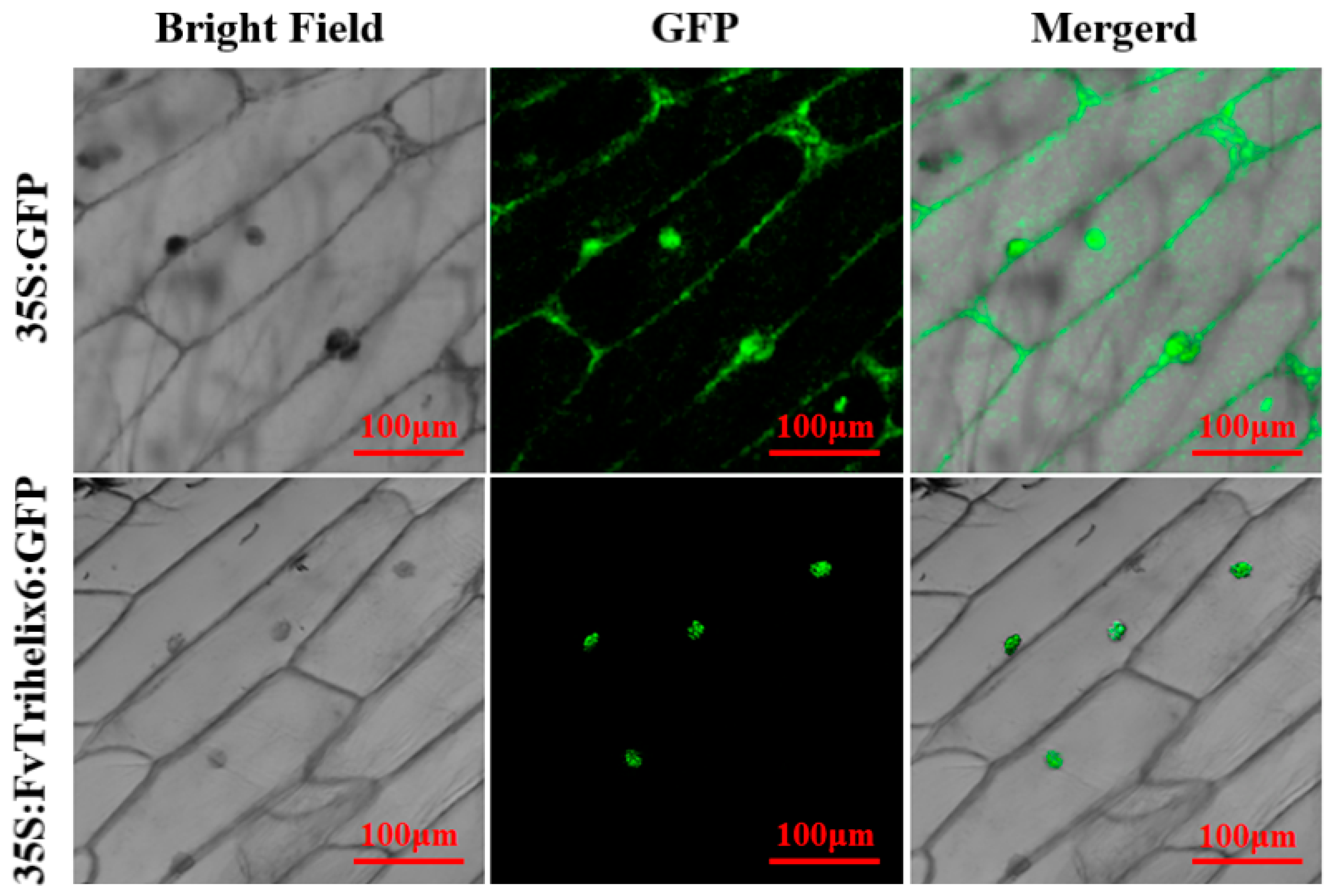
3.9. Subcellular Localization of FvTrihelix6
3.10. Response of the FvTrihelix6 Promoter to SA and C. gloeosporioides Using a GUS Reporter
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, P.J.; Kay, S.A.; Chua, N.H. Sequence-specific interactions of a pea nuclear factor with light-responsive elements upstream of the rbcS-3A gene. EMBO J. 1987, 6, 2543–2549. [Google Scholar] [CrossRef]
- Yu, C.; Cai, X.; Ye, Z.; Li, H. Genome-wide identification and expression profiling analysis of trihelix gene family in tomato. Biochem. Biophys. Res. Commun. 2015, 468, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Wu, D.; Fan, Q.; Tian, C.; Chen, S.; Guan, Z.; Xin, J.; Zhao, K.; Chen, F. Transcriptome-Wide Identification and Expression Profiling Analysis of Chrysanthemum Trihelix Transcription Factors. Int. J. Mol. Sci. 2016, 17, 198. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Q.; Wang, H.; Zhang, H.; Xu, X.; Li, C.; Yang, C. Comprehensive analysis of trihelix genes and their expression under biotic and abiotic stresses in Populus trichocarpa. Sci. Rep. 2016, 6, 36274. [Google Scholar] [CrossRef]
- Kaplan-Levy, R.N.; Brewer, P.B.; Quon, T.; Smyth, D.R. The trihelix family of transcription factors—Light, stress and development. Trends Plant Sci. 2012, 17, 163–171. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; et al. Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Li, W.; Lin, Y.; Wang, C.; Xu, R.; Zhang, L. Genome-wide characterization and expression analysis of soybean trihelix gene family. PeerJ 2020, 8, e8753. [Google Scholar] [CrossRef]
- Li, K.; Duan, L.; Zhang, Y.; Shi, M.; Chen, S.; Yang, M.; Ding, Y.; Peng, Y.; Dong, Y.; Yang, H.; et al. Genome-wide identification and expression profile analysis of trihelix transcription factor family genes in response to abiotic stress in sorghum [Sorghum bicolor (L.) Moench]. BMC Genom. 2021, 22, 738. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Bin, J.; Ding, H.; Pan, D.; Tian, Q.; Yang, X.; Wang, L.; Yue, Y. Insights into the trihelix transcription factor responses to salt and other stresses in Osmanthus fragrans. BMC Genom. 2022, 23, 334. [Google Scholar] [CrossRef]
- Li, K.; Fan, Y.; Zhou, G.; Liu, X.; Chen, S.; Chang, X.; Wu, W.; Duan, L.; Yao, M.; Wang, R.; et al. Genome-wide identification, phylogenetic analysis, and expression profiles of trihelix transcription factor family genes in quinoa (Chenopodium quinoa Willd.) under abiotic stress conditions. BMC Genom. 2022, 23, 499. [Google Scholar] [CrossRef]
- Wang, J.; Ouyang, Y.; Wei, Y.; Kou, J.; Zhang, X.; Zhang, H. Identification and Characterization of Trihelix Transcription Factors and Expression Changes during Flower Development in Pineapple. Horticulturae 2022, 8, 894. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Pan, Q.; Chen, S.; Feng, C.; Hai, J.; Li, H. Comparison of Trihelix transcription factors between wheat and Brachypodium distachyon at genome-wide. BMC Genom. 2019, 20, 142. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Hu, J.; Behera, J.R.; Kilaru, A.; Yuan, Y.; Zhai, Y.; Xu, Y.; Xie, L.; Zhang, Y.; Zhang, Q.; et al. A Tree Peony Trihelix Transcription Factor PrASIL1 Represses Seed Oil Accumulation. Front. Plant Sci. 2021, 12, 796181. [Google Scholar] [CrossRef]
- Feng, C.; Song, X.; Tang, H. Molecular cloning and expression analysis of GT-2-like genes in strawberry. 3 Biotech 2019, 9, 105. [Google Scholar] [CrossRef]
- Tzafrir, I.; Pena-Muralla, R.; Dickerman, A.; Berg, M.; Rogers, R.; Hutchens, S.; Sweeney, T.C.; McElver, J.; Aux, G.; Patton, D.; et al. Identification of genes required for embryo development in Arabidopsis. Plant Physiol. 2004, 135, 1206–1220. [Google Scholar] [CrossRef]
- Weng, H.; Yoo, C.Y.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. Poplar GTL1 is a Ca2+/calmodulin-binding transcription factor that functions in plant water use efficiency and drought tolerance. PLoS ONE 2012, 7, e32925. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.H.; Chen, F.; Torres-Jerez, I.; Tang, Y.; Wang, M.; Du, Q.; Cheng, X.; Wen, J.; Dixon, R. Transcriptome analysis of secondary cell wall development in Medicago truncatula. BMC Genom. 2016, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, D.; Shan, T.; Xu, S.; Qin, R.; Li, H.; Negm, M.; Wu, D.; Li, J. The trihelix transcription factor OsGTγ-2 is involved adaption to salt stress in rice. Plant Mol. Biol. 2020, 103, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Song, L.; Song, J.; Ouyang, B.; Guo, L.; Shang, L.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. ShCIGT, a Trihelix family gene, mediates cold and drought tolerance by interacting with SnRK1 in tomato. Plant Sci. 2018, 270, 140–149. [Google Scholar] [CrossRef]
- Li, Y.; Hu, Z.; Dong, Y.; Xie, Z. Trihelix Transcriptional Factor GhGT26 of Cotton Enhances Salinity Tolerance in Arabidopsis. Plants 2022, 11, 2694. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, M.; Zhang, Y.; Huang, M.; Wei, L.; Lin, Y.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D.; et al. Coordinated regulation of plant defense and autoimmunity by paired trihelix transcription factors ASR3/AITF1 in Arabidopsis. New Phytol. 2023, 237, 914–929. [Google Scholar] [CrossRef] [PubMed]
- Völz, R.; Kim, S.K.; Mi, J.; Mariappan, K.G.; Guo, X.; Bigeard, J.; Alejandro, S.; Pflieger, D.; Rayapuram, N.; Al-Babili, S.; et al. The Trihelix transcription factor GT2-like 1 (GTL1) promotes salicylic acid metabolism, and regulates bacterial-triggered immunity. PLoS Genet. 2018, 14, e1007708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhong, T.E.L.; Xu, M.; Dai, W.; Sun, S.; Ye, J. GT Factor ZmGT-3b Is Associated With Regulation of Photosynthesis and Defense Response to Fusarium graminearum Infection in Maize Seedling. Front. Plant Sci. 2021, 12, 724133. [Google Scholar] [CrossRef] [PubMed]
- Park, H.C.; Kim, M.L.; Kang, Y.H.; Jeon, J.M.; Yoo, J.H.; Kim, M.C.; Park, C.Y.; Jeong, J.C.; Moon, B.C.; Lee, J.H.; et al. Pathogen- and NaCl-induced expression of the SCaM-4 promoter is mediated in part by a GT-1 box that interacts with a GT-1-like transcription factor. Plant Physiol. 2004, 135, 2150–2161. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hong, G.; Han, B. Transcript abundance of rml1, encoding a putative GT1-like factor in rice, is up-regulated by Magnaporthe grisea and down-regulated by light. Gene 2004, 324, 105–115. [Google Scholar] [CrossRef]
- Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Strawberry as a health promoter: An evidence based review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef]
- Miller, K.; Feucht, W.; Schmid, M. Bioactive Compounds of Strawberry and Blueberry and Their Potential Health Effects Based on Human Intervention Studies: A Brief Overview. Nutrients 2019, 11, 1510. [Google Scholar] [CrossRef]
- Morkeliūnė, A.; Rasiukevičiūtė, N.; Valiuškaitė, A. Conditions in a Temperate Climate for Colletotrichum acutatum, Strawberry Pathogen Distribution and Susceptibility of Different Cultivars to Anthracnose. Agriculture 2021, 11, 80. [Google Scholar] [CrossRef]
- Chen, X.Y.; Dai, D.J.; Zhao, S.F.; Shen, Y.; Wang, H.D.; Zhang, C.Q. Genetic Diversity of Colletotrichum spp. Causing Strawberry Anthracnose in Zhejiang, China. Plant Dis. 2020, 104, 1351–1357. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Encinas-Villarejo, S.; Santos, B.d.l.; Schilirò, E.; Yubero-Serrano, E.-M.; Amil-Ruíz, F.; Pocovi, M.I.; Pliego-Alfaro, F.; Dorado, G.; Rey, M.; et al. Analysis of strawberry genes differentially expressed in response to Colletotrichum infection. Physiol. Plant. 2006, 128, 633–650. [Google Scholar] [CrossRef]
- Wen, Z.; Bai, J.; Wang, L.; Yao, L.; Ahmad, B.; Hanif, M.; Chen, Q. Over expression of a Chitinase 2 gene from Chinese Wild Strawberry improves resistance to anthracnose disease in transgenic Arabidopsis thaliana. Plant Biotechnol. Rep. 2020, 14, 725–736. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-mediated gene editing in Arabidopsis thaliana and inheritance of modified genes in the T2 and T3 generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Shen, Y.; Ahmad, B.; Yao, L.; He, T.; Fan, J.; Liu, Y.; Chen, Q.; Wen, Z. Genome-wide identification and expression analysis of dirigent gene family in strawberry (Fragaria vesca) and functional characterization of FvDIR13. Sci. Hortic. 2022, 297, 110913. [Google Scholar] [CrossRef]
- Kosugi, S.; Ohashi, Y.; Nakajima, K.; Arai, Y.J.P. An improved assay for ß-glucuronidase (GUS) in transformed cells: Methanol almost suppresses a putative endogenous GUS activity. Plant Sci. 1990, 70, 133–140. [Google Scholar] [CrossRef]
- Wang, L.; Huang, M.; Yao, L.; Bai, J.; Chen, Q.; Wen, F. Cloning and Expression Analysis of FnMYB24 Transcription Factor Gene and Promoter from Fragaria nilgerrensis Schltdl (Chinese). Acta Bot. Boreali-Occident. Sinica 2020, 40, 1646–1654. [Google Scholar]
- Le Gourrierec, J.; Li, Y.F.; Zhou, D.X. Transcriptional activation by Arabidopsis GT-1 may be through interaction with TFIIA-TBP-TATA complex. Plant J. 1999, 18, 663–668. [Google Scholar] [CrossRef]
- Ji, J.H.; Zhou, Y.J.; Wu, H.H.; Yang, L.M. Genome-wide analysis and functional prediction of the Trihelix transcription factor family in rice. Yi Chuan Hered. 2015, 37, 1228–1241. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Schmelzer, E.; Hahlbrock, K.; Somssich, I.E. Early nuclear events in plant defence signalling: Rapid gene activation by WRKY transcription factors. EMBO J. 1999, 18, 4689–4699. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Reinstädler, A.; Lipka, V.; Lippok, B.; Somssich, I.E. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling. Plant Cell 2002, 14, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shi, L.; Yang, S.; Lin, Y.; Weng, Y.; Li, X.; Hussain, A.; Noman, A.; He, S. Functional and Promoter Analysis of ChiIV3, a Chitinase of Pepper Plant, in Response to Phytophthora capsici Infection. Int. J. Mol. Sci. 2017, 18, 1661. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Garrido-Gala, J.; Gadea, J.; Blanco-Portales, R.; Muñoz-Mérida, A.; Trelles, O.; de Los Santos, B.; Arroyo, F.T.; Aguado-Puig, A.; Romero, F.; et al. Partial Activation of SA- and JA-Defensive Pathways in Strawberry upon Colletotrichum acutatum Interaction. Front. Plant Sci. 2016, 7, 1036. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Strawn, M.A.; Marr, S.K.; Inoue, K.; Inada, N.; Zubieta, C.; Wildermuth, M.C. Arabidopsis isochorismate synthase functional in pathogen-induced salicylate biosynthesis exhibits properties consistent with a role in diverse stress responses. J. Biol. Chem. 2007, 282, 5919–5933. [Google Scholar] [CrossRef]
- Uknes, S.; Mauch-Mani, B.; Moyer, M.; Potter, S.; Williams, S.; Dincher, S.; Chandler, D.; Slusarenko, A.; Ward, E.; Ryals, J. Acquired resistance in Arabidopsis. Plant Cell 1992, 4, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14 (Suppl. S1), S153–S164. [Google Scholar] [CrossRef]
- Dubouzet, J.G.; Maeda, S.; Sugano, S.; Ohtake, M.; Hayashi, N.; Ichikawa, T.; Kondou, Y.; Kuroda, H.; Horii, Y.; Matsui, M.; et al. Screening for resistance against Pseudomonas syringae in rice-FOX Arabidopsis lines identified a putative receptor-like cytoplasmic kinase gene that confers resistance to major bacterial and fungal pathogens in Arabidopsis and rice. Plant Biotechnol. J. 2011, 9, 466–485. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Accession No | Locus Name | Chr | Location | CDS (bp) | ORF (aa) | MW (kDa) | pI | Instability Index | Predicted Location |
|---|---|---|---|---|---|---|---|---|---|---|
| FvTrihelix1 | XM_004288645.2 | XP_004288693.1 | 1 | 458973..461535 | 1095 | 364 | 42.27 | 4.87 | 54.62 | Nucl |
| FvTrihelix2 | XM_004301984.2 | XP_004302032.1 | 1 | 346457..349017 | 1824 | 607 | 69.18 | 6.17 | 72.34 | Nucl |
| FvTrihelix3 | XM_004290471.2 | XP_004290519.1 | 2 | 14323195..14325618 | 1737 | 578 | 65.76 | 6.21 | 55.76 | Nucl |
| FvTrihelix4 | XM_004290472.2 | XP_004290520.1 | 2 | 14336737..14340021 | 1920 | 639 | 71.32 | 6.35 | 59.55 | Nucl |
| FvTrihelix5 | XM_011460207.1 | XP_011458509.1 | 2 | 14397063..14399118 | 1356 | 451 | 52.00 | 6.27 | 52.21 | Nucl |
| FvTrihelix6 | XM_004292325.2 | XP_004292373.1 | 2 | 9441933..9443903 | 840 | 279 | 32.40 | 5.91 | 71.36 | Nucl |
| FvTrihelix7 | XM_004291753.2 | XP_004291801.1 | 2 | 31810046..31812474 | 1356 | 451 | 51.28 | 6.08 | 48.94 | Nucl |
| FvTrihelix8 | XM_004293354.2 | XP_004293402.1 | 3 | 1308854..1311352 | 1419 | 472 | 52.26 | 6.15 | 52.56 | Nucl |
| FvTrihelix9 | XM_011462305.1 | XP_011460607.1 | 3 | 7431462..7435172 | 930 | 309 | 34.22 | 5.10 | 49.46 | Nucl |
| FvTrihelix10 | XM_011462154.1 | XP_011460456.1 | 3 | 5320278..5321873 | 1119 | 372 | 40.81 | 9.06 | 66.70 | Nucl |
| FvTrihelix11 | XM_004307199.2 | XP_004307247.1 | 3 | 13178449..13180706 | 1071 | 356 | 39.37 | 9.27 | 68.86 | Nucl |
| FvTrihelix12 | XM_004298478.2 | XP_004298526.1 | 5 | 19672557..19675420 | 1083 | 360 | 40.95 | 5.49 | 64.73 | Nucl |
| FvTrihelix13 | XM_011464980.1 | XP_011463282.1 | 5 | 20486591..20487951 | 725 | 239 | 27.29 | 9.07 | 64.54 | Nucl |
| FvTrihelix14 | XM_004297485.2 | XP_004297533.1 | 5 | 20506253..20513461 | 2694 | 897 | 99.00 | 8.65 | 43.88 | Chlo/Cyto |
| FvTrihelix15 | XM_004309446.2 | XP_004309494.1 | 5 | 151517..156036 | 2310 | 769 | 83.55 | 5.56 | 66.17 | Nucl |
| FvTrihelix16 | XM_004300273.2 | XP_004300321.1 | 5 | 22767294..22768593 | 792 | 263 | 29.49 | 9.40 | 50.32 | Nucl |
| FvTrihelix17 | XM_004301682.2 | XP_004301730.2 | 5 | 22820370..22821359 | 990 | 329 | 37.67 | 9.32 | 50.04 | Nucl |
| FvTrihelix18 | XM_004300356.2 | XP_004300404.1 | 5 | 24151374..24159101 | 840 | 279 | 32.74 | 8.91 | 52.34 | Nucl |
| FvTrihelix19 | XM_004302539.2 | XP_004302587.1 | 6 | 7489058..7490852 | 948 | 315 | 36.96 | 6.38 | 50.81 | Nucl |
| FvTrihelix20 | XM_011468504.1 | XP_011466806.1 | 6 | 13448505..13450635 | 1470 | 489 | 55.60 | 6.14 | 58.04 | Nucl |
| FvTrihelix21 | XM_004305323.2 | XP_004305371.1 | 6 | 13433800..13437353 | 1533 | 510 | 55.46 | 7.60 | 40.45 | Chlo |
| FvTrihelix22 | XM_004305314.2 | XP_004305362.1 | 6 | 13068235..13071846 | 1740 | 579 | 65.77 | 6.42 | 51.56 | Nucl |
| FvTrihelix23 | XM_011468923.1 | XP_011467225.1 | 6 | 20865049..20867313 | 1689 | 562 | 63.95 | 5.83 | 57.07 | Nucl |
| FvTrihelix24 | XM_004302867.2 | XP_004302915.1 | 6 | 11311732..11314065 | 1188 | 395 | 45.27 | 4.68 | 52.21 | Nucl |
| FvTrihelix25 | XM_004305721.2 | XP_004305769.1 | 6 | 24878967..24881044 | 1137 | 378 | 40.84 | 9.78 | 48.36 | Chlo/Nucl |
| FvTrihelix26 | XM_004306551.2 | XP_004306599.1 | 7 | 2582175..2584656 | 1695 | 564 | 63.61 | 6.13 | 67.38 | Nucl |
| FvTrihelix27 | XM_004307044.2 | XP_004307092.1 | 7 | 10119028..10121258 | 1005 | 334 | 37.01 | 9.37 | 64.51 | Nucl |
| FvTrihelix28 | XM_004308502.2 | XP_004308550.1 | 7 | 9002471..9004601 | 1302 | 433 | 48.97 | 6.19 | 35.38 | Nucl |
| FvTrihelix29 | XM_004308900.2 | XP_004308948.2 | 7 | 18420295..18421713 | 684 | 227 | 25.75 | 8.29 | 52.00 | Nucl |
| FvTrihelix30 | XM_011472120.1 | XP_011470422.1 | 7 | 23036260..23040696 | 2871 | 956 | 107.64 | 6.73 | 49.91 | Chlo/Nucl. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Jiang, F.; Sun, H.; He, T.; Liu, Y.; Jiao, G.; Ahmad, B.; Bokhari, S.A.M.; Chen, Q.; Wen, Z. Expression Analysis of Trihelix Transcription Factor Family in Strawberries and Functional Characterization of FvTrihelix6. Horticulturae 2023, 9, 633. https://doi.org/10.3390/horticulturae9060633
Fan J, Jiang F, Sun H, He T, Liu Y, Jiao G, Ahmad B, Bokhari SAM, Chen Q, Wen Z. Expression Analysis of Trihelix Transcription Factor Family in Strawberries and Functional Characterization of FvTrihelix6. Horticulturae. 2023; 9(6):633. https://doi.org/10.3390/horticulturae9060633
Chicago/Turabian StyleFan, Jianshuai, Fan Jiang, Hongyuan Sun, Tiannan He, Yuhan Liu, Gaozhen Jiao, Bilal Ahmad, Syeda Anum Masood Bokhari, Qingxi Chen, and Zhifeng Wen. 2023. "Expression Analysis of Trihelix Transcription Factor Family in Strawberries and Functional Characterization of FvTrihelix6" Horticulturae 9, no. 6: 633. https://doi.org/10.3390/horticulturae9060633
APA StyleFan, J., Jiang, F., Sun, H., He, T., Liu, Y., Jiao, G., Ahmad, B., Bokhari, S. A. M., Chen, Q., & Wen, Z. (2023). Expression Analysis of Trihelix Transcription Factor Family in Strawberries and Functional Characterization of FvTrihelix6. Horticulturae, 9(6), 633. https://doi.org/10.3390/horticulturae9060633