Abstract
Holoparasitic broomrape weeds (Orobanche and Phelipanche spp.) are a major biotic constraint limiting the production of important food crops. Parasitism caused by Phelipanche spp. is considered to be a serious threat, particularly in the Mediterranean region. In this study, the broomrape species collected from infested fennel fields in Sid L’Mokhtar in the Central West of Morocco was described and identified morphologically and molecularly. For molecular identification, 700 bp ITS regions were PCR-amplified from two representative individuals, OA and OY-4. The obtained sequences exhibited more than 99% similarity with the P. schultzii sequence (MT026593). In addition, maximum likelihood analysis placed our species with other referred P. schultzii in the same clade. Through molecular analysis, the broomrape species attacking fennel was identified as P. schultzii. This is the first record of the broomrape species P. schultzii parasitizing fennel (Foeniculum vulgare syn. Foeniculum officinale) in Morocco. Therefore, it should be considered that this parasite could spread to other cultivated fields causing severe yield losses, requiring urgent action through efficient control measures to avoid further spread of this parasitic weed.
1. Introduction
Parasitic weeds of the genera Orobanche and Phelipanche are considered to be a major problem and serious threat to agriculture worldwide due to their tremendously high impact on several agricultural crops and the difficulty of their eradication through common weed control measures. Belonging to the family Orobanchaceae, these parasitic weeds are known to have evolved a particular multicellular organ, termed a haustorium, that invades the host, forms a connection with the conductive tissue, and withdraws the parasite’s needed nutrients and water [1]. The broomrape is the common name for obligatory holoparasitic herbaceous plants of the genera Orobanche L. and Phelipanche L. There are about 200 species that debilitate several important dicotyledonous crops from various botanical families, including Fabaceae, Solanaceae, Asteraceae, Apiaceae, Brassicaceae, and Cucurbitaceae [2,3]. Due to their lack of chlorophyll, these parasitic weeds depend entirely on their hosts for mineral nutrients and water requirements [4]. Their diversity range focuses mainly in the Mediterranean region, Asia, and Southern and Eastern Europe [5,6]. Broomrape parasitism causes extensive yield losses and crop damage can range from 5% to 100%, depending on host susceptibility, infestation intensity, and environmental conditions [7].
Regarding the taxonomy of the genera Orobanche/Phelipanche and according to Beck-Mannagetta’s (1930) monograph, most authors have divided the genus Orobanche into four sections: Orobanche (=Osproleon Wallr.), Trionychon Wallr., Gymnocaulis Nutt., and Myzorrhiza (Philippi) Beck [8,9,10]. However, at the beginning of the 21st century, on the basis of morphological, karyological, and molecular phylogenetic analyses, the genus Orobanche was divided into two separate genera: Orobanche and Phelipanche [11,12]. Hence, some species belonging to the old genus Orobanche have been transferred to the newly formed genus Phelipanche. Compared to the genus Orobanche, the genus Phelipanche is characterized by stems generally branched above ground and the presence of bracteoles. There are several Phelipanche species, of which the major ones are P. ramosa, P. aegyptiaca, P. rosamrina, and P. schultzii. Differentiation between these species is based mainly on stem thickness, inflorescence type, pilosity of anthers, corolla form/color, and calyx teeth length [13,14]. Phelipanche ramosa is characterized by a simple or branched stem with glandular hairs, cream to mauve in color. The inflorescence is generally lax, with few to many flowers, sometimes with pedicelled basal flowers. The bracts are ovate-lanceolate with glandular hairs, while the bracteoles are linear-lanceolate. The calyx is bluish gray with glandular hairs and with subequal teeth that gradually narrow. The corolla is generally erect and distally tubular-infundibuliform, slightly curved, white at the base to bluish-white or dark blue at the apex, with glandular hairs. The upper lip is somewhat bilobed, while the lobes of the lower lip are pointed or obtuse, with more or less entire margins. Phelipanche aegyptiaca exhibits a branched stem usually 20–35 cm high, sometimes larger. The flowers are above but longer and the connective is hairy between the anther lobes. The Phelipanche rosmarina stem is simple with glandular hairs. The inflorescence is moderately loose, usually with few flowers. The bracts are ovate and glandular, while the bracteoles are narrowly ovate. The calyx has subequal teeth and glandular hairs. The corolla is erect or erect-spreading and dark blue in color with scattered glandular hairs. The upper lip is bilobed, obtuse, and slightly crenulated, and the lower lip has equal and rounded lobes. The stigmas are yellowish. Phelipanche schultzii displays a simple but sometimes branched stem. The inflorescence is cylindrical and gradually narrows towards the top. The flowers are upright, with some of the lower ones occasionally pedicellate, and they have lanceolate bracts with glandular hairs and narrow bracteoles. The calyx teeth are longer than the tube, with glandular hairs. The violet-blue corolla is narrow, with elliptical lobes on the lower lip.
In Morocco, the root parasitic species described above are among the parasitic genera recorded in the country. Since broomrape invaded Morocco, researchers have conducted investigations to identify and characterize the different species of these parasitic weeds and their respective host crops. Among the parasitic weeds, four main species have been reported as major constraints for many important agricultural crops in the country, including O. crenata Forsk., O. cumana Wallr., P. ramosa Pomel., and O. foetida Poir. In Morocco, a broomrape infestation by O. crenata on faba bean was officially reported in 1943 in the Fes region. Ever since, infestation by this parasitic weed has expanded to other zones (Zair, Pre-Rif, Chaouia, Sais, Fez, Meknes, Taounate, Sidi Kacem, Chefchaouen, Doukala, and Abda) [15,16] and the rate of field infestation progressed from 12% in 1981, to 26% in 1994, 30% in 2001, and reached 51% in 2013 [17,18]. Orobanche crenata then became a serious threat not only to faba bean but also to lentil and chickpea [19,20]. The parasitic weed O. cumana was first identified on sunflower in 2016 in the Kenitra province [21]. In 2004, O. foetida was found infecting wild legumes and common vetch [22,23]. Additionally, the branched broomrape P. ramosa has reportedly parasitized different cultivated crops, including tomato in the Abda region; carrot in the Abda, Saïs, and Chaouia regions; and lentil in the Doukkala, Chaouia, and Saïs regions [24]. The reported data indicates that these broomrape species have proven to be highly destructive parasitic plants threatening a plethora of hosts across multiple environments [20,25,26,27,28,29,30,31]. Facing this scourge, many control strategies have been used, including cultural practices, herbicidal control, and biological methods [32,33,34]. The development of resistant varieties is proposed as the most environmentally friendly control method [35,36]. Until now, no effective control strategy has yet to be developed [4]. The development of a promising breeding program requires a better understanding and deep comprehension of broomrape species and their interactions with hosts at morphological and molecular levels. However, updated data and information about the occurrence, present distribution, preferred host plants, and infestation levels of different Orobanche/Phelipanche species in Morocco are still lacking. A species of broomrape was recorded growing on fennel for the first time in Morocco. Hence, the current study was conducted to identify this holoparasitic plant species found in the Central West of Morocco by performing morphological and molecular analyses.
2. Materials and Methods
2.1. Plant Material
During the cropping season 2019–2020, several field surveys were carried out in different regions of Morocco. In April 2019, a survey of agricultural fields revealed the infestation of a broomrape species on fennel plants (Foeniculum vulgare Mill.) in Chichaoua province, Marrakech-Safi region, Morocco (31°35′27.5″ N; 9°06′16.9″ W, 365 m above sea level). This province is characterized by a semi-arid to arid climate with low rainfall, of which the majority of cultivated agricultural crops are mainly irrigated.
Several individuals of the collected broomrape species were sampled and only two representative individuals (OA and OY-4) were used for molecular analysis. The parasitism was confirmed by carefully excavating the fennel plant by verifying the attachment of the parasite to the host’s roots. Specimens were examined, photographed in the field, collected, and then taken to the lab. Sample examination and identification were carried out on fresh materiel. One specimen was dried and deposited in the National Herbarium of Morocco at the Scientific Institute of Rabat.
2.2. Morphological Characterization
The identification of the collected broomrape plant was performed on the basis of its morphological characteristics. The samples were categorized on the following characteristics: plant height, stem, flowers, calyx, corolla, stamens, anthers, and ovary. The plant was observed under a light microscope.
2.3. DNA Extraction, PCR and Sequencing
Total genomic DNA was extracted from 50 mg of fresh material using a standard cetyl trimethyl ammonium bromide (CTAB) protocol (1M Tris-HCl, 5M NaCl, 0.5 M EDTA, 2% (w/v) CTAB, and 0.2% (v/v) β-mercaptoethanol) with slight modifications [37]. The DNA quantity was assessed by electrophoresis on 1.8% agarose gel stained with ethidium bromide and visualized under ultraviolet (UV) light, and the DNA quality and concentration were measured spectroscopically using a NanoDrop Nano-100 microspectrophotometer.
The polymerase chain reaction (PCR) was performed using the universal plant internal transcribed spacer (ITS) primers ITS-p5 (5′ CCTTATCAYTTAGAGGAAGGAG 3′) and ITS-u4 (5′ RGTTTCTTTTCCTCCGCTTA 3′) designed by [38]. The PCR reaction was performed with an automated thermocycler (Aeris™ Thermal Cycler Model G96) using 25 µL of reaction volume containing 1 x buffer, 1 U of Taq™ DNA polymerase (BIOLINE, London, UK), 0.5 μM of each primer, 15.3 μL of ultrapure water, and 1 µL of DNA template. The ITS-p5 and ITS-u4 regions were amplified according to the following PCR conditions: initial denaturation at 94 °C for 4 min, followed by 34 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 40 s, and elongation at 72 °C for 1 min. The PCR process was completed with a final extension at 72 °C for 10 min.
PCR amplicons were separated by electrophoresis on 1.8% agarose gel at 100 V for 1.5 h and visualized by staining with ethidium bromide for 20 min. The amplified DNA fragments were observed under a UV transilluminator and photographed using the GelDoc XR+ system (Bio-Rad, Hercules, CA, USA)) with Image Lab Software. After amplification, the PCR products were purified with the ExS Pure™ Enzymatic PCR Cleanup Kit. The purified PCR products were sequenced using the ABI3500 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) and the BigDye X-terminator v3.1 Cycle Sequencing Kit (Applied Biosystems) was used according to the supplier’s protocol.
2.4. Phylogenetic Analysis
Phylogenetic analysis was conducted using MEGA7 and multiple sequence alignment was carried out with ClustalW embedded in this software [39]. The nucleotide sequences were submitted to NCBI (National Center for Biotechnology Information, Bethesda, MD, USA) (https://www.ncbi.nlm.nih.gov, accessed on 2 March 2023) and analyzed using BLASTn available online (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 2 March 2023) to identify sequences of interest by alignment with known sequences. As the outgroup, the sequence of Digitalis purpurea belonging to the family Scrophulariaceae (AY911261) was used. The maximum likelihood (ML) approach using heuristic searches was applied to construct a phylogenetic tree and other parameters were set as default. The bootstrap method with 1000 replicates was applied to estimate the robustness of clades [40]. Generated sequences for this study were deposited in the NCBI database. All sequences included in this study, voucher information (or reference), and GenBank accession numbers are given in Table 1.

Table 1.
Lists of taxon names, voucher information or literature reference, and GenBank accession numbers. Sequences of unknown origin are marked by a question mark and those with unavailable voucher information are marked by an asterisk.
3. Results
3.1. Description and Morphological Identification of Broomrape Species
From a field survey, the authors found that a fennel-growing field in Sid L’Mokhtar locality, Chichaoua province, Marrakech Safi region (Central West, Morocco) was higly infested with a branched broomrape species (Figure 1A). The attachment of this parasitic plant to fennel roots was confirmed by checking the haustorial connection between host and parasite (Figure 1B). According to farmers’ statements, the average yield losses in all surveyed locations were estimated at 20% to 50%, depending on the field infestation level.

Figure 1.
(A) Fennel plants infected by the branched broomrape P. schultzii; (B) broomrape shoots attached to fennel roots.
The parasite’s taxonomy was defined based on the following main botanical traits: Up to 50 cm plant length. Stem erect with lateral buds. Flowers surrounded by one triangulate ovate bract, 8 to 11 mm of reddish brown color (Figure 2A,D), and two lateral 8–10 mm lanceolate bracteoles (Figure 2E). Gamosepalous, campanulate, and cupiliform calyx 10–14 mm in length and four unequal divisions with welded sepals (Figure 2B). Calyx teeth much longer than the tube with 3 to 6 sepals and corolla with three-lobed lower lip and bilobed upper lip. Tubular corolla 15 mm in length and five light purple petals (Figure 2C). Whitish, epipetalous, dyalistemonous, and pubescent stamens with brown and hairy anthers surrounding one whitish stigma with inferior ovary (Figure 2F). Capsule with several hundred narrow, oblong, and dust-like seeds.
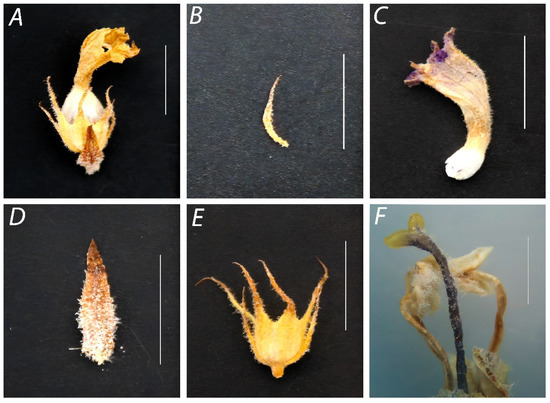
Figure 2.
Morphological characterization of the parasitic plant flower. (A) Whole flower; (B) calyx; (C) corolla; (D) bract; (E) bracteole; (F) stamens, style, and stigma. The white bar represents a scale of 1 cm.
3.2. Molecular Analysis
3.2.1. DNA Amplification
Internal transcribed spacers (ITS) of nuclear ribosomal DNA are the most commonly used markers in plant molecular systematics and DNA barcoding analyses at the genus and species levels because of the presence of highly variable loci. These DNA fragments were used in the present study to examine and infer phylogenetic relationships of our two individual P. schultzii samples with other broomrape species.
The broomrape species found was molecularly identified by amplifying its genomic DNA using the ITS sequences ITS-p5 and ITS-u4 as universal primers. A single specific PCR-amplified band of approximately 700 bp was generated (Figure 3). The nucleotide sequences of the PCR-amplified products were determined by Sanger sequencing, involving 592 nucleotide sequences in the final dataset.
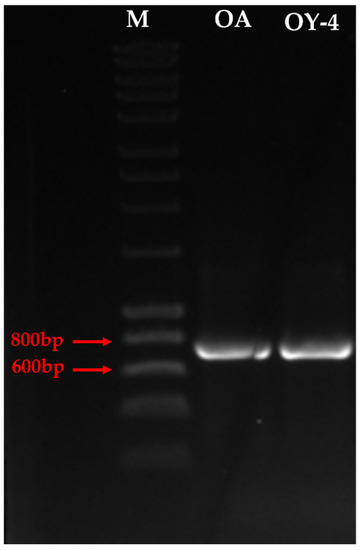
Figure 3.
PCR-amplified products of ITS primers from two broomrape representative individuals OA and OY-4. M: 1 kb DNA size marker.
3.2.2. Sequence Analysis of the ITS-p5 and ITS-u4 rDNA Regions
After sequencing the PCR products and analysis with BLASTn, the results indicated that the sequences of the two broomrape samples had more than 99% sequence homology with the Phelipanche schultzii nucleotide sequence contained in the NCBI database under accession number MT026593. The GenBank accession numbers of our samples OA and OY-4 were OM475767 and OM475768, respectively. The two reported P. schultzii sequences described in the current study showed 100% similarity with each other.
3.2.3. Phylogenetic Analysis
To further determine the broomrape species causing the parasitism on fennel, a phylogenetic tree was constructed using the ITS sequences of this study and corresponding sequences from other species. Analysis of the phylogenetic tree based on the maximum likelihood method revealed that our samples were gathered together with the broomrape species downloaded from GenBank in the same branch of the phylogenetic tree. The relationship between the P. schultzii sequences was strongly close and the similarity was over 88% (Figure 4). Moreover, the divergent branch of broomrape species was occupied by Digitalis purpurea (outgroup).

Figure 4.
Phylogenetic tree constructed according to the maximum likelihood method using gene sequences of ITS-P5 and ITS-u4 regions. Numbers at branches indicate the bootstrap support values (expressed as percentages) calculated for 1000 replications. Digitalis purpurea (Scrophulariaceae) acts as outgroup in this analysis.
4. Discussion
F. vulgare Mill., commonly known as fennel, figures among the most important and highly valued medicinal and aromatic plants in the world, especially in Mediterranean countries including Morocco [45]. Nevertheless, as with any crop, the fennel plant is subjected to a series of abiotic and biotic stresses. Parasitic weeds represent a severe constraint for several important agricultural crops in Morocco. In this work, we report the infestation of fennel plants by a parasitic weed. Based on the identification keys of Phelipanche species [13], P. schultzii was clearly distinguishable from the related species P. ramosa, P. rosmarina, and P. aegyptiaca by having few or no pedicellate flowers and more or less acute lower corolla lip. Furthermore, P. schultzii was easily separated from P. ramosa and P. rosmarina by having distinctly longer calyx teeth than the tube, whereas the two other species have subequal teeth. These morphological features of the P. schultzii species were correlated with those of our two representative individuals OA and OY-4, in which the inflorescence was characterized by a few pedicellate flowers, acute lower corolla lip, and the calyx teeth were distinctly longer than the tube (Figure 2).
The implementation of new molecular tools such as PCR amplification, sequencing, and sequence analysis has enabled reliable identification of different parasitic species. Comparison of the ITS regions is one of the most reliable tools for taxonomy and molecular phylogeny [38,46]. In several previous studies, these markers have shown a high degree of variation in closely related Orobanche species [42,47,48,49]. In fact, the phylogenetic tree, generated based on the ITS nuclear regions, showed the clustering of each species in a separate subgroup. The sequences of our two representative individuals OA and OY-4 were clustered together in the same clade according to their similarity with the referenced P. schultzii. Additionally, they were phylogenetically distant to P. ramosa and P. aegyptiaca, which have been reported in fennel host plants. Both morphological and molecular characterization clearly confirmed the identity of the two broomrape individuals as P. schultzii.
According to previous studies, P. schultzii (Mutel) Pomel (syn. Orobanche schultzii Mutel) has occurred in many countries in the Mediterranean basin, such as Algeria [50], Jordan [51], Irak [52,53], Italy [54,55,56], Spain [57,58,59,60], and the Canary Islands [61]. This parasitic plant is also present in Portugal, where it has been described as Orobanche trichocalyx f. lusitanica Welw [61]. Unlike P. ramosa, which has a very wide range of host plants including cultivated crops and weeds, P. schultzii has a comparatively small host range [7,14]. This parasitic plant has been reported on different host plants; it was found infesting Galium spp. (Rubiaceae) [53], Distichoselinum tenuifolium (Lag.) (Apiaceae) [57], and Scorpiurus (Fabaceae) [62]. In Jordan, P. schultzii has been found to parasitize olive and almond trees [63]. No previous study reported infection of F. vulgare Mill. by P. schultzii. However, several studies have reported the susceptibility of fennel to broomrape species in different countries. In Jordan, fennel has been recorded as a host for P. ramosa, which is considered to be the most common parasitic plant, showing light-to-high infestation levels with common prevalence [64,65]. Furthermore, in pot experiments under glasshouse conditions, Qasem and Foy [66] reported a moderate infestation level of fennel by P. ramosa (3.3 plant parasites per host plant). In Sudan, this parasitic weed has been reported on fennel plants, where researchers identified the incidence as 12–40 P. ramosa plant parasites/host plant [67]. In Italy, it has been reported that fennel is vulnerable to the branched broomrape P. ramosa [68,69]. Additionally, fennel has been reported to be a host for P. aegyptiaca. In fact, a rare prevalence of this parasite was observed by Khatri, Jha [70] in Nepal. Moreover, Parker and Riches [2] reported that fennel plants had been lightly attacked by P. aegyptiaca and P. ramosa. Within the genus Orobanche, fennel plants were also reportedly parasitized by O. crenata in the study by Foy, Jain [71]. Therefore, the high susceptibility of fennel to the broomrape species P. schultzii may allow the spread of this parasitic weed especially in Chichaoua province and even in other Moroccan localities. This in turn could increase the level of infestation in subsequent years, which will pose a significant threat to important socio-economic crops in the country such as olive and almond trees, which were previously reported as host plants for P. schultzii. Furthermore, broomrape populations endure severe modifications in allele frequencies due to the use of control methods and the adaptation of these parasitic weeds to a wide range of host plants [72]. For this reason, it would be relevant to monitor variations in the genetic diversity of broomrape populations collected from the same field for many consecutive years. Understanding the relationship between levels of genetic variability and the ecological and life-history traits within and among parasitic plant populations is crucial to successful selection programs and breeding approaches for host plants in different geographical zones. Information on plant genetic diversity gained by molecular analysis with the use of specific DNA markers will help to identify the evolutionary processes responsible for the genetic structure of plant populations [73,74].
In Morocco, fennel is considered among the most important aromatic medicinal plants and culinary herbs commonly cultivated in large fields for its bulb and seeds [75,76]. Phelipanche schultzii, as reported in this study, attaches to the fennel plant’s roots and deprives it of necessary nutrients, leading to wilting and affecting growth, resulting in a decrease in fennel seed yield and quality. This can have economic implications for fennel growers and producers. Therefore, it is imperative to raise awareness among horticulturists about the potential threat posed by this parasitic plant to fennel production and cultivation in the country.
5. Conclusions
The parasitic plant P. schultzii was reported for the first time infesting fennel (Foeniculum vulgare Mill.) in Morocco. Both morphological characterization and molecular analyses were used to accurately identify this parasitic plant. Our findings make a significant contribution to the horticultural community by providing important insights into the first occurrence of P. schultzii on fennel in Morocco. Considering the infestation level of fennel fields and important yield losses, high-priority actions need to be considered for the development of an integrated successful management and control strategy against P. schultzii.
Author Contributions
Conceptualization, M.A., Z.E.A.T., S.K. and R.M.; methodology, M.E.A., M.A., E.M.K., Z.E.A.T., S.K. and R.M.; software, M.E.A., E.M.K., S.K. and R.M.; validation, M.A., S.K. and R.M.; formal analysis, M.E.A., S.K. and R.M.; investigation, M.E.A., Z.E.A.T. and R.M.; resources, M.A., Z.E.A.T., S.K. and R.M.; data curation, M.E.A., E.M.K. and R.M.; writing—original draft preparation, M.E.A., M.A. and R.M.; writing—review and editing, M.E.A., M.A., Z.E.A.T., S.K. and R.M.; visualization, M.E.A., M.A., S.K. and R.M.; supervision, M.A., Z.E.A.T., S.K. and R.M.; project administration, M.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data are in the manuscript.
Acknowledgments
This research was supported by National Institute of Agricultural Research (INRA, Morocco) and Ministry of Higher Education, Scientific Research and Professional Training of Morocco (MESRSFC) through funding of “DIVICIA” project within the PRIMA program supported under Horizon 2020 of the European Union’s Framework Program for Research and Innovation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The haustorium, a specialized invasive organ in parasitic plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Westwood, J.H.; Depamphilis, C.W.; Das, M.; Fernández-Aparicio, M.; Honaas, L.A.; Timko, M.P.; Wafula, E.K.; Wickett, N.J.; Yoder, J.I. The parasitic plant genome project: New tools for understanding the biology of Orobanche and Striga. Weed Sci. 2012, 60, 295–306. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Reboud, X.; Gibot-Leclerc, S. Broomrape weeds. Underground mechanisms of parasitism and associated strategies for their control: A review. Front. Plant Sci. 2016, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Delavault, P. Knowing the parasite: Biology and genetics of Orobanche. Helia 2015, 38, 15–29. [Google Scholar] [CrossRef]
- Joel, D.; Hershenhorn, J.; Eizenberg, H.; Aly, R.; Ejeta, G.; Rich, P.; Ransom, J.; Sauerborn, J.; Rubiales, D. Biology and management of weedy root parasites. Hortic. Rev. 2007, 33, 267–349. [Google Scholar]
- Parker, C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Holub, J. New names inPhanerogamae 6. Folia Geobot. Phytotaxon. 1977, 12, 417–432. [Google Scholar] [CrossRef]
- Sojak, J. Nomenklatorické poznámky (Phanerogamae). Casopis 1972, 140, 127–134. [Google Scholar]
- Teryokhin, E. Weed Broomrapes: Systematics, Ontogenesis, Biology, Evolution; Aufstieg Verlag: Lanshut, Germany, 1997. [Google Scholar]
- Joel, D. The new nomenclature of Orobanche and Phelipanche. Weed Res. 2009, 49, 6–7. [Google Scholar] [CrossRef]
- Schneeweiss, G.M. Correlated evolution of life history and host range in the nonphotosynthetic parasitic flowering plants Orobanche and Phelipanche (Orobanchaceae). J. Evol. Biol. 2007, 20, 471–478. [Google Scholar] [CrossRef]
- Castroviejo, S.; Muñoz Garmendia, F. Flora Ibérica: Plantas Vasculares de la Península Ibérica e Islas Baleares: Vol. IX; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2015; pp. 1–612. [Google Scholar]
- Parker, C. The parasitic weeds of the Orobanchaceae. In Parasitic Orobanchaceae; Springer: Berlin/Heidelberg, Germany, 2013; pp. 313–344. [Google Scholar]
- FAO. The Problem of Orobanche spp. in Africa and Near East. Food and Agriculture Organization of the United Nations, 2014. Available online: http://www.fao.org/agriculture/crops/thematic-sitemap/theme/biodiversity/weeds/issues/oro/en/ (accessed on 19 December 2022).
- Labrada, R. Farmer training on parasitic weed management. In Progress on Farmer Training in Parasitic Weed Management; Labrada, R., Ed.; FAO: Rome, Italy, 2007; pp. 1–5. [Google Scholar]
- Sauerborn, J. Parasitic Flowering Plants: Ecology and Management; Verlag Josef Margraf: Wiekersheim, Germany, 1991. [Google Scholar]
- Stewart, G.R.; Press, M.C. The physiology and biochemistry of parasitic angiosperms. Annu. Rev. Plant Biol. 1990, 41, 127–151. [Google Scholar] [CrossRef]
- Mabsoute, L.; Saadaoui, E. Acquis de recherche sur le parasitisme des légumineuses alimentaires au Maroc: Synthese bibliographique. Al Awamia 1996, 92, 55–67. [Google Scholar]
- Ennami, M.; Briache, F.Z.; Gaboun, F.; Abdelwahd, R.; Ghaouti, L.; Belqadi, L.; Westwood, J.; Mentag, R. Host differentiation and variability of Orobanche crenata populations from legume species in Morocco as revealed by cross-infestation and molecular analysis. Pest Manag. Sci. 2017, 73, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Nabloussi, A.; Velasco, L.; Assissel, N. First report of sunflower broomrape, Orobanche cumana Wallr., in Morocco. Plant Dis. 2018, 102, 457. [Google Scholar] [CrossRef]
- Rubiales, D.; Sadiki, M.; Román, B. First report of Orobanche foetida on common vetch (Vicia sativa) in Morocco. Plant Dis. 2005, 89, 528. [Google Scholar] [CrossRef] [PubMed]
- Vaz Patto, M.; Díaz-Ruiz, R.; Satovic, Z.; Román, B.; Pujadas-Salvà, A.; Rubiales, D. Genetic diversity of Moroccan populations of Orobanche foetida: Evolving from parasitising wild hosts to crop plants. Weed Res. 2008, 48, 179–186. [Google Scholar] [CrossRef]
- Zehhar, N. Recherche des Mécanismes Impliqués dans la Germination d’Orobanche ramosa L. et de Sources et de Mécanismes de Résistance à ce Parasite; Etude Préliminaire de Possibilités de Lutte Chimique Contre Orobanche ramosa et Cuscuta Lupuliformis Krock. Master’s Thesis, Université Sidi Mohamed Ben Abdellah, Fès, Morocco, 2003. [Google Scholar]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Gaboun, F.; Abdelwahd, R.; Fatemi, Z.E.A.; El-Rodeny, W.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Field and controlled conditions screenings of some faba bean (Vicia faba L.) genotypes for resistance to the parasitic plant Orobanche crenata Forsk. and investigation of involved resistance mechanisms. J. Plant Dis. Prot. 2019, 126, 211–224. [Google Scholar] [CrossRef]
- Briache, F.Z.; Ennami, M.; Mbasani-Mansi, J.; Lozzi, A.; Abousalim, A.; Rodeny, W.E.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Effects of salicylic acid and indole acetic acid exogenous applications on induction of faba bean resistance against Orobanche crenata. Plant Pathol. J. 2020, 36, 476–490. [Google Scholar] [CrossRef]
- Ennami, M.; Briache, F.; Mansi, J.; Gaboun, F.; Ghaouti, L.; Belqadi, L.; Mentag, R. Genetic diversity of Moroccan Orobanche crenata populations revealed by sequence-related amplified polymorphism markers. J. Agric. Sci. 2017, 9, 164. [Google Scholar] [CrossRef]
- Ennami, M.; Mbasani-mansi, J.; Briache, F.Z.; Oussible, N.; Gaboun, F.; Ghaouti, L.; Belqadi, L.; Ghanem, M.E.; Aberkani, K.; Westwood, J.; et al. Growth-defense tradeoffs and source-sink relationship during both faba bean and lentil interactions with Orobanche crenata Forsk. Crop Prot. 2020, 127. [Google Scholar] [CrossRef]
- Mbasani-Mansi, J.; Briache, F.Z.; Ennami, M.; Gaboun, F.; Benbrahim, N.; Triqui, Z.E.A.; Mentag, R. Resistance of Moroccan lentil genotypes to Orobanche crenata infestation. J. Crop Improv. 2019, 33, 306–326. [Google Scholar] [CrossRef]
- Abdallah, F.; Kumar, S.; Amri, A.; Mentag, R.; Kehel, Z.; Mejri, R.K.; Triqui, Z.E.A.; Hejjaoui, K.; Baum, M.; Amri, M. Wild Lathyrus species as a great source of resistance for introgression into cultivated grass pea (Lathyrus sativus L.) against broomrape weeds (Orobanche crenata Forsk. and Orobanche foetida Poir.). Crop Sci. 2021, 61, 263–276. [Google Scholar] [CrossRef]
- Briache, F.Z.; El Amri, M.; Ennami, M.; Amri, M.; Triqui, Z.E.A.; Mentag, R. Induction of systemic resistance to Orobanche crenata in lentil by exogenous application of salicylic acid and indole acetic acid. J. Plant Prot. Res. 2023, 63, 83–96. [Google Scholar]
- Bayoumi, T.; Ammar, S.M.; El-Bramawy, M.; Emam, M. Effect of some broomrape control methods on growth and seed yield attributes of faba bean Vicia faba, L. cultivars. J. Plant Prod. Sci. 2013, 1, 17–24. [Google Scholar]
- Fenández-Aparicio, M.; Sillero, J.C.; Rubiales, D. Intercropping with cereals reduces infection by Orobanche crenata in legumes. Crop Prot. 2007, 26, 1166–1172. [Google Scholar] [CrossRef]
- Amsellem, Z.; Barghouthi, S.; Cohen, B.; Goldwasser, Y.; Gressel, J.; Hornok, L.; Kerenyi, Z.; Kleifeld, Y.; Klein, O.; Kroschel, J. Recent advances in the biocontrol of Orobanche (broomrape) species. BioControl 2001, 46, 211–228. [Google Scholar] [CrossRef]
- Amri, M.; Trabelsi, I.; Abbes, Z.; Kharrat, M. Release of a new faba bean variety “Chourouk” resistant to the parasitic plants Orobanche foetida and O. crenata in Tunisia. Int. J. Agric. Biol. 2019, 21, 499–505. [Google Scholar] [CrossRef]
- Amri, M.; Abbes, Z.; Trabelsi, I.; Ghanem, M.E.; Mentag, R.; Kharrat, M. Chlorophyll content and fluorescence as physiological parameters for monitoring Orobanche foetida Poir. infection in faba bean. PLoS ONE 2021, 16, e0241527. [Google Scholar] [CrossRef]
- Lassner, M.W.; Peterson, P.; Yoder, J.I. Simultaneous amplification of multiple DNA fragments by polymerase chain reaction in the analysis of transgenic plants and their progeny. Plant Mol. Biol. Report. 1989, 7, 116–128. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- Swofford, D. PAUP* Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Piwowarczyk, R.; Schneider, A.C.; Góralski, G.; Kwolek, D.; Denysenko-Bennett, M.; Burda, A.; Ruraż, K.; Joachimiak, A.J.; Pedraja, Ó.S. Phylogeny and historical biogeography analysis support Caucasian and Mediterranean centres of origin of key holoparasitic Orobancheae (Orobanchaceae) lineages. PhytoKeys 2021, 174, 165. [Google Scholar] [CrossRef] [PubMed]
- Schneeweiss, G.M.; Colwell, A.; Park, J.-M.; Jang, C.-G.; Stuessy, T.F. Phylogeny of holoparasitic Orobanche (Orobanchaceae) inferred from nuclear ITS sequences. Mol. Phylogenet. Evol. 2004, 30, 465–478. [Google Scholar] [CrossRef]
- Parsai, S.Z.M.; Vaezi, J.; Ejtehadi, H.; Memariani, F.; Joharchi, M.R. Phelipanche pouyanii (Orobanchaceae), a new species from Iran. Nova Biol. Reper. 2021, 7, 457–467. [Google Scholar]
- Wolfe, A.D.; Randle, C.P.; Liu, L.; Steiner, K.E. Phylogeny and biogeography of Orobanchaceae. Folia Geobot. 2005, 40, 115–134. [Google Scholar] [CrossRef]
- Abdellaoui, M.; Kasrati, A.; El Rhaffari, L. The effect of domestication on seed yield, essential oil yield and antioxidant activities of fennel seed (Foeniculum vulgare Mill.) grown in Moroccan oasis. J. Assoc. Arab Univ. Basic Appl. Sci. 2017, 24, 107–114. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Piwowarczyk, R.; Denysenko-Bennett, M.; Góralski, G.; Kwolek, D.; Pedraja, Ó.S.; Cygan, M.; Joachimiak, A. Phylogenetic relationships within Orobanche and Phelipanche (Orobanchaceae) from Central Europe, focused on problematic aggregates, taxonomy, and host ranges. Acta Biol. Cracoviensia Ser. Bot. 2018, 60, 45–64. [Google Scholar]
- Li, X.; Feng, T.; Randle, C.; Schneeweiss, G.M. Phylogenetic relationships in Orobanchaceae inferred from low-copy nuclear genes: Consolidation of major clades and identification of a novel position of the non-photosynthetic Orobanche clade sister to all other parasitic Orobanchaceae. Front. Plant Sci. 2019, 10, 902. [Google Scholar] [CrossRef]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenetics Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Pomel, A. Nouveaux Matériaux Pour la Flore Atlantique; Savy: Paris, France, 1874. [Google Scholar]
- Qasem, J.R.S. Thesium humile. In Parasitic Weeds of Jordan: Species, Hosts, Distribution and Management; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022; pp. 304–314. [Google Scholar]
- Al-Mayah, A.A.; Al-Asadi, W.M. Two new records of the genus Phelipanche (Orobanchaceae) from Iraq. Glob. J. Biol. Agric. Health Sci. 2017, 6, 4–7. [Google Scholar] [CrossRef]
- Al-Asadi, W.M.; Al-Mayah, A.A. Three new specific records of Phelipanche (Orobanchaceae) to the Flora of Iraq. Calyx (mm) 2016, 5, 970–972. [Google Scholar]
- Conti, F.; Cancellieri, L.; Cangelmi, G.; Filibeck, G.; Rosati, L.; Bartolucci, F. New records of native and alien vascular plants from Abruzzo, Lazio and Molise National Park (Italy)–and additions to the flora of Abruzzo and Molise administrative regions. Ann. Di Bot. 2022, 12, 23–34. [Google Scholar]
- Domina, G.; Marino, P.; Castellano, G. The genus Orobanche (Orobanchaceae) in Sicily. Flora Mediterr. 2011, 21, 205–242. [Google Scholar]
- Raimondo, F.M.; Spadaro, V. Addenda et emendanda to the “A catalogue of plants growing in Sicily”. Flora Mediterr. 2009, 19, 303–312. [Google Scholar]
- Ruiz, L.C.; Gallo, M.L.; Moral, G.M.; Pedraja, O.S. Phelipanche cernua Pomel (Orobanchaceae), a prioritary name for the western mediterranean species recently redescribed as Ph. inexpectata. Flora Montiberica 2013, 54, 75–83. [Google Scholar]
- Castillejo, M.; FERNÁNDEZ-APARICIO, M.; Satovic, Z.; Rubiales, D. Comparative proteomic analysis of Orobanche and Phelipanche species inferred from seed proteins. Weed Res. 2009, 49, 81–87. [Google Scholar] [CrossRef]
- Navarro, J.G.; Gisbert, J.B.P.; Franzi, A.V.; Duato, E.S.; Gimeno, R.R. Las plantas de interés del NE de la Provincia de Albacete e inmediaciones de la Provincia de Valencia, VI. Sabuco: Rev. Estud. Albacet. 2011, 8, 117–142. [Google Scholar]
- Morales, R. Catálogo de plantas vasculares de la Comunidad de Madrid (España)[Catalogue of the vascular plants from Madrid Community (Spain)]. Bot. Complut 2003, 27, 31–70. [Google Scholar]
- Carlón, L.; Gómez Casares, G.; Laínz, M.; Moreno Moral, G.; Sánchez Pedraja, Ó.; Schneeweiss, G.M. Más, a Propósito de Algunas Phelipanche Pomel, Boulardia FW Schultz y Orobanche L. (Orobanchaceae) del Oeste del Paleártico; Jardín Botánico Atlántico: Gijón, Spain, 2008. [Google Scholar]
- Foley, M.Y. Orobanchaceae in the” flora iberica” área: New taxa, excluded-taxa, and typification. In Anales del Jardín Botánico de Madrid; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2001; pp. 223–233. [Google Scholar]
- Qasem, J.R.S. Parasitic Weeds of Jordan: Species, Hosts, Distribution and Management-Part I: Root Parasites; Orobanchaceae, Santalaceae & Cynomoryaceae; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022; p. 360. [Google Scholar]
- Qasem, J.R. Parasitic weeds of the Orobanchaceae family and their natural hosts in Jordan. Weed Biol. Manag. 2009, 9, 112–122. [Google Scholar] [CrossRef]
- Qasem, J.R. Parasitic flowering plants on cultivated plants in Jordan-the present status and management. Pak. J. Weed Sci. Res. 2010, 16, 227–239. [Google Scholar]
- Qasem, J.; Foy, C. Screening studies on the host range of branched broomrape (Orobanche ramosa). J. Hortic. Sci. Biotechnol. 2007, 82, 885–892. [Google Scholar] [CrossRef]
- Babiker, A.; Ahmed, E.; Dawoud, D.; Abdella, N. Orobanche species in Sudan: History, distribution and management. Sudan J. Agric. Res. 2007, 10, 107–114. [Google Scholar]
- Lops, F.; Frabboni, L.; Carlucci, A.; Tarantino, A.; Raimondo, M.L.; Disciglio, G. Management of Branched Broomrape in Field Processing Tomato Crop; IntechOpen: London, UK, 2022; Available online: https://www.intechopen.com/chapters/82794 (accessed on 18 February 2023).
- Conversa, G.; Bonasia, A.; Elia, A. Chemical control of branched broomrape in processing tomato using sulfonylureas in Southern Italy. Ital. J. Agron. 2017, 12, 939. [Google Scholar] [CrossRef]
- Khatri, G.; Jha, P.; Agrawal, V.; Jacobsohn, R. Distribution, host range and phenology of Orobanche spp. in Nepal. In Proceedings of the International Workshop on Orobanche Research, Obermarchtal, Germany, 19–22 August 1989; pp. 18–24. [Google Scholar]
- Foy, C.L.; Jain, R.; Jacobsohn, R. Recent approaches for chemical control of broomrape (Orobanche spp.). Rev. Weed Sci. 1989, 4, 123–152. [Google Scholar]
- Satovic, Z.; Joel, D.M.; Rubiales, D.; Cubero, J.I.; Romn, B. Population genetics in weedy species of Orobanche. Australas. Plant Pathol. 2009, 38, 228–234. [Google Scholar] [CrossRef]
- Darvishzadeh, R. Genetic variability, structure analysis, and association mapping of resistance to broomrape (Orobanche aegyptiaca Pers.) in tobacco. J. Agric. Sci. Technol. 2018, 18, 1407–1418. [Google Scholar]
- Hamrick, J.L.; Godt, M.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar]
- Aboukhalaf, A.; El Amraoui, B.; Tabatou, M.; da Rocha, J.M.F.; Belahsen, R. Screening of the antimicrobial activity of some extracts of edible wild plants in Morocco. Funct. Foods Health Dis. 2020, 10, 265–273. [Google Scholar] [CrossRef]
- El Ouariachi, E.; Lahhit, N.; Bouyanzer, A.; Hammouti, B.; Paolini, J.; Majidi, L.; Desjobert, J.; Costa, J. Chemical composition and antioxidant activity of essential oils and solvent extracts of Foeniculum vulgare Mill. from Morocco. J. Chem. Pharm. Res. 2014, 6, 743–748. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).