
Molecular Identification and Characterization of UDP-glycosyltransferase (UGT) Multigene Family in Pomegranate


Abstract
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of UGT Genes and Basic Physicochemical Properties of Proteins
2.2. Sequence Alignment and Phylogenetic Analysis
2.3. Chromosomal Distribution of PgUGTs and Gene Duplications
2.4. Exon–Intron Structures and Conserved Motifs
2.5. Promoter Cis-Acting Element Analysis
2.6. Gene Expression Analysis Using RNA-seq
2.7. RNA Isolation, Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR) Analysis
2.8. Functional Prediction of PgUGT Involved in Flavonoid Biosynthesis
3. Results
3.1. Identification of UGTs in Pomegranate
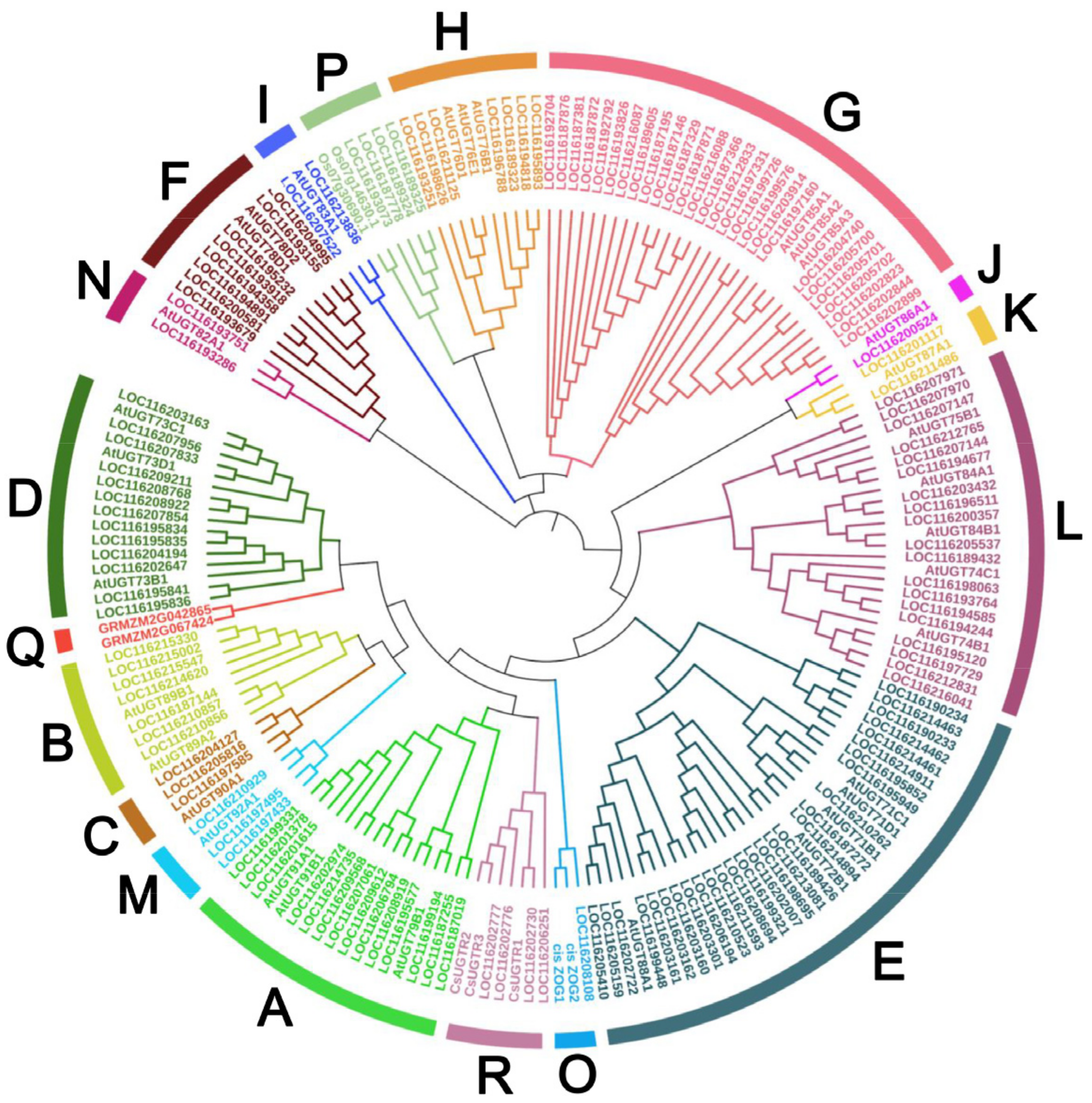
3.2. Phylogenetic Analysis of UGTs in Pomegranate
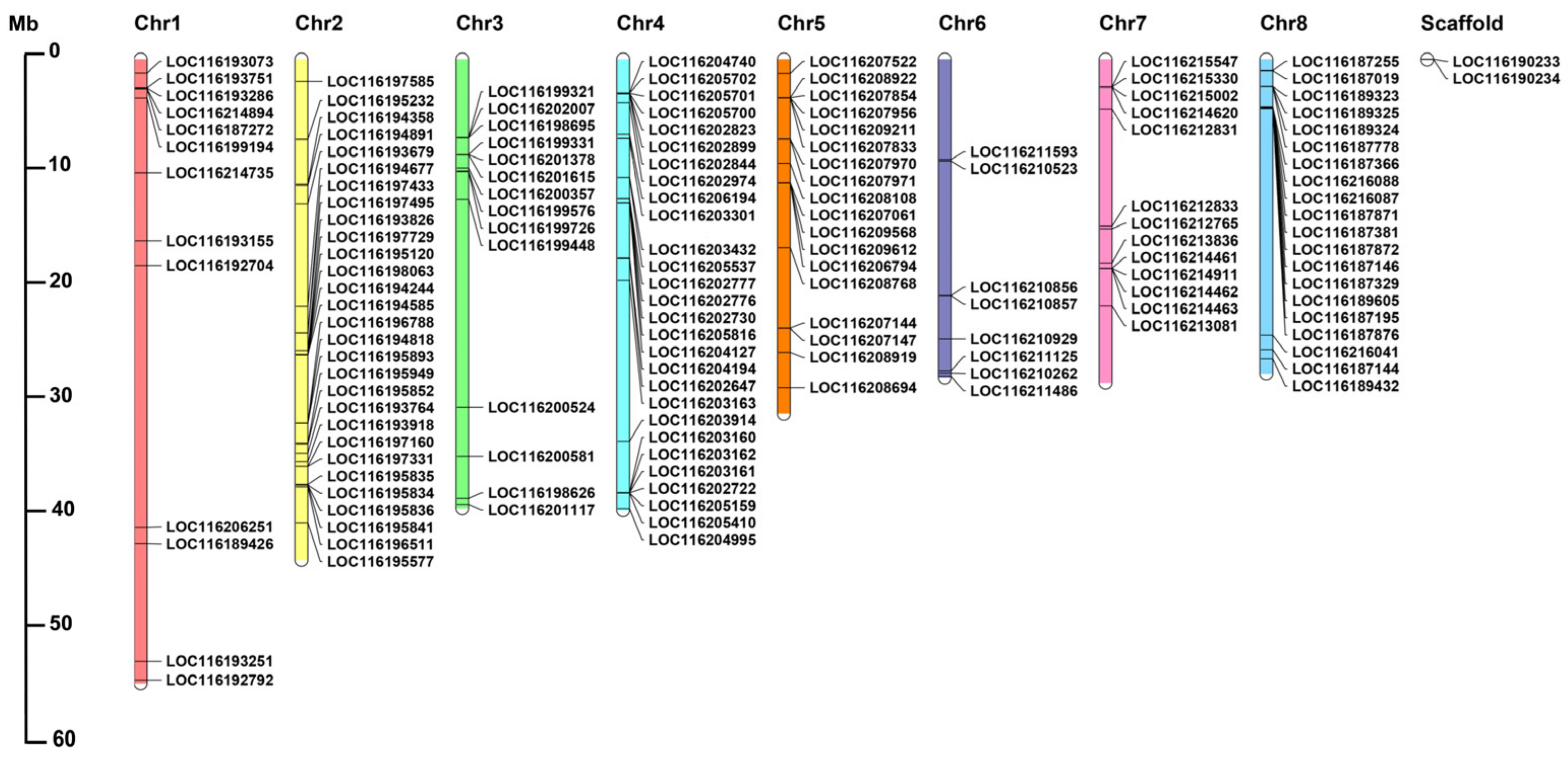
3.3. Chromosome Distribution and Gene Duplication
3.4. Conserved Motifs and Exon–Intron Organization of PgUGTs
3.5. Characterization of Cis-Acting Elements in the PgUGT Promoters
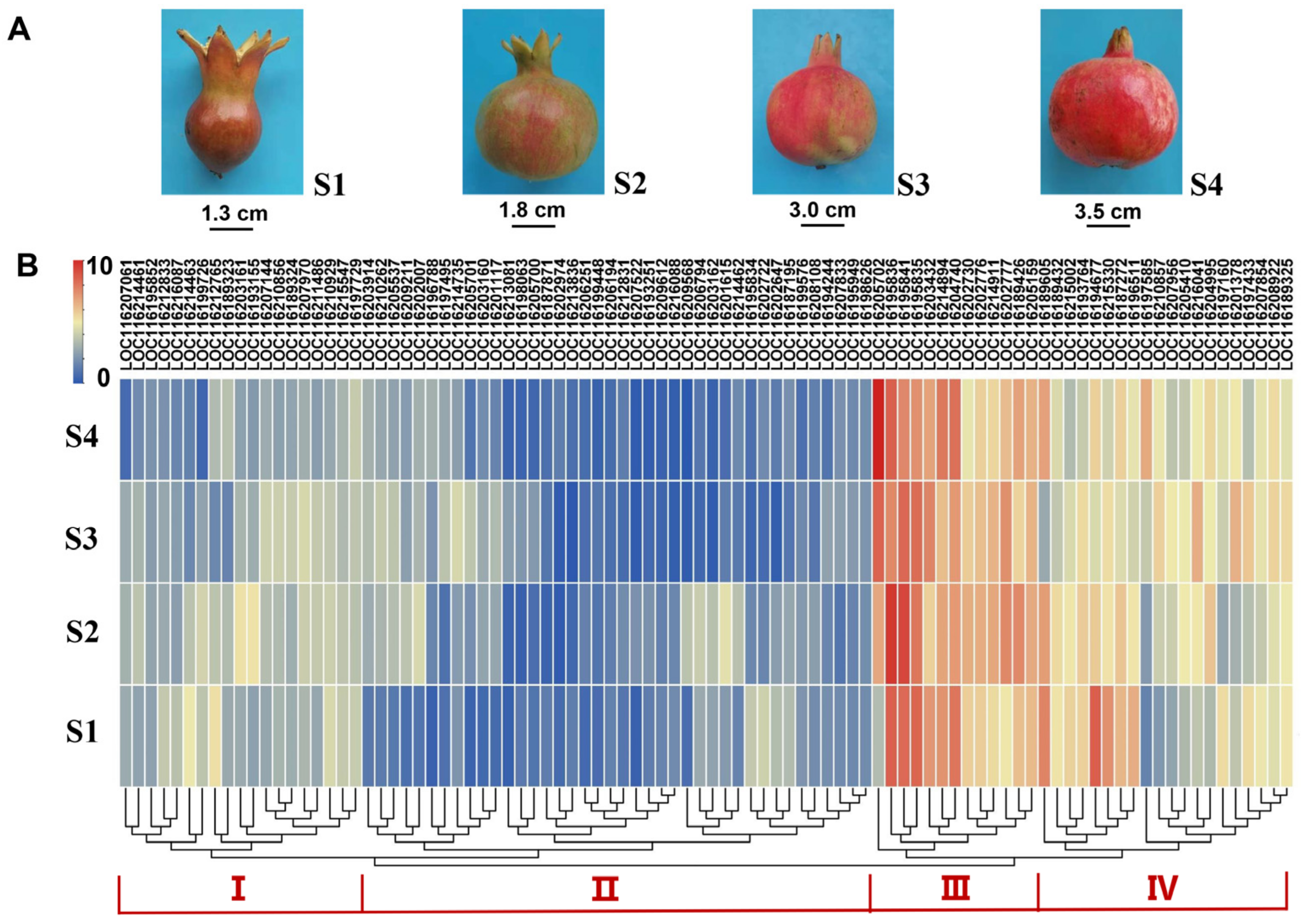
3.6. Temporal Expression Profiles of PgUGTs during Fruit Development
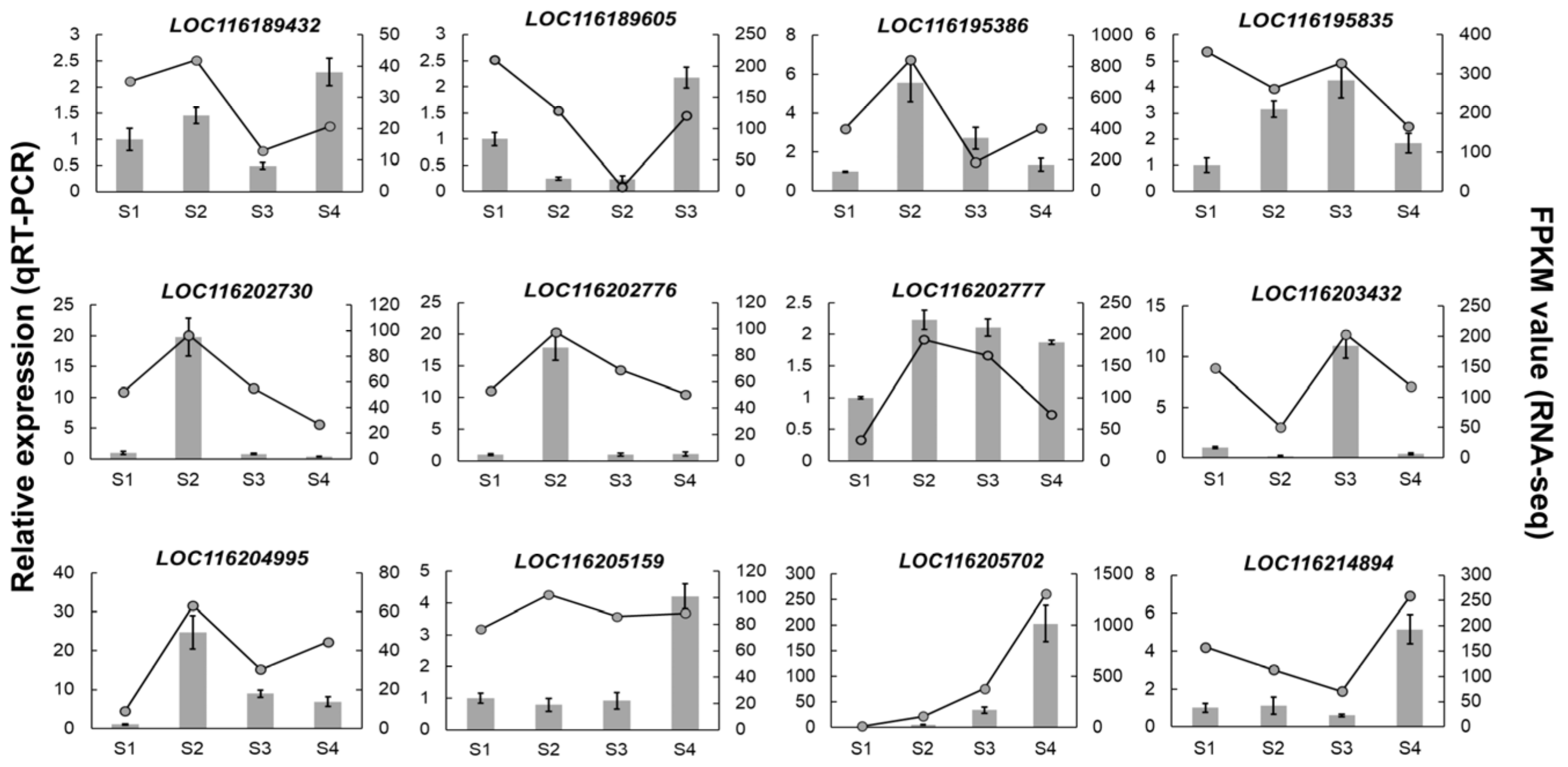
3.7. The Validation of PgUGT Expression with qRT-PCR
3.8. Functional Prediction of PgUGT Involved in Flavonoid Biosynthesis
4. Discussion
4.1. The Classification of PgUGTs Based on Phylogenetic Tree
4.2. Segmental and Tandem Duplication Contribute to the Expansion of PgUGT Family
4.3. PgUGT Transcription Analysis
4.4. Identification of PgUGTs Involved in Flavonoid Biosynthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, Z.; Fang, Y.; Zhang, T.; Fei, Z.; Han, F.; Liu, C.; Liu, M.; Xiao, W.; Zhang, W.; Wu, S.; et al. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2018, 16, 1363–1374. [Google Scholar] [PubMed]
- Ge, S.S.; Duo, L.; Wang, J.; Yang, J.F.; Li, Z.Y.; Tu, Y. A unique understanding of traditional medicine of pomegranate, Punica granatum L. and its current research status. J. Ethnopharmacol. 2021, 271, 113877. [Google Scholar] [PubMed]
- Ranjha, M.M.A.N.; Shafique, B.; Wang, L.; Irfan, S.; Safdar, M.N.; Murtaza, M.A.; Nadeem, M.; Mahmood, S.; Mueen-ud-Din, G.; Nadeem, H.R. A comprehensive review on phytochemistry, bioactivity and medicinal value of bioactive compounds of pomegranate (Punica granatum). Adv. Tradit. Med. 2023, 23, 37–57. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Fernandez-Lopez, J.; Perez-Alvarez, J.A. Pomegranate and its many functional components as related to human health: A review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 635–654. [Google Scholar]
- Zhao, X.; Yuan, Z. Anthocyanins from pomegranate (Punica granatum L.) and their role in antioxidant capacities in vitro. Chem. Biodivers. 2021, 18, e2100399. [Google Scholar] [PubMed]
- Paquette, S.; Moller, B.L.; Bak, S. On the origin of family 1 plant glycosyltransferases. Phytochemistry 2003, 62, 399–413. [Google Scholar] [CrossRef]
- Caputi, L.; Malnoy, M.; Goremykin, V.; Nikiforova, S.; Martens, S. A genome-wide phylogenetic reconstruction of family 1 UDP-glycosyltransferases revealed the expansion of the family during the adaptation of plants to life on land. Plant J. 2012, 69, 1030–1042. [Google Scholar] [CrossRef]
- Albesa-Jove, D.; Guerin, M.E. The conformational plasticity of glycosyltransferases. Curr. Opi. Struc. Biol. 2016, 40, 23–32. [Google Scholar] [CrossRef]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar] [CrossRef]
- Gachon, C.M.M.; Langlois-Meurinne, M.; Saindrenan, P. Plant secondary metabolism glycosyltransferases: The emerging functional analysis. Trends Plant Sci. 2005, 10, 542–549. [Google Scholar] [CrossRef]
- Vogt, T.; Jones, P. Glycosyltransferases in plant natural product synthesis: Characterization of a supergene family. Trends Plant Sci. 2000, 5, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.A.; Carle, R.; Kammerer, D.R. Identification and quantification of phenolic compounds from pomegranate (Punica granatum L.) peel, mesocarp, aril and differently produced juices by HPLC-DAD-ESI/MSn. Food Chem. 2011, 127, 807–821. [Google Scholar]
- Fourati, M.; Smaoui, S.; Ben Hlima, H.; Elhadef, K.; Ben Braiek, O.; Ennouri, K.; Mtibaa, A.C.; Mellouli, L. Bioactive compounds and pharmacological potential of pomegranate (Punica granatum) seeds—A review. Plant Food Hum. Nutr. 2020, 75, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Man, G.; Xu, L.; Wang, Y.; Liao, X.; Xu, Z. Profiling phenolic composition in pomegranate peel from nine selected cultivars using UHPLC-QTOF-MS and UPLC-QQQ-MS. Front Nutr. 2022, 8, 807447. [Google Scholar] [CrossRef] [PubMed]
- Zeghad, N.; Abassi, E.A.; Belkhiri, A.; Demeyer, K.; Heyden, Y.V. Phenolic compounds profile from Algerian pomegranate fruit extract (Punica granatum L.) by UPLC-DAD-ESI-MS. Chem. Afr. 2022, 5, 1295–1303. [Google Scholar]
- Wilson, A.E.; Wu, S.; Tian, L. PgUGT95B2 preferentially metabolizes flavones/flavonols and has evolved independently from flavone/flavonol UGTs identified in Arabidopsis thaliana. Phytochemistry 2019, 157, 184–193. [Google Scholar] [CrossRef]
- Ono, N.N.; Qin, X.; Wilson, A.E.; Li, G.; Tian, L. Two UGT84 family glycosyltransferases catalyze a critical reaction of hydrolyzable tannin biosynthesis in pomegranate (Punica granatum). PLoS ONE 2016, 11, e0156319. [Google Scholar] [CrossRef]
- Chang, L.; Wu, S.; Tian, L. Effective genome editing and identification of a regiospecific gallic acid 4-O-glycosyltransferase in pomegranate (Punica granatum L.). Hortic. Res. 2019, 6, 123. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Cavalcanti, A.; Chen, F.; Bouman, P.; Li, W. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yuan, Z.; Feng, L.; Fang, Y. Cloning and expression of anthocyanin biosynthetic genes in red and white pomegranate. J. Plant Res. 2015, 128, 687–696. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef]
- Akere, A.; Chen, S.H.; Liu, X.; Chen, Y.; Dantu, S.C.; Pandini, A.; Bhowmik, D. Structure-based enzyme engineering improves donor-substrate regognition of Arabidopsis thaliana glycosyltransferases. Bichem. J. 2020, 477, 2791–2805. [Google Scholar] [CrossRef]
- Rehman, H.M.; Nawaz, M.A.; Bao, L.; Shah, Z.H.; Lee, J.; Ahmad, M.Q.; Chung, G.; Yang, S.H. Genome-wide analysis of family-1 UDP-glycosyltransferases in soybean confirms their abundance and varied expression during seed development. J. Plant Physiol. 2016, 206, 87–97. [Google Scholar] [CrossRef]
- Wang, F.; Su, Y.; Chen, N.; Shen, S. Genome-wide analysis of the UGT gene family and identification of flavonids in Broussonetia papyrifera. Molecules 2021, 26, 3449. [Google Scholar] [CrossRef]
- Song, Z.; Niu, L.; Yang, Q.; Dong, B.; Wang, L.; Dong, M.; Fan, X.; Jian, Y.; Meng, D.; Fu, Y. Genome-wide identification and characterization of UGT family in pigeonpea (Cajanus cajan) and expression analysis in abiotic stress. Trees 2019, 33, 987–1002. [Google Scholar] [CrossRef]
- Cui, L.; Yao, S.; Dai, X.; Yin, Q.; Liu, Y.; Jiang, X.; Wu, Y.; Qian, Y.; Pang, Y.; Gao, L.; et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Bot. 2016, 67, 2285–2297. [Google Scholar] [CrossRef]
- Wu, B.; Liu, X.; Xu, K.; Zhang, B. Genome-wide characterization, evolution and expression profiling of UDP-glycosyltransferase family in pomelo (Citrus grandis) fruit. BMC Plant Biol. 2020, 20, 459. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Gu, J.; Luo, Y.; Wang, Y.; Pang, Y.; Shen, G.; Guo, B. Genome-wide analysis of UGT gene family identified key gene for the biosynthesis of bioactive flavonol glycosides in Epimedium pubescens Maxim. Syn. Syst. Biotechnol. 2022, 7, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Pang, C.; Fan, S.; Song, M.; Yu, J.; Wei, H.; Ma, Q.; Li, L.; Zhang, C.; Yu, S. Genome-wide analysis of the family 1 glycosyltransferases in cotton. Mol. Genet. Genom. 2015, 290, 1805–1818. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, L.; Huang, H.; Zhang, Y.; Wang, J.; Lu, X.; Wang, S.; Wang, D.; Chen, X.; Chen, C.; et al. Genome-wide identifcation, evolution and function analysis of UGTs superfamily in cotton. Front. Mol. Biosci. 2022, 9, 965403. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Tsukamota, C.; Ishimoto, M. Reconstruction of the evolutionary histories of UGT gene sumperfamily in Legumes clarifies the functional divergence of duplicates in specialized metabolism. Int. J. Mol. Sci. 2020, 21, 1855. [Google Scholar] [CrossRef]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Kadoo, N.Y.; Gupta, V.S. Phylogenomic analysis of UDP glycosyltransferase 1 multigene family in Linum usitatissimum identified genes with varied expression patterns. BMC Genom. 2012, 13, 175. [Google Scholar] [CrossRef]
- Zhou, K.; Hu, L.; Li, P.; Gong, X.; Ma, F. Genome-wide identification of glycosyltransferases converting phloretin to phloridzin in Malus species. Plant Sci. 2017, 265, 131–145. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.; Zhang, L.; Shu, J.; Court, M.H.; Sun, Z.; Jiang, L.; Zheng, C.; Shu, H.; Ji, L.; et al. Genome-wide analysis of the apple family 1 glycosyltransferases identified a flavonoid-modifying UGT, MdUGT83L3, which is targeted by MdMYB88 and contributes to stress adaptation. Plant Sci. 2022, 321, 111314. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Dai, J.; Chen, Z.; Tie, W.; Yan, Y.; Yang, H.; Zeng, J.; Hu, W. Comprehensive analysis and expression profiles of cassava UDP-glycosyltransferases (UGT) family reveal their involvement in development and stress responses in cassava. Genomics 2021, 113, 3415–3429. [Google Scholar] [CrossRef]
- Ao, B.; Han, Y.; Wang, S.; Wu, F.; Zhang, J. Genome-Wide analysis and profile of UDP-glycosyltransferases family in alfalfa (Medicago sativa L.) under drought stress. Int. J. Mol. Sci. 2022, 23, 7243. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Yan, Q.; Wu, F.; Wang, Y.; Wang, S.; Zong, X.; Zhou, X.; Zhang, J. Genome-wide analysis of the UDP-glycosyltransferase family reveals its role in coumarin biosynthesis and abiotic stress in Melilotus albus. Int. J. Mol. Sci. 2021, 22, 10826. [Google Scholar] [CrossRef]
- Ren, C.; Guo, Y.; Xie, L.; Zhao, Z.; Xing, Z.; Cao, Y.; Liu, Y.; Lin, J.; Grierson, D.; Zhang, B.; et al. Identification of UDP-rhamnosyltransferases and UDP-galactosyltransferase involved in flavonol glycosylation in Morella rubra. Hortic. Res. 2022, 9, uhac138. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, X.; Lu, M.; Chen, J.; Shi, T. Gene expression and evolution of family-1 UDPglycosyltransferases—Insights from an aquatic flowering plant (sacred lotus). Aquat. Bot. 2020, 166, 103270. [Google Scholar] [CrossRef]
- Dong, L.; Tang, Z.; Yang, T.; Hao, F.; Deng, X. Genome-wide analysis of UGT genes in Petunia and identification of PhUGT51 involved in the regulation of salt resistance. Plants 2022, 11, 2434. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Yan, X.; Zhang, Q. Comparative genomic and transcriptomic analyses of family-1 UDP glycosyltransferase in Prunus Mume. Int. J. Mol. Sci. 2018, 19, 3382. [Google Scholar] [CrossRef]
- Wu, B.; Gao, L.; Gao, J.; Xu, Y.; Liu, H.; Cao, X.; Zhang, B.; Chen, K. Genome-wide identification, expression patterns, and functional analysis of UDP glycosyltransferase family in peach (Prunus persica L. Batsch). Front. Plant Sci. 2017, 8, 389. [Google Scholar] [CrossRef]
- Li, G.; Li, J.; Qin, G.; Liu, C.; Liu, X.; Cao, Z.; Jia, B.; Zhang, H. Characterization and expression analysis of the UDP glycosyltransferase family in pomegranate (Punica granatum L.). Horticulturae 2023, 9, 119. [Google Scholar] [CrossRef]
- Cheng, X.; Muhammad, A.; Li, G.; Zhang, J.; Cheng, J.; Qiu, J.; Jiang, T.; Jin, Q.; Cai, Y.; Lin, Y. Family-1 UDP glycosyltransferases in pear (Pyrus bretschneideri): Molecular identification, phylogenomic characterization and expression profiling during stone cell formation. Mol. Biol. Rep. 2019, 46, 2152–2175. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Ahmad, D.; Zhang, X.; Zhang, Y.; Wu, L.; Jiang, P.; Ma, H. Genome-wide analysis of family-1 UDP glycosyltransferases (UGT) and identification of UGT genes for FHB resistance in wheat (Triticum aestivum L.). BMC Plant Biol. 2018, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Mu, H.; Xu, G.; Wang, Y.; Li, S.; Wang, L. Genome-wide analysis and functional characterization of the UDP-glycosyltransferase family in grapes. Horticulturae 2021, 7, 204. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.; Wang, Y.; Dong, R.; Yu, H.; Hou, B. Genome-wide identification and phylogenetic analysis of family-1 UDP-glycosyltransferases in maize (Zea mays). Planta 2014, 239, 1256–1279. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Wu, Z.; Yao, W.; Cao, S. The pomegranate (Punica granatum L.) draft genome dissects genetic divergence between soft- and hard-seeded cultivars. Plant Biotechnol. J. 2019, 18, 955–968. [Google Scholar]
- Ross, J.; Li, Y.; Lim, E.; Bowles, D.J. Higher plant glycosyltransferases. Genome Biol. 2001, 2, REVIEWS3004. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S. Evolution of gene dulication in plants. Plant Physiol. 2016, 171, 1194–2316. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Zhao, X.; Shen, Y.; Yan, M.; Yuan, Z. Flavonoid profiles in peels and arils of pomegranate cutlivars. J. Food Meas. Charact. 2022, 16, 880–890. [Google Scholar] [CrossRef]
- Sharma, R.; Rawat, V.; Suresh, C.G. Genome-wide identfication and tissue-specific expression analysis of UDP-glycosyltransferases genes confirm their abundance in Cicer arietinum (Chickea). PLoS ONE 2014, 9, e109715. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lin, C.; Ma, X.; Tan, Y.; Wang, J.; Zeng, M. Functional characterization of a flavonoid glycosyltransferase in sweet orange (Citrus sinensis). Front. Plant Sci. 2018, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.E.; Tian, L. Phylogenomic analysis of UDP-dependent glycosyltransferases provides insights into the evolutionary landscape of glycosylation in plant metabolism. Plant J. 2019, 100, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, A.; Horikawa, M.; Fukui, Y.; Fukuchi-Mizutani, M.; Iuchi-Okada, A.; Ishiguro, M.; Kiso, Y.; Nakayama, T.; Ono, E. Local differentiation of sugar donor specificity of flavonoid glycosyltransferase in Lamiales. Plant Cell 2009, 21, 1556–1572. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Saito, K. Function, structure, and evolution of flavonoid glycosyltransferases in Plants. In Recent Advances in Polyphenol Research; Romani, A., Lattanzio, V., Quideau, S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 61–82. [Google Scholar]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S.; et al. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1875. [Google Scholar] [CrossRef]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitajima, M.; et al. Functional genomics by integrated analysis of metabolome and transcriptome of Arabidopsis plants over-expressing an MYB transcription factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Nakabayashi, R.; Sugawara, S.; Tohge, T.; Ito, T.; Koyanagi, M.; Kitajima, M.; Takayama, H.; Saito, K. A flavonoid 3-O-glucoside:2″-O-glucosyltransferase responsible for terminal modification of pollen-specific flavonols in Arabidopsis thaliana. Plant J. 2014, 79, 769–782. [Google Scholar] [CrossRef]
- Osmani, S.A.; Bak, S.; Møller, B.L. Substrate specificity of plant UDP-dependent glycosyltransferases predicted from crystal structures and homology modeling. Phytochemistry 2009, 70, 325–347. [Google Scholar] [CrossRef]
- Modolo, L.V.; Blount, J.W.; Achnine, L.; Naoumkina, M.; Wang, X. A functional genomics approach to (iso)flavonoid glycosylation in the model legume Medicago truncatula. Plant Mol. Biol. 2007, 64, 499–518. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Total | A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Arabidopsis thaliana | 122 | 19 | 4 | 4 | 13 | 25 | 4 | 7 | 21 | 1 | 2 | 2 | 18 | 1 | 1 | 0 | 0 | 0 | 0 | [29] |
| 2 | Brassica napus | 251 | 17 | 10 | 10 | 36 | 48 | 2 | 14 | 35 | 2 | 3 | 6 | 61 | 4 | 3 | 0 | 0 | 0 | 0 | [30] |
| 3 | Brassica oleraca | 154 | 15 | 7 | 4 | 23 | 32 | 0 | 8 | 23 | 1 | 2 | 3 | 32 | 2 | 2 | 0 | 0 | 0 | 0 | [30] |
| 4 | Brassica rapa | 140 | 12 | 4 | 4 | 24 | 31 | 1 | 9 | 18 | 1 | 3 | 3 | 26 | 2 | 2 | 0 | 0 | 0 | 0 | [30] |
| 5 | Broussonetia papyrifera | 155 | 13 | 4 | 2 | 20 | 46 | 6 | 19 | 6 | 3 | 6 | 7 | 15 | 5 | 1 | 2 | 0 | 0 | 0 | [31] |
| 6 | Cajanus cajan | 120 | 2 | 2 | 1 | 36 | 33 | 0 | 9 | 0 | 5 | 0 | 0 | 12 | 2 | 0 | 6 | 12 | 0 | 0 | [32] |
| 7 | Camellia sinensis | 132 | 15 | 5 | 2 | 20 | 23 | 2 | 13 | 2 | 2 | 2 | 1 | 27 | 3 | 0 | 6 | 6 | 0 | 3 | [33] |
| 8 | Citrus grandis | 145 | 17 | 3 | 1 | 18 | 25 | 2 | 9 | 17 | 17 | 3 | 2 | 12 | 7 | 1 | 4 | 7 | 0 | 0 | [34] |
| 9 | Cucumis sativus | 85 | 10 | 1 | 2 | 12 | 13 | 0 | 11 | 5 | 0 | 2 | 1 | 17 | 2 | 1 | 3 | 5 | 0 | 0 | [7] |
| 10 | Epimedium pubescens | 339 | 105 | 9 | 23 | 24 | 49 | 5 | 15 | 17 | 11 | 6 | 6 | 49 | 2 | 1 | 15 | 0 | 2 | 0 | [35] |
| 11 | Gossypium arboreum | 146 | 20 | 8 | 3 | 21 | 28 | 7 | 13 | 7 | 7 | 4 | 1 | 13 | 4 | 1 | 1 | 8 | 0 | 0 | [36] |
| 12 | Gossypium arboreum | 143 | 19 | 7 | 1 | 21 | 30 | 5 | 9 | 8 | 7 | 4 | 1 | 17 | 3 | 1 | 1 | 8 | 0 | 1 | [37] |
| 13 | Gossypium barbadense | 220 | 26 | 12 | 2 | 34 | 37 | 10 | 14 | 13 | 14 | 5 | 2 | 29 | 5 | 1 | 2 | 10 | 0 | 4 | [37] |
| 14 | Gossypium hirsutum | 196 | 17 | 12 | 0 | 36 | 38 | 8 | 20 | 16 | 10 | 4 | 2 | 18 | 2 | 2 | 1 | 10 | 0 | 0 | [36] |
| 15 | Gossypium hirsutum | 220 | 27 | 14 | 2 | 29 | 41 | 12 | 14 | 15 | 11 | 5 | 2 | 27 | 4 | 2 | 2 | 9 | 0 | 4 | [37] |
| 16 | Gossypium raimondii | 152 | 15 | 9 | 2 | 20 | 30 | 8 | 5 | 8 | 7 | 4 | 1 | 15 | 4 | 1 | 1 | 12 | 0 | 0 | [36] |
| 17 | Gossypium raimondii | 149 | 13 | 9 | 1 | 21 | 29 | 8 | 5 | 8 | 7 | 4 | 1 | 20 | 3 | 1 | 1 | 12 | 0 | 6 | [37] |
| 18 | Glycine max | 182 | 25 | 3 | 1 | 43 | 36 | 1 | 15 | 3 | 18 | 3 | 2 | 19 | 4 | 1 | 5 | 3 | 0 | 0 | [7] |
| 19 | Glycine max | 149 | 5 | 1 | 2 | 38 | 46 | 6 | 16 | 2 | 4 | 0 | 0 | 18 | 5 | 0 | 6 | 0 | 0 | 0 | [30] |
| 20 | Glycine max | 208 | 21 | 3 | 0 | 46 | 52 | 8 | 16 | 3 | 17 | 7 | 0 | 19 | 5 | 1 | 6 | 4 | 0 | 0 | [38] |
| 21 | Linum usitatissimum | 137 | 16 | 5 | 6 | 21 | 22 | 1 | 19 | 6 | 9 | 4 | 5 | 19 | 3 | 1 | 0 | 0 | 0 | 0 | [39] |
| 22 | Lotus japonicus | 94 | 9 | 3 | 0 | 25 | 22 | 2 | 9 | 1 | 2 | 1 | 0 | 10 | 1 | 1 | 6 | 1 | 0 | 1 | [38] |
| 23 | Malus × domestica | 241 | 33 | 4 | 7 | 13 | 55 | 6 | 40 | 14 | 11 | 12 | 6 | 16 | 13 | 1 | 5 | 5 | 0 | 0 | [7] |
| 24 | Malus × domestica | 237 | 34 | 0 | 8 | 11 | 50 | 3 | 50 | 17 | 12 | 13 | 7 | 15 | 7 | 1 | 2 | 7 | 0 | 0 | [40] |
| 25 | Malus × domestica | 229 | 30 | 3 | 6 | 16 | 42 | 6 | 37 | 21 | 8 | 10 | 4 | 21 | 7 | 2 | 2 | 5 | 4 | 4 | [41] |
| 26 | Manihot esculenta | 121 | 14 | 4 | 3 | 18 | 18 | 3 | 12 | 7 | 11 | 2 | 3 | 22 | 3 | 1 | 0 | 0 | 0 | 0 | [42] |
| 27 | Medicago sativa | 409 | 0 | 4 | 2 | 100 | 62 | 6 | 0 | 13 | 134 | 0 | 10 | 43 | 9 | 0 | 10 | 14 | 0 | 2 | [43] |
| 28 | Medicago truncatula | 243 | 28 | 4 | 0 | 55 | 55 | 2 | 39 | 3 | 5 | 9 | 0 | 33 | 2 | 1 | 3 | 3 | 0 | 1 | [38] |
| 29 | Melilotus albus | 189 | 21 | 3 | 0 | 39 | 52 | 0 | 33 | 4 | 1 | 2 | 0 | 24 | 2 | 1 | 5 | 2 | 0 | 0 | [44] |
| 30 | Morella rubra | 152 | 8 | 8 | 5 | 9 | 33 | 4 | 26 | 11 | 0 | 8 | 8 | 24 | 4 | 0 | 4 | 0 | 0 | 0 | [45] |
| 31 | Nelumbo nucifera | 108 | 11 | 7 | 5 | 6 | 20 | 6 | 10 | 3 | 4 | 2 | 1 | 16 | 8 | 1 | 4 | 4 | 0 | 0 | [46] |
| 32 | Oryza sativa | 180 | 14 | 9 | 8 | 26 | 38 | 0 | 20 | 7 | 9 | 3 | 1 | 23 | 5 | 2 | 6 | 9 | 0 | 0 | [7] |
| 33 | Petunia hybrida | 129 | 27 | 1 | 2 | 12 | 20 | 2 | 8 | 3 | 5 | 6 | 1 | 10 | 5 | 1 | 22 | 4 | 0 | 0 | [47] |
| 34 | Phaseolus vulgaris | 168 | 19 | 3 | 2 | 33 | 33 | 5 | 18 | 3 | 15 | 3 | 0 | 17 | 4 | 1 | 6 | 5 | 0 | 1 | [38] |
| 35 | Populus trichocarpa | 178 | 12 | 2 | 6 | 14 | 49 | 0 | 42 | 5 | 5 | 6 | 2 | 23 | 6 | 1 | 3 | 2 | 0 | 0 | [7] |
| 36 | Prunus mume | 130 | 16 | 2 | 3 | 17 | 23 | 3 | 18 | 10 | 4 | ? | 8 | 17 | 3 | ? | 0 | 0 | 0 | 0 | [48] |
| 37 | Prunus persica | 168 | 10 | 2 | 4 | 19 | 29 | 4 | 34 | 9 | 5 | 7 | 7 | 18 | 14 | 1 | 1 | 4 | 0 | 0 | [49] |
| 38 | Punica granatum | 120 | 13 | 7 | 3 | 12 | 23 | 6 | 21 | 7 | 2 | 2 | 0 | 13 | 2 | 1 | 0 | 4 | 0 | 4 | [50] |
| 39 | Punica granatum | 145 | 14 | 7 | 3 | 13 | 28 | 8 | 27 | 7 | 2 | 1 | 2 | 19 | 3 | 2 | 1 | 4 | 0 | 4 | This study |
| 40 | Pyrus bretschneideri | 139 | 5 | 4 | 2 | 8 | 31 | 6 | 33 | 10 | 10 | 2 | 9 | 10 | 3 | 3 | 3 | 0 | 0 | 0 | [51] |
| 41 | Sorglum bicolor | 180 | 10 | 4 | 6 | 24 | 50 | 0 | 17 | 12 | 8 | 3 | 1 | 26 | 6 | 3 | 8 | 2 | 0 | 0 | [7] |
| 42 | Trifolium pratense | 121 | 11 | 3 | 0 | 29 | 39 | 1 | 13 | 3 | 1 | 2 | 0 | 12 | 1 | 0 | 2 | 3 | 0 | 1 | [38] |
| 43 | Triticum aestivum | 179 | 22 | 3 | 2 | 17 | 37 | 2 | 4 | 5 | 7 | 5 | 0 | 19 | 3 | 1 | 3 | 13 | 36 | 0 | [52] |
| 44 | Vitis vinifera | 228 | 25 | 4 | 6 | 9 | 45 | 8 | 29 | 7 | 13 | 7 | 2 | 33 | 5 | 1 | 3 | 0 | 0 | 0 | [53] |
| 45 | Vitis vinifera | 181 | 23 | 3 | 4 | 8 | 46 | 5 | 15 | 7 | 14 | 4 | 2 | 31 | 5 | 1 | 2 | 11 | 0 | 0 | [7] |
| 46 | Zea mays | 147 | 8 | 3 | 5 | 18 | 34 | 2 | 12 | 9 | 9 | 3 | 1 | 23 | 3 | 4 | 5 | 1 | 7 | 0 | [54] |
| Function | Promoter Name | Promoter Annotation | Total Number |
|---|---|---|---|
| Hormone response | ABRE | cis-acting element involved in abscisic acid responsiveness | 393 |
| AuxRR-core | cis-acting regulatory element involved in auxin responsiveness | 34 | |
| CGTCA-motif | cis-acting regulatory element involved in MeJA responsiveness | 312 | |
| GARE-motif | gibberellin-responsive element | 19 | |
| P-box | gibberellin-responsive element | 50 | |
| TATC-box | cis-acting element involved in gibberellin responsiveness | 30 | |
| TCA-element | cis-acting element involved in salicylic acid responsiveness | 70 | |
| TGA-box | part of an auxin-responsive element | 7 | |
| TGA-element | auxin-responsive element | 66 | |
| TGACG-motif | cis-acting regulatory element involved in MeJA responsiveness | 313 | |
| Stress response | ARE | cis-acting regulatory element essential for anaerobic induction | 222 |
| GC-motif | enhancer-like element involved in anoxic-specific inducibility | 52 | |
| LTR | cis-acting element involved in low-temperature responsiveness | 137 | |
| MBS | MYB binding site involved in drought-inducibility | 81 | |
| TC-rich repeats | cis-acting element involved in defense and stress responsiveness | 63 | |
| WUN-motif | wound-responsive element | 13 | |
| Developmental response | CAT-box | cis-acting regulatory element related to meristem expression | 84 |
| circadian | cis-acting regulatory element involved in circadian control | 34 | |
| GCN4_motif | cis-regulatory element involved in endosperm expression | 28 | |
| HD-Zip 1 | element involved in differentiation of palisade mesophyll cells | 15 | |
| O2-site | cis-acting regulatory element involved in zein metabolism regulation | 73 | |
| MBSI | MYB binding site involved in flavonoid biosynthetic gene regulation | 11 | |
| RY-element | cis-acting regulatory element involved in seed-specific regulation | 12 | |
| MSA-like | cis-acting element involved in cell cycle regulation | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Feng, Y.; Ke, D.; Teng, Y.; Chen, Y.; Langjia, R. Molecular Identification and Characterization of UDP-glycosyltransferase (UGT) Multigene Family in Pomegranate. Horticulturae 2023, 9, 540. https://doi.org/10.3390/horticulturae9050540
Zhao X, Feng Y, Ke D, Teng Y, Chen Y, Langjia R. Molecular Identification and Characterization of UDP-glycosyltransferase (UGT) Multigene Family in Pomegranate. Horticulturae. 2023; 9(5):540. https://doi.org/10.3390/horticulturae9050540
Chicago/Turabian StyleZhao, Xueqing, Yingyi Feng, Ding Ke, Yingfen Teng, Ying Chen, and Renzeng Langjia. 2023. "Molecular Identification and Characterization of UDP-glycosyltransferase (UGT) Multigene Family in Pomegranate" Horticulturae 9, no. 5: 540. https://doi.org/10.3390/horticulturae9050540
APA StyleZhao, X., Feng, Y., Ke, D., Teng, Y., Chen, Y., & Langjia, R. (2023). Molecular Identification and Characterization of UDP-glycosyltransferase (UGT) Multigene Family in Pomegranate. Horticulturae, 9(5), 540. https://doi.org/10.3390/horticulturae9050540