
Optimized Regeneration of Petunia Protoplast and Its Association with Tissue Identity Regulators


Abstract
1. Introduction
2. Results and Discussion
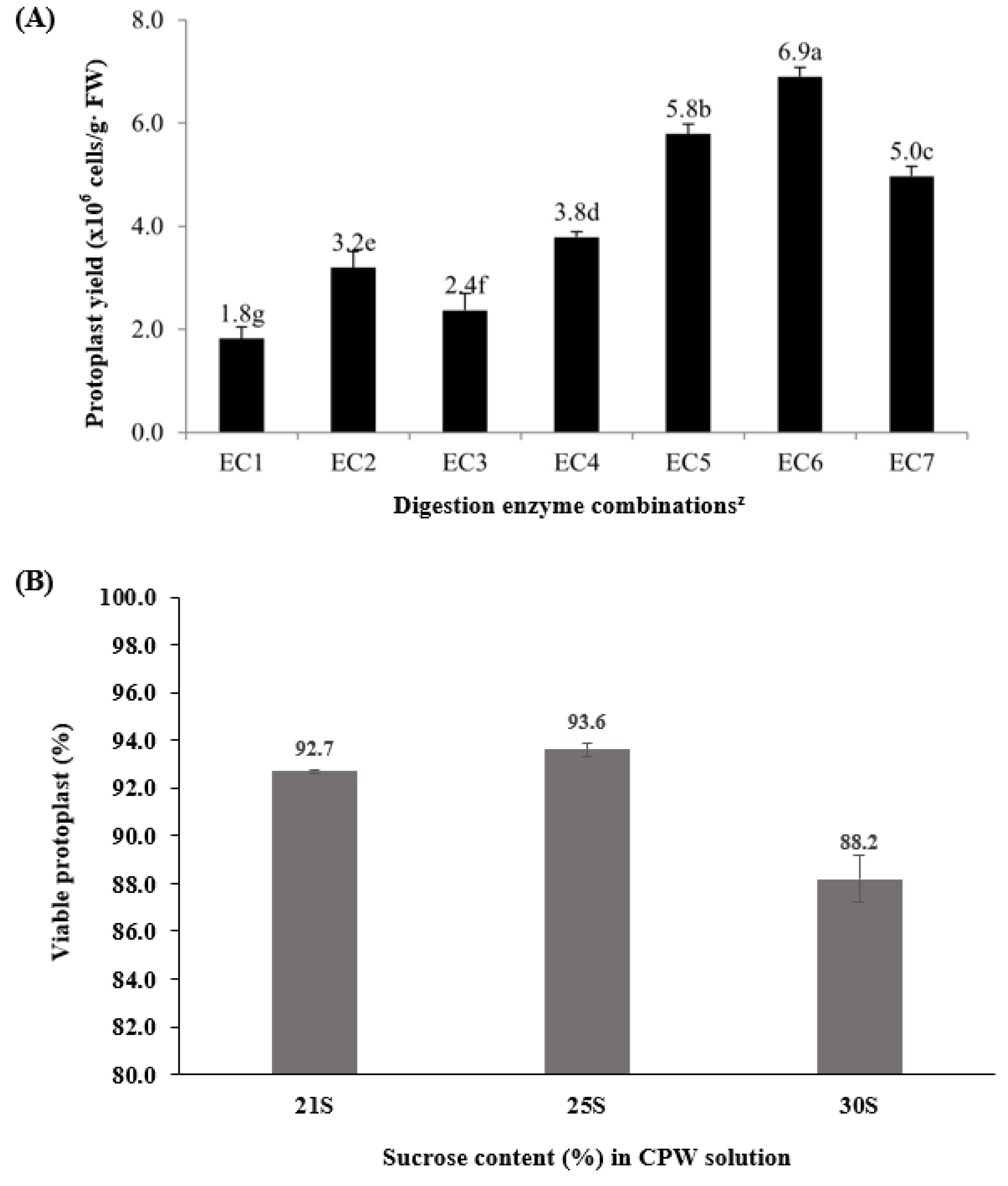
2.1. Enzyme Combinations for Protoplast Isolation
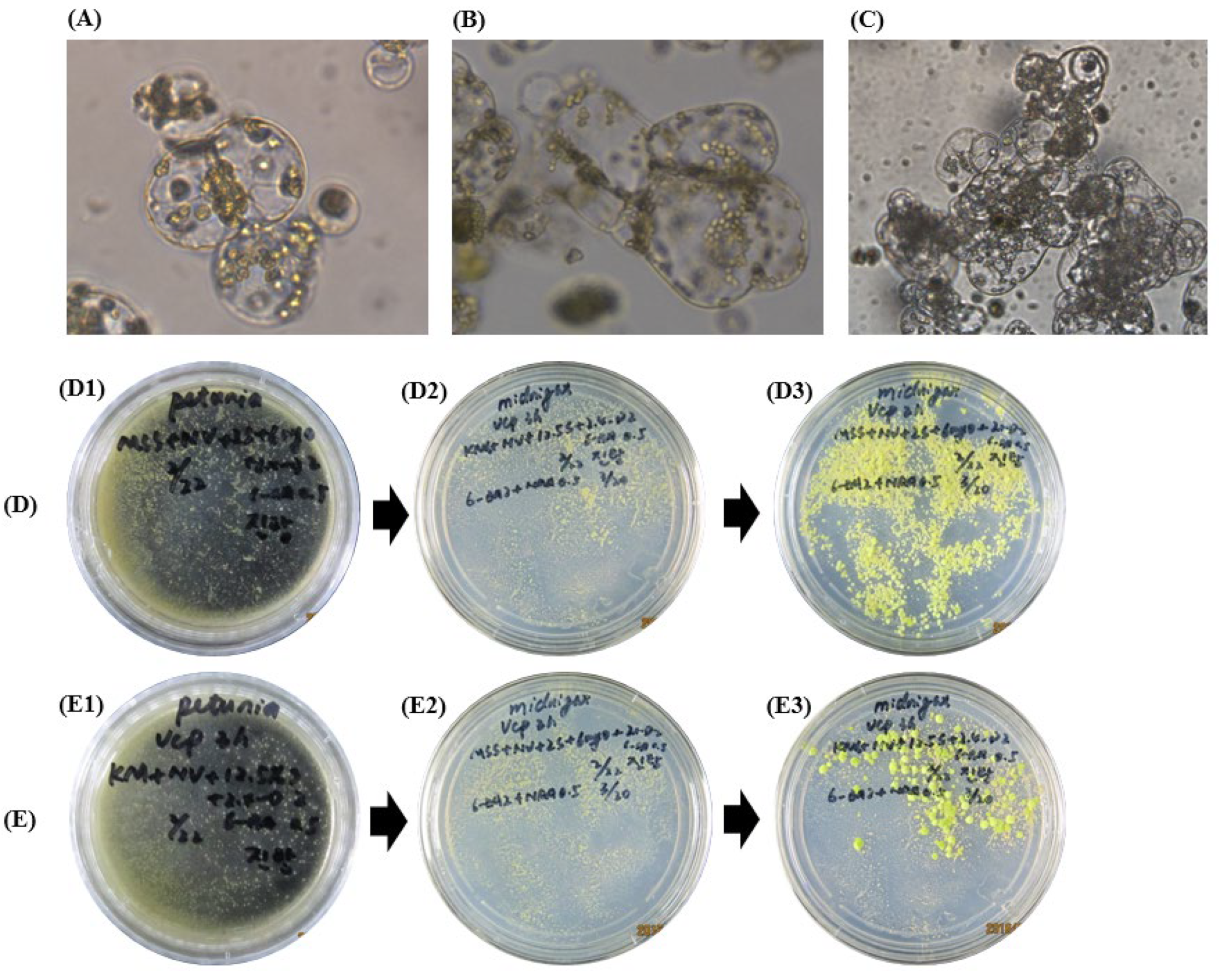
2.2. Protoplast Purification and Protoplast Viability
2.3. Different Culture Media for Colony and Calli Induction
2.4. Hormones for Shoot and Root Induction
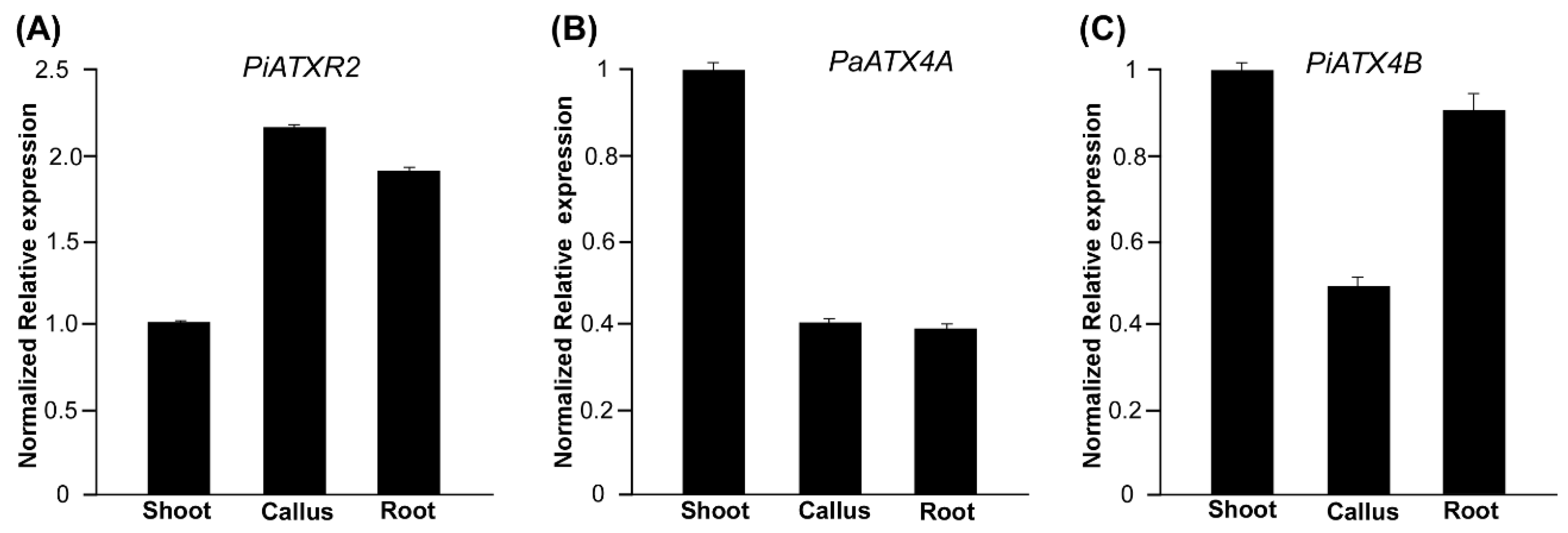
2.5. Epigenetic Regulation of Callus, Shoot, and Root Development in Petunia
3. Materials and Methods
3.1. Plant Material
3.2. Enzyme Digestion and Purification of Protoplasts
3.3. Induction of Colony and Callus Formation
3.4. Induction of Root and Shoot Generation
3.5. In Silico Identification of Candidate Genes and Estimation of mRNA Expression Using qRT-PCR
3.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Valente, P.; Tao, W.H.; Verbelen, J.P. Auxins and cytokinins control DNA endoreduplication and deduplication in single cells of tobacco. Plant Sci. 1998, 134, 207–215. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Sheen, J. Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant Physiol. 2001, 127, 1466–1475. [Google Scholar] [CrossRef]
- Meyer, L.; Serek, M.; Winkelmann, T. Protoplast isolation and plant regeneration of different genotypes of Petunia and Calibrachoa. Plant Cell Tiss. Org. Cult. 2009, 99, 27–34. [Google Scholar] [CrossRef]
- Oh, M.H.; Kim, S.G. Plant-regeneration from petal protoplast culture of Petunia hybrida. Plant Cell Tiss. Org. Cult. 1994, 36, 275–283. [Google Scholar] [CrossRef]
- Bona, C.; Gould, J.; Miller, J.C.; Stelly, D.M.; Louzada, E.S. In vitro regeneration of somatic symmetric and asymmetric hybrid citrus plantlets produced via protoplast fusion. Hortscience 2008, 43, 1206. [Google Scholar]
- Yu, J.; Tu, L.; Subburaj, S.; Bae, S.; Lee, G.J. Simultaneous targeting of duplicated genes in Petunia protoplasts for flower color modification via CRISPR-Cas9 ribonucleoproteins. Plant Cell Rep. 2021, 40, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Fordlogan, J.; Sink, K.C. Plantlet Regeneration from Protoplasts of Petunia alpicola. Hortscience 1988, 23, 393–395. [Google Scholar] [CrossRef]
- Sangthong, R.; Chin, D.P.; Hayashi, M.; Supaibulwatana, K.; Mii, M. Direct isolation of female germ units from ovules of Petunia hybrida by enzymatic treatment without releasing somatic protoplasts from ovular tissue. Plant Biotechnol. 2009, 26, 369–375. [Google Scholar] [CrossRef]
- Subburaj, S.; Chung, S.J.; Lee, C.; Ryu, S.M.; Kim, D.H.; Kim, J.S.; Bae, S.; Lee, G.J. Site-directed mutagenesis in Petunia x hybrida protoplast system using direct delivery of purified recombinant Cas9 ribonucleoproteins. Plant Cell Rep. 2016, 35, 1535–1544. [Google Scholar] [CrossRef]
- Oberwalder, B.; Schilde-Rentschler, L.; Ruoss, B.; Wittemann, S.; Ninnemann, H. Asymmetric protoplast fusions between wild species and breeding lines of potato—Effect of recipients and genome stability. Theor. Appl. Genet. 1998, 97, 1347–1354. [Google Scholar] [CrossRef]
- Jeong, Y.Y.; Lee, H.Y.; Kim, S.W.; Noh, Y.S.; Seo, P.J. Optimization of protoplast regeneration in the model plant Arabidopsis thaliana. Plant Methods 2021, 17, 21. [Google Scholar] [CrossRef]
- Lee, K.; Park, O.S.; Seo, P.J. Arabidopsis ATXR2 deposits H3K36me3 at the promoters of LBD genes to facilitate cellular dedifferentiation. Sci. Signal. 2017, 10, eaan0316. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, O.S.; Seo, P.J. ATXR2 as a core regulator of de novo root organogenesis. Plant Signal. Behav. 2018, 13, e1449543. [Google Scholar] [CrossRef]
- Lee, K.; Park, O.S.; Choi, C.Y.; Seo, P.J. ARABIDOPSIS TRITHORAX 4 Facilitates Shoot Identity Establishment during the Plant Regeneration Process. Plant Cell Physiol. 2019, 60, 826–834. [Google Scholar] [CrossRef]
- Yang, X.; Ma, D.-D.; Jiang, F.-S.; Chen, N.-P.; Ding, B.; Jin, L.-X.; Qian, C.-D.; Ding, Z.-S. Protoplasts isolation, purification and plant regeneration of Pinellia cordata. China J. Chin. Mater. Med. 2014, 39, 4211–4215. [Google Scholar]
- Zhang, Y.; Miao, J.; Xie, M.; Tian, S.; Dong, M.; Jiang, G. Isolation and purification of protoplast cell from the leaves of Torreya nucifera. Biotechnol. Bulletin. 2013, 11, 75. [Google Scholar]
- Cui, J.; Mackenzie, K.K.; Eeckhaut, T.; Muller, R.; Lutken, H. Protoplast isolation and culture from Kalanchoe species: Optimization of plant growth regulator concentration for efficient callus production. Plant Cell Tissue Org. Cult. 2019, 138, 287–297. [Google Scholar] [CrossRef]
- Klimek-Chodacka, M.; Kadluczka, D.; Lukasiewicz, A.; Malec-Pala, A.; Baranski, R.; Grzebelus, E. Effective callus induction and plant regeneration in callus and protoplast cultures of Nigella damascena L. Plant Cell Tissue Org. Cult. 2020, 143, 693–707. [Google Scholar] [CrossRef]
- Adedeji, O.S.; Naing, A.H.; Kim, C.K. Protoplast isolation and shoot regeneration from protoplast-derived calli of Chrysanthemum cv. White ND. Plant Cell Tissue Org. Plant Cell 2020, 141, 571–581. [Google Scholar] [CrossRef]
- Inokuma, C.; Sugiura, K.; Cho, C.; Okawara, R.; Kaneko, S. Plant regeneration from protoplasts of Japanese lawngrass. Plant Cell Rep. 1996, 15, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Mii, M.; Yuzawa, Y.; Suetomi, H.; Motegi, T.; Godo, T. Fertile Plant-Regeneration from Protoplasts of a seed-propagated cultivar of Lilium X Formolongi by utilizing meristematic nodular cell clumps. Plant Sci. 1994, 100, 221–226. [Google Scholar] [CrossRef]
- Sihachakr, D.; Ducreux, G. Plant-regeneration from protoplast culture of sweet-potato (Ipomoea-Batatas Lam). Plant Cell Rep. 1987, 6, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Frearson, E.M.; Power, J.; Cocking, E. The isolation, culture and regeneration of Petunia leaf protoplasts. Dev. Biol. 1973, 33, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Vasil, V.; Vasil, I.K. Regeneration of tobacco and petunia plants from protoplasts an culture of corn protoplasts. In Vitro 1974, 10, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Luo, J.H.; Cui, Z.H.; Xue, M.; Wang, L.; Zhang, X.Y.; Pawlowski, W.P.; He, Y. ATX3, ATX4, and ATX5 encode putative H3K4 methyltransferases and are critical for plant development. Plant Physiol. 2017, 174, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kao, K. Chromosomal behaviour in somatic hybrids of soybean-Nicotiana glauca. Mol. Gen. Genet. 1977, 150, 225–230. [Google Scholar] [CrossRef]
- Nitsch, C.; Nitsch, J. The induction of flowering in vitro in stem segments of Plumbago indica L. Planta 1967, 72, 355–370. [Google Scholar] [CrossRef]
- Subburaj, S.; Ha, H.J.; Jin, Y.T.; Jeon, Y.; Tu, L.; Kim, J.B.; Kang, S.Y.; Lee, G.J. Identification of gamma-radiation-responsive microRNAs and their target genes in Tradescantia (BNL clone 4430). J. Plant Biol. 2017, 60, 116–128. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormones | Concentration (mg/L) | Shoot Induction Efficiency (%) |
|---|---|---|
| 6-BAP + NAA | 2.0 + 0.1 | 3.7 d z |
| TDZ + IAA | 1.0 + 0.1 | 11.1 c |
| TDZ | 0.5 | 74.0 a |
| TDZ | 1.0 | 70.3 a |
| Zeatin | 1.0 | 77.8 a |
| Zeatin | 2.0 | 25.9 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, L.; Subburaj, S.; Lee, K.; Jeon, Y.; Yan, F.; Yao, J.; Kim, Y.-S.; Koo, O.-J.; Lee, G.-J. Optimized Regeneration of Petunia Protoplast and Its Association with Tissue Identity Regulators. Horticulturae 2023, 9, 216. https://doi.org/10.3390/horticulturae9020216
Tu L, Subburaj S, Lee K, Jeon Y, Yan F, Yao J, Kim Y-S, Koo O-J, Lee G-J. Optimized Regeneration of Petunia Protoplast and Its Association with Tissue Identity Regulators. Horticulturae. 2023; 9(2):216. https://doi.org/10.3390/horticulturae9020216
Chicago/Turabian StyleTu, Luhua, Saminathan Subburaj, Kayoun Lee, Yongsam Jeon, Fanzhuang Yan, Jian Yao, Young-Sun Kim, Ok-Jae Koo, and Geung-Joo Lee. 2023. "Optimized Regeneration of Petunia Protoplast and Its Association with Tissue Identity Regulators" Horticulturae 9, no. 2: 216. https://doi.org/10.3390/horticulturae9020216
APA StyleTu, L., Subburaj, S., Lee, K., Jeon, Y., Yan, F., Yao, J., Kim, Y.-S., Koo, O.-J., & Lee, G.-J. (2023). Optimized Regeneration of Petunia Protoplast and Its Association with Tissue Identity Regulators. Horticulturae, 9(2), 216. https://doi.org/10.3390/horticulturae9020216