
Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Light Source Design
2.2.1. Spectral Design
2.2.2. Design of Light Quality Ratio
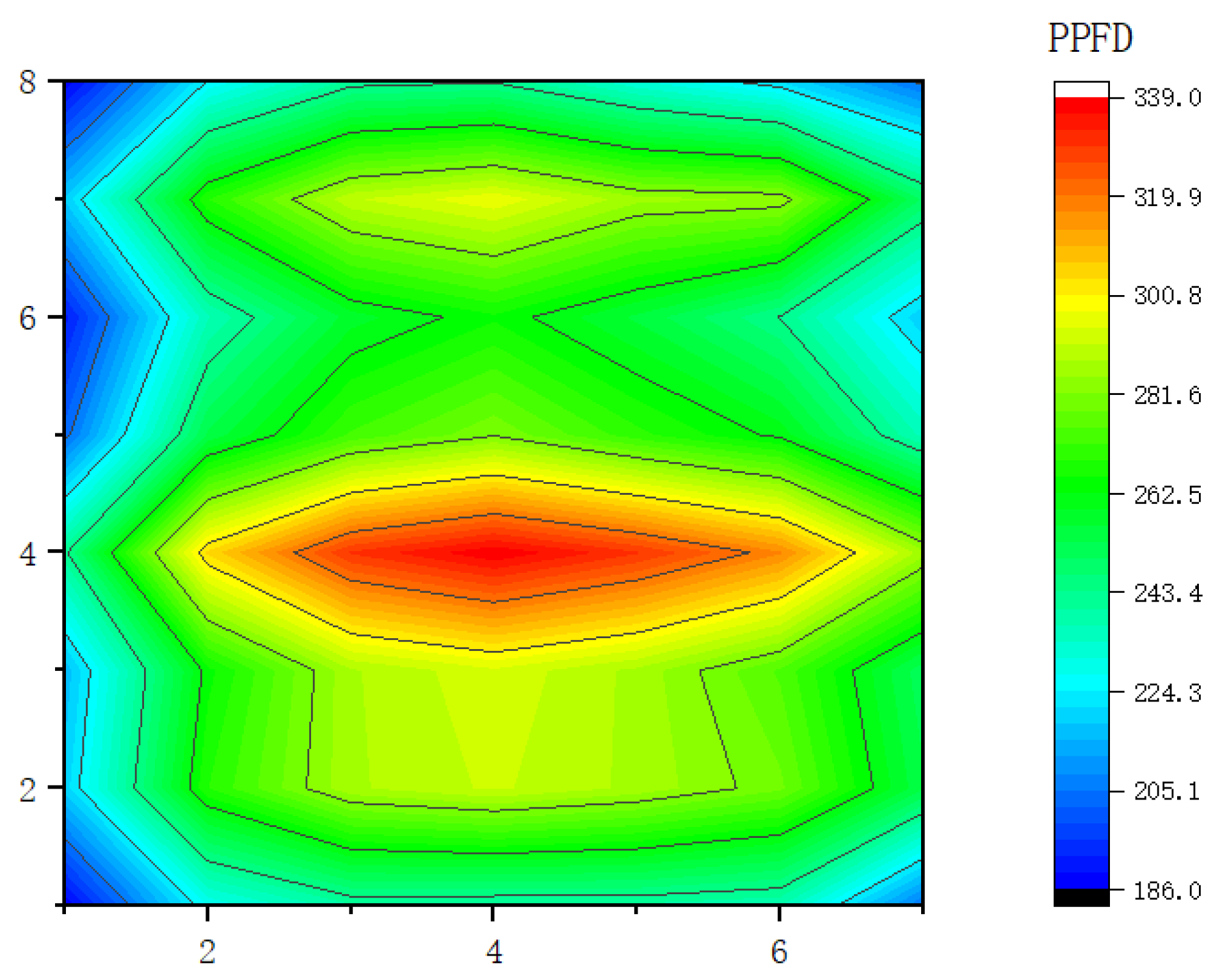
2.2.3. Optical Uniformity Design
2.3. Methods of Measurement
3. Result and Discussion
3.1. Plant Growth and Morphology
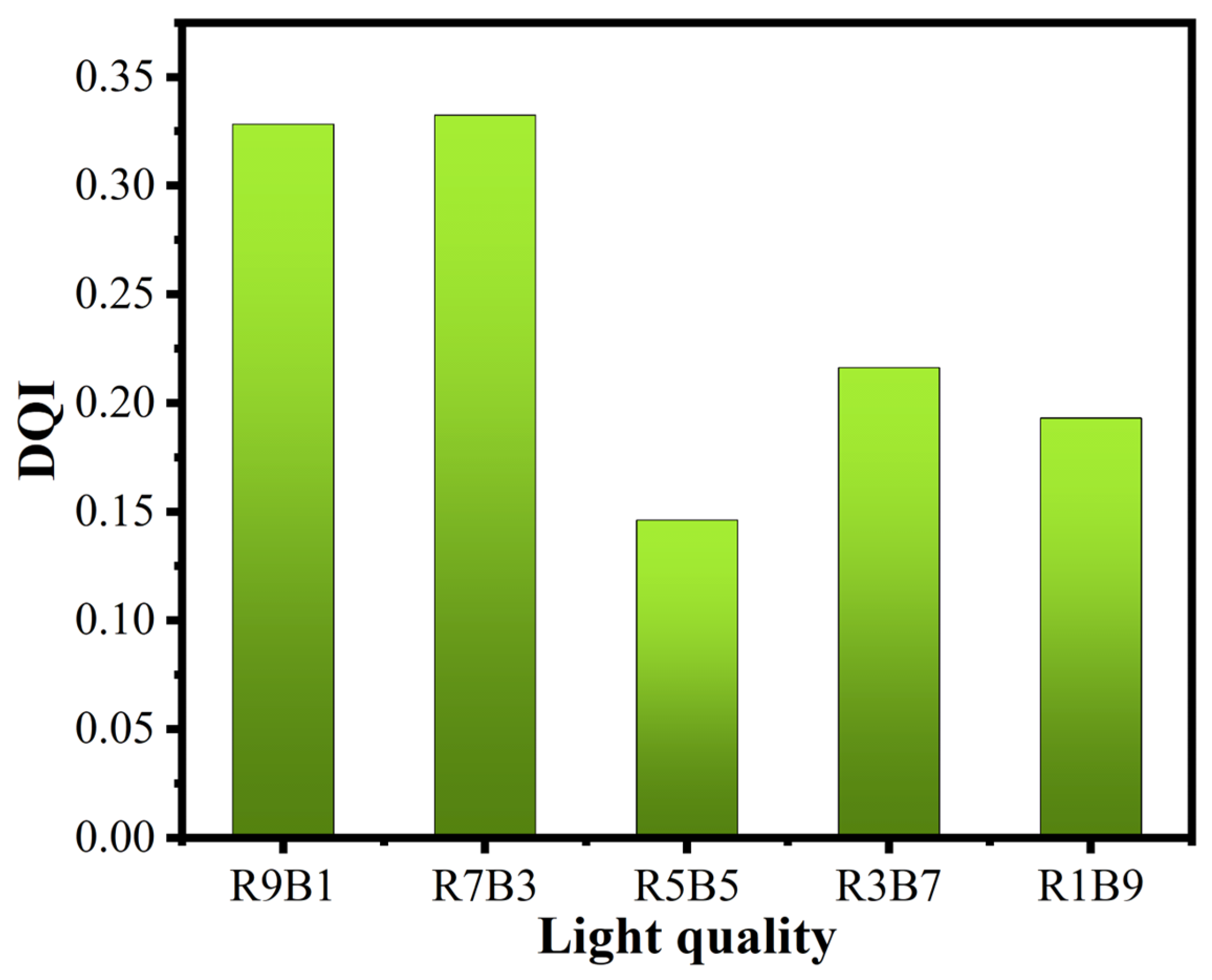
3.2. Healthy Seedling Coefficient of Seedlings
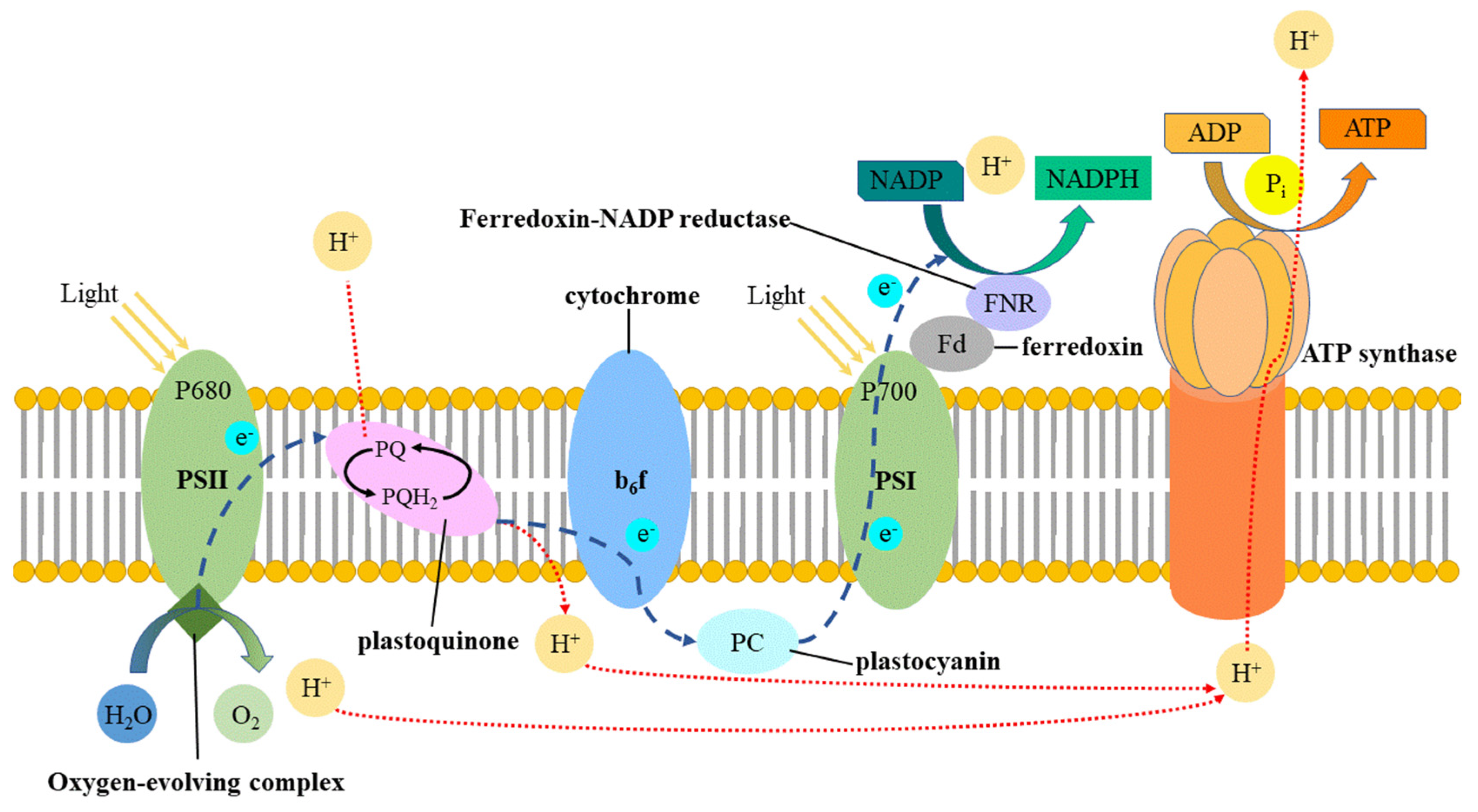
3.3. Changes in Chlorophyll Content and Photosynthetic Parameters of Seedlings
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, X.; Xu, H.; Shao, L.; Li, T.; Wang, Y.; Wang, R. Response of Photosynthetic Capacity of Tomato Leaves to Different LED Light Wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Särkkä, L.E.; Jokinen, K.; Ottosen, C.-O.; Kaukoranta, T. Effects of HPS and LED Lighting on Cucumber Leaf Photosynthesis, Light Quality Penetration and Temperature in the Canopy, Plant Morphology and Yield. AFSci 2017, 26, 102–110. [Google Scholar] [CrossRef]
- Xu, H.; Xu, Q.; Li, F.; Feng, Y.; Qin, F.; Fang, W. Applications of Xerophytophysiology in Plant Production—LED Blue Light as a Stimulus Improved the Tomato Crop. Sci. Hortic. 2012, 148, 190–196. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J. Effects of Light Quality on Growth and Phytonutrient Accumulation of Herbs under Controlled Environments. Horticulturae 2017, 3, 36. [Google Scholar] [CrossRef]
- Meiramkulova, K.; Tanybayeva, Z.; Kydyrbekova, A.; Turbekova, A.; Aytkhozhin, S.; Zhantasov, S.; Taukenov, A. The Efficiency of LED Irradiation for Cultivating High-Quality Tomato Seedlings. Sustainability 2021, 13, 9426. [Google Scholar] [CrossRef]
- Goto, E. Plant Production in a closed plant factory with artificial lighting. Acta Hortic. 2012, 956, 37–49. [Google Scholar] [CrossRef]
- Ding, B.-J.; Hofvander, P.; Wang, H.-L.; Durrett, T.P.; Stymne, S.; Löfstedt, C. A Plant Factory for Moth Pheromone Production. Nat. Commun. 2014, 5, 3353. [Google Scholar] [CrossRef] [PubMed]
- Seedapalee, T.; Inkham, C.; Ruamrungsri, S.; Jogloy, S.; Hongpakdee, P. Physiological Responses of Sun Choke’s Seedlings under Different Wavelength LED Lighting. Sci. Hortic. 2021, 282, 110029. [Google Scholar] [CrossRef]
- Liu, X.Y.; Chang, T.T.; Guo, S.R.; Xu, Z.G.; Li, J. Effect of different light quality of LED on growth and photosynthetic character in cherry tomato seedling. Acta Hortic. 2011, 907, 325–330. [Google Scholar] [CrossRef]
- McCree, K.J. The Action Spectrum, Absorptance and Quantum Yield of Photosynthesis in Crop Plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular Mechanisms Underlying Phytochrome-Controlled Morphogenesis in Plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef] [PubMed]
- Sng, B.J.R.; Mun, B.; Mohanty, B.; Kim, M.; Phua, Z.W.; Yang, H.; Lee, D.-Y.; Jang, I.-C. Combination of Red and Blue Light Induces Anthocyanin and Other Secondary Metabolite Biosynthesis Pathways in an Age-Dependent Manner in Batavia Lettuce. Plant Sci. 2021, 310, 110977. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D. Specific Functions of Red, Far Red, and Blue Light in Flowering and Stem Extension of Long-Day Plants. Jashs 2001, 126, 275–282. [Google Scholar] [CrossRef]
- Izzo, L.G.; Hay Mele, B.; Vitale, L.; Vitale, E.; Arena, C. The Role of Monochromatic Red and Blue Light in Tomato Early Photomorphogenesis and Photosynthetic Traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue Light-Emitting Diode Light Irradiation of Seedlings Improves Seedling Quality and Growth after Transplanting in Red Leaf Lettuce. Horts 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Weller, J.L.; Perrotta, G.; Schreuder, M.E.L.; Van Tuinen, A.; Koornneef, M.; Giuliano, G.; Kendrick, R.E. Genetic Dissection of Blue-Light Sensing in Tomato Using Mutants Deficient in Cryptochrome 1 and Phytochromes A, B1 and B2: Cryptochrome 1 in Tomato. Plant J. 2001, 25, 427–440. [Google Scholar] [CrossRef]
- Fantini, E.; Facella, P. Cryptochromes in the Field: How Blue Light Influences Crop Development. Physiol. Plantarum 2020, 169, 336–346. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf Morphology, Photosynthetic Performance, Chlorophyll Fluorescence, Stomatal Development of Lettuce (Lactuca Sativa, L.) Exposed to Different Ratios of Red Light to Blue Light. Front. Plant Sci. 2016, 7, 250. [Google Scholar] [CrossRef]
- Xin, X.; Chen, W.; Wang, B.; Zhu, F.; Li, Y.; Yang, H.; Li, J.; Ren, D. Arabidopsis MKK10-MPK6 Mediates Red-Light-Regulated Opening of Seedling Cotyledons through Phosphorylation of PIF3. J. Exp. Bot. 2018, 69, 423–439. [Google Scholar] [CrossRef]
- Cao, K.; Yu, J.; Ye, L.; Zhao, H.; Zou, Z. Red Light Treatments at Night during Seedling Stage in Greenhouse Promoting Tomato Vegetative Growth and Improving Yield. Trans. Chin. Soc. Agric. Eng. 2016, 32, 180–186. [Google Scholar]
- Hwang, H.; An, S.; Lee, B.; Chun, C. Improvement of Growth and Morphology of Vegetable Seedlings with Supplemental Far-Red Enriched LED Lights in a Plant Factory. Horticulturae 2020, 6, 109. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Scopel, A.L.; Sánchez, R.A.; Radosevich, S.R. Photomorphogenic processes in the agricultural environment. Photochem. Photobiol. 1992, 56, 777–788. [Google Scholar] [CrossRef]
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the Performance of Cucumber (Cucumis Sativus, L.) Seedlings by LED Illumination. Sci. Rep. 2022, 12, 852. [Google Scholar] [CrossRef] [PubMed]
- Ménard, C.; Dorais, M.; Hovi, T.; Gosselin, A. Developmental and physiological responses of tomato and cucumber to additional blue light. Acta Hortic. 2006, 711, 291–296. [Google Scholar] [CrossRef]
- Hernández, R.; Kubota, C. Growth and Morphological Response of Cucumber Seedlings to Supplemental Red and Blue Photon Flux Ratios under Varied Solar Daily Light Integrals. Sci. Hortic. 2014, 173, 92–99. [Google Scholar] [CrossRef]
- Hernández, R.; Kubota, C. Physiological Responses of Cucumber Seedlings under Different Blue and Red Photon Flux Ratios Using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Cui, J. Effects of Supplemental Lighting with LED Light Quality on Growth and Photosynthetic Characteristics of Cucumber Seedlings. China Veg. 2012, 24, 48–54. [Google Scholar]
- Song, J.; Cao, K.; Hao, Y.; Song, S.; Su, W.; Liu, H. Hypocotyl Elongation Is Regulated by Supplemental Blue and Red Light in Cucumber Seedling. Gene 2019, 707, 117–125. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue Light Dose-Responses of Leaf Photosynthesis, Morphology, and Chemical Composition of Cucumis Sativus Grown under Different Combinations of Red and Blue Light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Wang, S.; Fang, H.; Xie, J.; Wu, Y.; Tang, Z.; Liu, Z.; Lv, J.; Yu, J. Physiological Responses of Cucumber Seedlings to Different Supplemental Light Duration of Red and Blue LED. Front. Plant Sci. 2021, 12, 709313. [Google Scholar] [CrossRef]
- Wang, S.; Meng, X.; Tang, Z.; Wu, Y.; Xiao, X.; Zhang, G.; Hu, L.; Liu, Z.; Lyu, J.; Yu, J. Red and Blue LED Light Supplementation in the Morning Pre-Activates the Photosynthetic System of Tomato (Solanum Lycopersicum, L.) Leaves and Promotes Plant Growth. Agronomy 2022, 12, 897. [Google Scholar] [CrossRef]
- Novičkovas, A.; Brazaitytė, A.; Duchovskis, P.; Jankauskienė, J.; Samuolienė, G.; Virsilė, A.; Sirtautas, R.; Bliznikas, Z.; Zukauskas, A. Solid-state lamps (LEDs) for the short-wavelength supplementary lighting in greenhouses: Experimental results with cucumber. Acta Hortic. 2012, 927, 723–730. [Google Scholar] [CrossRef]
- Kim, H.-H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-Light Supplementation for Enhanced Lettuce Growth under Red- and Blue-Light-Emitting Diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Z.; Chang, T.; Guo, S. Growth and Photosynthesis of Cherry Tomato Seedling Exposed to Different Low Light of LED Light Quality. Acta Bot. Boreali-Occident. Sin. 2010, 30, 725–732. [Google Scholar]
- Li, Z.; Chen, G.; Gao, F.; Luo, J.; Li, C.; He, Z.; Zhou, X. Effects of Different Light Quality Ratio on Growth and Photosynthesis Capacity in Pakchoi under Excess Nitrate Stress. IOP Conf. Ser. Earth Environ. Sci. 2021, 621, 012105. [Google Scholar] [CrossRef]
- Dyśko, J.; Stanisław, K. Effects of LED and HPS Lighting on the Growth, Seedling Morphology and Yield of Greenhouse Tomatoes and Cucumbers. Hort. Sci. 2021, 48, 22–29. [Google Scholar] [CrossRef]
- Matysiak, B.; Kaniszewski, S.; Dyśko, J.; Kowalczyk, W.; Kowalski, A.; Grzegorzewska, M. The Impact of LED Light Spectrum on the Growth, Morphological Traits, and Nutritional Status of ‘Elizium’ Romaine Lettuce Grown in an Indoor Controlled Environment. Agriculture 2021, 11, 1133. [Google Scholar] [CrossRef]
- Matysiak, B.; Kowalski, A. The growth, photosynthetic parameters and nitrogen status of basil, coriander and oregano grown under different LED light spectra. Asphc 2021, 20, 13–22. [Google Scholar] [CrossRef]
- Barber, J.; Tran, P.D. From Natural to Artificial Photosynthesis. J. R. Soc. Interface. 2013, 10, 20120984. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Jeong, H.W.; Lee, H.R.; Kim, H.M.; Kim, H.M.; Hwang, H.S.; Hwang, S.J. Using Light Quality for Growth Control of Cucumber Seedlings in Closed-Type Plant Production System. Plants 2020, 9, 639. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Ji, F.; Xu, L.; He, D.; Key Lab. Agricultural Engineering in Structure and Environment of Ministry of Agriculture and Rural Affairs, College of Water Resources & Civil Engineering, China Agricultural University, Beijing 100083, China Effects of LED Light Quality on the Growth of Pepper Seedling in Plant Factory. Int. J. Agric. Biol. Eng. 2019, 12, 44–50. [Google Scholar]
- Baek, S.; Jeon, E.; Park, K.S.; Yeo, K.-H.; Lee, J. Monitoring of Water Transportation in Plant Stem With Microneedle Sap Flow Sensor. J. Microelectromech. Syst. 2018, 27, 440–447. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, H.; Song, S.; Su, W.; Liu, H. Morphological and Physiological Responses of Cucumber Seedlings to Supplemental LED Light under Extremely Low Irradiance. Agronomy 2020, 10, 1698. [Google Scholar] [CrossRef]
- Głowacka, B. The Effect of Blue Light on the Height and Habit of the Tomato (Lycopersicon esculentum Mill.) Transplant. Folia Hortic. 2004, 16, 3–10. [Google Scholar]
- Jinxiu, S.; Qingwu, M.; Weifen, D.; Dongxian, H. Effects of Light Quality on Growth and Development of Cucumber Seedlings in Controlled Environment. Biol. Eng. 2017, 10, 312–317. [Google Scholar]
- Silva, E.M.; Costa, G.G.S.; Andrade, A.F.; Ferreira, H.C.P.; Steiner, F. Light Spectral Quality on Production of Lettuce, Cucumber and Sweet Pepper Seedlings. Sci. Agrar. Parana. 2016, 15, 446–452. [Google Scholar] [CrossRef]
- Zhong-Xi, C.H.U.; Zhe, T.; Li-Jie, F.; Qun, Z.; Xiao-Gang, W.E.N.; Sen-Tian, S.; Xiao-Feng, Z.H.U. Effect of Different Light Quality on Photosynthetic Characteristics of Cucumber Leaves. J. Integr. Plant Biol. 1999, 41, 867–870. [Google Scholar]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light Quality Affects Photosynthesis and Leaf Anatomy of Birch Plantlets in Vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Shen, Z.; Xia, K.; Cui, J. Effects of Light Quality on the Chloroplastic Ultrastructure and Photosynthetic Characteristics of Cucumber Seedlings. Plant Growth Regul. 2014, 73, 227–235. [Google Scholar] [CrossRef]
- Omasa, K.; Shimazaki, K.-I.; Aiga, I.; Larcher, W.; Onoe, M. Image Analysis of Chlorophyll Fluorescence Transients for Diagnosing the Photosynthetic System of Attached Leaves. Plant Physiol. 1987, 84, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Chow, W.S.; Telfer, A.; Chapman, D.J.; Barber, J. State 1-State 2 Transition in Leaves and Its Association with ATP-Induced Chlorophyll Fluorescence Quenching. Biochim. Biophys. Acta-Bioenerg. 1981, 638, 60–68. [Google Scholar] [CrossRef]
- Chow, W.S.; Melis, A.; Anderson, J.M. Adjustments of Photosystem Stoichiometry in Chloroplasts Improve the Quantum Efficiency of Photosynthesis. Proc. Natl. Acad. Sci. USA 1990, 87, 7502–7506. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Photosystem-II Damage and Repair Cycle in Chloroplasts: What Modulates the Rate of Photodamage in Vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef]
- Rajagopal, S.; Murthy, S.D.S.; Mohanty, P. Effect of Ultraviolet-B Radiation on Intact Cells of the Cyanobacterium Spirulina Platensis: Characterization of the Alterations in the Thylakoid Membranes. J. Photochem. Photobiol. B Biol. 2000, 54, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Wang, X.; Gao, L.; Chen, Q.; Qu, M. Blue Light Is More Essential than Red Light for Maintaining the Activities of Photosystem II and I and Photosynthetic Electron Transport Capacity in Cucumber Leaves. J. Integr. Agric. 2016, 15, 87–100. [Google Scholar] [CrossRef]
- Kim, S.-J.; Hahn, E.-J.; Heo, J.-W.; Paek, K.-Y. Effects of LEDs on Net Photosynthetic Rate, Growth and Leaf Stomata of Chrysanthemum Plantlets in Vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Thylakoid. Available online: https://www.wikidoc.org/index.php/Thylakoid (accessed on 6 September 2012).
- Zeiger, E. The Biology of Stomatal Guard Cells. Annu. Rev. Plant. Physiol. 1983, 34, 441–447. [Google Scholar] [CrossRef]
- Frechilla, S.; Zhu, J.; Talbott, L.D.; Zeiger, E. Stomata from Npql, a Zeaxanthin-Less Arabidopsis Mutant, Lack a Specific Response to Blue Light. Plant Cell Physiol. 1999, 40, 949–954. [Google Scholar] [CrossRef]
- Zeiger, E.; Zhu, J. Role of Zeaxanthin in Blue Light Photoreception and the Modulation of Light–CO2 Interactions in Guard Cells. J. Exp. Bot. 1998, 49, 433–442. [Google Scholar] [CrossRef]
- Frechilla, S.; Talbott, L.D.; Bogomolni, R.A.; Zeiger, E. Reversal of Blue Light-Stimulated Stomatal Opening by Green Light. Plant Cell Physiol. 2000, 41, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Li, X.U.; Shiqi, L.I.U.; Liandong, Q.; Qingling, L.; Wenyan, Y.U. Effect of Light Quality on Leaf Lettuce Photosynthesis and Chlorophyll Fluorescence. 2010, 26, 96–100. Available online: https://europepmc.org/article/CBA/647442 (accessed on 7 October 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Quality | Hypocotyl Length (cm) | Plant Height (cm) | Leaf Length (cm) | Leaf Width (cm) | Stem Diameter (mm) |
|---|---|---|---|---|---|
| R9B1 | 1.90 ± 0.10b | 4.16 ± 0.38a | 7.04 ± 0.09a | 8.60 ± 0.45a | 3.77 ± 0.20a |
| R7B3 | 1.92 ± 0.33b | 3.78 ± 0.31bc | 7.02 ± 0.58a | 8.74 ± 0.43a | 3.55 ± 0.15a |
| R5B5 | 2.42 ± 0.45a | 4.02 ± 0.33ab | 6.44 ± 0.66a | 7.44 ± 0.88b | 3.65 ± 0.25a |
| R3B7 | 1.72 ± 0.04b | 3.18 ± 0.22d | 5.38 ± 0.61b | 6.38 ± 0.76c | 3.22 ± 0.19b |
| R1B9 | 1.88 ± 0.16b | 3.42 ± 0.16cd | 5.22 ± 0.32b | 6.22 ± 0.61c | 3.20 ± 0.09b |
| Light Quality | Hypocotyl Length (cm) | Plant Height (cm) | Leaf Length (cm) | Leaf Width (cm) | Stem Diameter (mm) |
| R9B1 | 2.10 ± 0.20bc | 4.34 ± 0.32ab | 10.80 ± 0.27a | 14.10 ± 0.14a | 4.41 ± 0.42a |
| R7B3 | 2.20 ± 0.27ab | 4.20 ± 0.19b | 10.32 ± 0.54a | 13.92 ± 0.66a | 4.41 ± 0.20a |
| R5B5 | 2.54 ± 0.36a | 4.64 ± 0.26a | 8.14 ± 0.65b | 9.06 ± 0.55b | 4.04 ± 0.14b |
| R3B7 | 1.76 ± 0.18c | 3.72 ± 0.26c | 7.72 ± 0.47b | 10.12 ± 0.41b | 3.95 ± 0.11b |
| R1B9 | 2.00 ± 0.38bc | 3.80 ± 0.27c | 7.60 ± 0.19b | 10.14 ± 0.75c | 3.92 ± 0.14b |
| Light Quality | Hypocotyl Length (cm) | Plant Height (cm) | Leaf Length (cm) | Leaf Width (cm) | Stem Diameter (mm) |
| R9B1 | 2.46 ± 0.17a | 6.22 ± 0.31a | 13.08 ± 0.65a | 17.12 ± 1.08a | 5.53 ± 0.41ab |
| R7B3 | 2.26 ± 0.36ab | 5.76 ± 0.40ab | 12.48 ± 0.53a | 17.12 ± 0.72a | 5.74 ± 0.30a |
| R5B5 | 2.60 ± 0.20a | 5.42 ± 0.29bc | 11.3 ± 0.91b | 13.02 ± 1.50b | 5.29 ± 0.35abc |
| R3B7 | 2.02 ± 0.33b | 4.90 ± 0.83c | 10.92 ± 0.80b | 14.00 ± 0.84b | 5.14 ± 0.33bc |
| R1B9 | 2.02 ± 0.22b | 5.40 ± 0.47bc | 9.92 ± 0.31c | 13.60 ± 0.92b | 4.93 ± 0.35c |
| Light Quality | Fresh Weight | Dry Weight | ||
|---|---|---|---|---|
| Shoot (g) | Root (g) | Shoot (g) | Root (g) | |
| R9B1 | 20.46 ± 1.23a | 20.55 ± 2.37a | 2.57 ± 0.19a | 1.65 ± 0.05a |
| R7B3 | 20.99 ± 1.28a | 19.61 ± 1.22a | 2.19 ± 0.09b | 1.63 ± 0.09a |
| R5B5 | 15.24 ± 0.94b | 13.51 ± 0.89b | 1.43 ± 0.09c | 1.06 ± 0.16b |
| R3B7 | 13.27 ± 2.19c | 8.95 ± 1.75d | 1.17 ± 0.25d | 0.63 ± 0.12c |
| R1B9 | 16.07 ± 0.82b | 11.28 ± 2.24c | 1.52 ± 0.09c | 1.01 ± 0.16b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, D.; Su, X.; Li, Y.; Shi, M.; Yang, B.; Wan, W.; Wen, X.; Yang, S.; Ding, X.; Zou, J. Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory. Horticulturae 2023, 9, 124. https://doi.org/10.3390/horticulturae9020124
Jin D, Su X, Li Y, Shi M, Yang B, Wan W, Wen X, Yang S, Ding X, Zou J. Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory. Horticulturae. 2023; 9(2):124. https://doi.org/10.3390/horticulturae9020124
Chicago/Turabian StyleJin, Dezhi, Xiaofeng Su, Yuefeng Li, Mingming Shi, Bobo Yang, Wenchang Wan, Xing Wen, Shaojun Yang, Xiaotao Ding, and Jun Zou. 2023. "Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory" Horticulturae 9, no. 2: 124. https://doi.org/10.3390/horticulturae9020124
APA StyleJin, D., Su, X., Li, Y., Shi, M., Yang, B., Wan, W., Wen, X., Yang, S., Ding, X., & Zou, J. (2023). Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory. Horticulturae, 9(2), 124. https://doi.org/10.3390/horticulturae9020124