
Somatic Embryogenesis and Flow Cytometric Assessment of Nuclear Genetic Stability for Sansevieria spp.: An Approach for In Vitro Regeneration of Ornamental Plants

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Media and Culture Conditions
2.3. Embryo Germination, Plant Development and Acclimatization
2.4. Flow Cytometry Analysis
2.5. Data Analyses
3. Results
3.1. Plant Regeneration
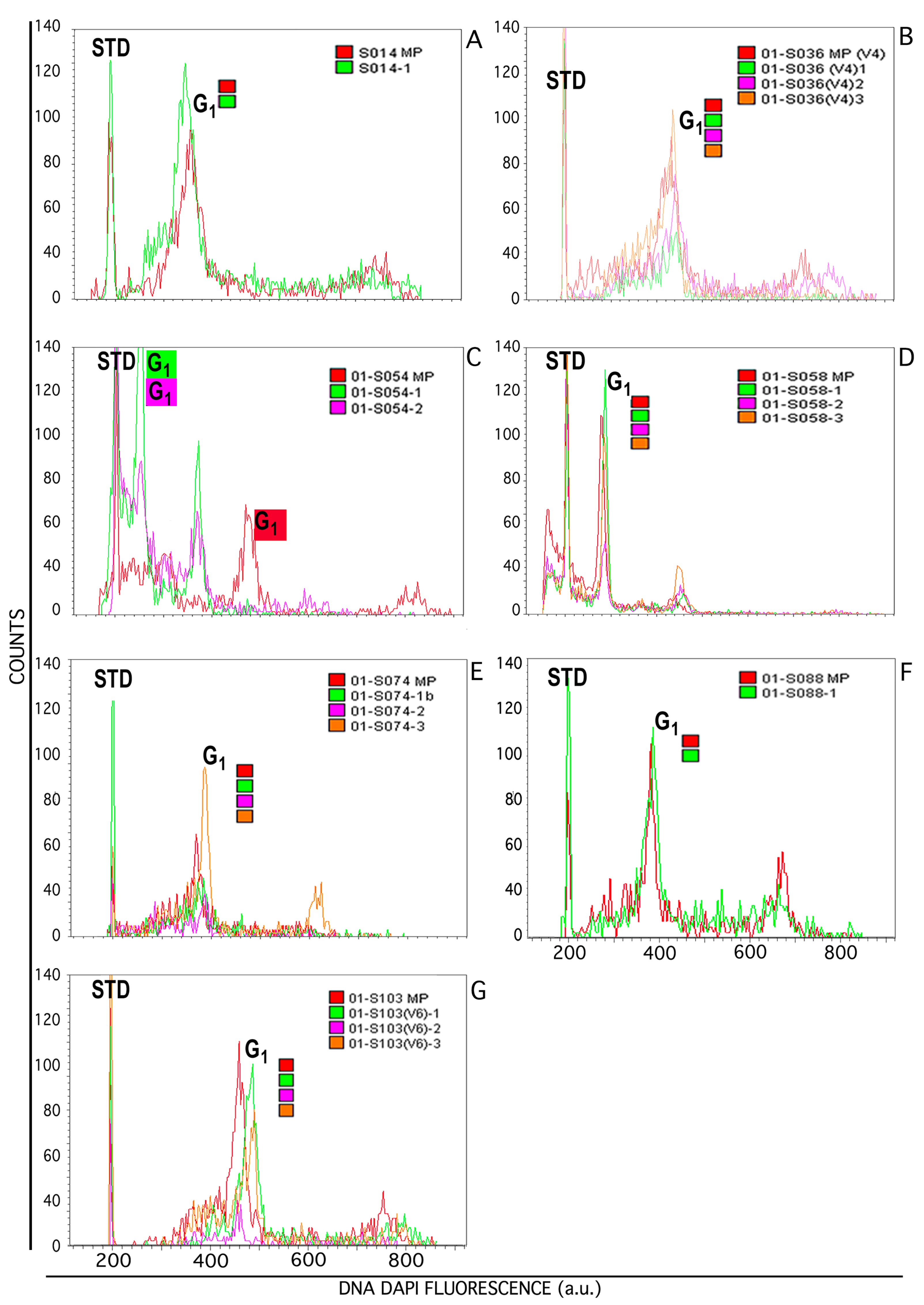
3.2. Flow Cytometric Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Webb, R.H.; Newton, L.E. The Genus Sansevieria: A Pictorial Guide to the Species; Arid Lands Press: Tucson, Arizona, 2022; p. 200. [Google Scholar]
- Brown, N.E. Sansevieria. A monograph of all the known species. Bull. Misc. Inform. Kew. 1915, 5, 185–261. [Google Scholar] [CrossRef]
- García-Hernández, E.; Loera-Quezada, M.M.; Morán-Velázquez, D.C.; López, M.G.; Chable-Vega, M.A.; Santillán-Fernández, A.; Zavaleta-Mancera, H.A.; Tang, J.Z.; Azadi, P.; Ibarra-Laclette, E.; et al. Indirect organogenesis for high frequency shoot regeneration of two cultivars of Sansevieria trifasciata Prain differing in fiber production. Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bandehali, S.; Miri, T.; Onyeaka, H.; Kumar, P. Current state of indoor air phytoremediation using potted plants and green walls. Atmosphere 2021, 12, 473. [Google Scholar] [CrossRef]
- Chatakul, P.; Janpathompong, S. Interior plants: Trends, species, and their benefits. Build Environ. 2022, 222, 109325. [Google Scholar] [CrossRef]
- Suberi, I.; Noor, N.; Darnis, D.; Mukai, Y.; Usup, G. The Potential of Ornamental Plant, Sansevieria trifasciata to Inhibit the Growth of Harmful Algal Bloom Species; Isals Publishing: Yazd, Iran, 2014; p. 42. [Google Scholar]
- Takawira-Nyenya, R.; Newton, L.E.; Wabuyele, E.; Stedje, B. Ethnobotanical uses of Sansevieria Thunb (Asparagaceae) in Coast Province of Kenya. Ethnobot Res. App. 2014, l12, 51–69. [Google Scholar]
- Yusnita, Y.; Pungkastiani, W.; Hapsoro, D. In vitro organogenesis of two Sansevieria trifasciata cultivars on different concentrations of benzyladenine (BA). AGRIVITA J. Agric. Sci. 2011, 33, 147–153. [Google Scholar] [CrossRef]
- Henley, R.W.; Chase, A.R.; Osborne, L.S. Sansevieria Production Guide. USA: University of Florida. 1991. Available online: https://mrec.ifas.uf.edu/foliage/folnotes/sansevie.htm (accessed on 10 November 2022).
- Kaur, J.; Mudgal, G. An efficient and quick protocol for in vitro multiplication of snake plant, Sansevieria trifasciata var. Laurentii [Prain]. Plant Cell Tissue Organ Cult. 2021, 147, 405–411. [Google Scholar] [CrossRef]
- Sarmast, M.K.; Salehi, M.; Salehi, H. The potential of different parts of Sansevieria trifasciata L. leaf for meristemoids production. Aust. J. Basic Appl. Sci. 2009, 3, 2506–2509. [Google Scholar]
- Agrotan, J.; Inderiati, S. Indirect organogenesis and induction of morphogenic callus for in vitro propagation of Sansevieria masoniana. Jurnal Agrotan 2015, 1, 1–8. [Google Scholar]
- Carra, A.; De Pasquale, F.; Ricci, A.; Carimi, F. Diphenylurea derivatives induce somatic embryogenesis in Citrus. Plant Cell Tissue Organ Cult. 2006, 87, 41–48. [Google Scholar] [CrossRef]
- Carra, A.; Sajeva, M.; Abbate, L.; Siragusa, M.; Pathirana, R.; Carimi, F. Factors affecting somatic embryogenesis in eight Italian grapevine cultivars and the genetic stability of embryo-derived regenerants as assessed by molecular markers. Sci. Hortic. 2016, 204, 123–127. [Google Scholar] [CrossRef]
- Carra, A.; Sajeva, M.; Abbate, L.; Siragusa, M.; Sottile, F.; Carimi, F. In vitro plant regeneration of caper (Capparis spinosa L.) from floral explants and genetic stability of regenerants. Plant Cell Tissue Organ Cult. 2012, 109, 373–381. [Google Scholar] [CrossRef]
- Fiore, M.C.; Carimi, F.; Carra, A.; Sunseri, F. Efficient plant regeneration via somatic embryogenesis in bulbing fennel using immature flower explants. In Vitro Cell. Dev. Biol. Plant. 2012, 48, 440–445. [Google Scholar] [CrossRef]
- Lou, Q.; Liu, H.; Luo, W.; Chen, K.; Liu, Y. Creating a novel petal regeneration system for function identification of colour gene of grape hyacinth. Plant Methods 2021, 17, 1–10. [Google Scholar] [CrossRef]
- de Almeida, N.V.; Rivas, E.B.; Cardoso, J.C. Somatic embryogenesis from flower tepals of Hippeastrum aiming regeneration of virus-free plants. Plant Sci. 2022, 317, 111191. [Google Scholar] [CrossRef]
- Botelho, F.B.S.; Rodrigues, C.S.; Bruzi, A.T. Ornamental plant breeding. Ornam. Hortic. 2015, 21, 9–16. [Google Scholar] [CrossRef]
- Datta, S.K. Breeding of ornamentals: Success and technological status. Nucleus 2022, 65, 107–128. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dolezel, J.; Binarova, P.; Lucretti, S. Analysis of nuclear-DNA content in plant-cells by flow-cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Otto, F. Preparation and staining of cells for high-resolution DNA analysis. In Flow Cytometry and Cell Sorting; Radbruch, A., Ed.; Springer: Berlin, Germany, 1992; pp. 101–104. [Google Scholar] [CrossRef]
- Catalano, C.; Abbate, L.; Motisi, A.; Crucitti, D.; Cangelosi, V.; Pisciotta, A.; Di Lorenzo, R.; Carimi, F.; Carra, A. Autotetraploid emergence via somatic embryogenesis in Vitis vinifera induces marked morphological changes in shoots, mature leaves, and stomata. Cells 2021, 10, 1336. [Google Scholar] [CrossRef]
- Palma, M.A.; Hall, C.R.; Collart, A.J. Repeat buying behavior for ornamental plants: A consumer profile. J. Food Distrib. Res. 2011, 42, 67–77. [Google Scholar] [CrossRef]
- Carimi, F.; Barizza, E.; Gardiman, M.; Schiavo, F.L. Somatic embryogenesis from stigmas and styles of grapevine. In Vitro Cell Dev. Biol. Plant 2005, 41, 249–252. [Google Scholar] [CrossRef]
- Sajeva, M.; Carra, A.; de Pasquale, F.; Carimi, F. Somatic embryogenesis and plant regeneration from pistil transverse thin cell layers of lemon (Citrus limon). Plant Biosyst. 2008, 142, 199–203. [Google Scholar] [CrossRef]
- Wojciechowicz, M.K. Organogenesis and somatic embryogenesis induced in petal cultures of Sedum species. Acta Biol. Cracov. Bot. 2009, 51, 83–90. [Google Scholar]
- Vidal, J.R.; Rama, J.; Taboada, L.; Martin, C.; Ibañez, M.; Segura, A.; González-Benito, M.E. Improved somatic embryogenesis of grapevine (Vitis vinifera) with focus on induction parameters and efficient plant regeneration. Plant Cell Tissue Organ Cult. 2009, 96, 85–94. [Google Scholar] [CrossRef]
- Prado, M.J.; Grueiro, M.P.; González, M.V.; Testillano, P.S.; Domínguez, C.; López, M.; Rey, M. Efficient plant regeneration through somatic embryogenesis from anthers and ovaries of six autochthonous grapevine cultivars from Galicia (Spain). Sci. Hortic 2010, 125, 342–352. [Google Scholar] [CrossRef][Green Version]
- Akin-Idowu, P.E.; Ibitoye, D.O.; Ademoyegun, O.T. Tissue culture as a plant production technique for horticultural crops. Afr. J. Biotechnol. 2009, 8, 3782–3788. [Google Scholar]
- Shumilina, D.; Kornyukhin, D.; Domblides, E.; Soldatenko, A.; Artemyeva, A. Effects of genotype and culture conditions on microspore embryogenesis and plant regeneration in Brassica rapa ssp. rapa L. Plants 2020, 9, 278. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R.; Koebner, R.M.D.; Zimny, J. Quantification of the tissue-culture induced variation in barley (Hordeum vulgare L.). BMC Plant Biol. 2007, 7, 10. [Google Scholar] [CrossRef]
- Garcia, C.; Furtado de Almeida, A.A.; Costa, M.; Britto, D.; Valle, R.; Royaert, S.; Marelli, J.P. Abnormalities in somatic embryogenesis caused by 2, 4-D: An overview. Plant Cell Tissue Organ Cult. 2019, 137, 193–212. [Google Scholar] [CrossRef]
- De-la-Peña, C.; Nic-Can, G.; Galaz-Ávalos, R.; Avilez-Montalvo, R.; Loyola-Vargas, V. The role of chromatin modifications in somatic embryogenesis in plant. Front. Plant Sci. 2015, 6, 635. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 1–18. [Google Scholar] [CrossRef]
- Bairu, M.W.; Aremu, A.O.; Van Staden, J. Somaclonal variation in plants: Causes and detection methods. Plant Growth Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Loureiro, J.; Pinto, G.; Lopes, T.; Doležel, J.; Santos, C. Assessment of ploidy stability of the somatic embryogenesis process in Quercus suber L. using flow cytometry. Planta 2005, 221, 815–822. [Google Scholar] [CrossRef]
- Sharma, S.; Bryan, G.; Winfield, M.; Millam, S. Stability of potato (Solanum tuberosum L.) plants regenerated via somatic embryos, axillary bud proliferated shoots, microtubers and true potato seeds: A comparative phenotypic, cytogenetic and molecular assessment. Planta 2007, 226, 1449–1458. [Google Scholar] [CrossRef]
- Grosso, V.; Farina, A.; Giorgi, D.; Nardi, L.; Diretto, G.; Lucretti, S. A high-throughput flow cytometry system for early screening of in vitro made polyploids in Dendrobium hybrids. Plant Cell Tissue Organ Cult. 2018, 132, 57–70. [Google Scholar] [CrossRef]
- Dolezel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef]
- Zonneveld, B.J.M.; Leitch, I.J.; Bennett, M.D. First nuclear DNA amounts in more than 300 Angiosperms. Ann. Bot. 2005, 96, 229–244. [Google Scholar] [CrossRef]
- Smarda, P.; Bure, P.; Horová, L.; Leitch, I.J.; Mucina, L.; Pacini, E.; Tichý, L.; Grulich, V.; Rotreklová, O. Ecological and evolutionary significance of genomic GC content diversity in monocots. Proc. Natl. Acad. Sci. USA 2014, 111, E4096–E4102. [Google Scholar] [CrossRef]
- Nazeer, M.A.; Khoshoo, T.N. Cytology of Some Species of Sansevieria Thunb. Cytologia 1984, 49, 325–332. [Google Scholar] [CrossRef]
- Catalano, C.; Abbate, L.; Fatta Del Bosco, S.; Motisi, A.; Carimi, F.; De Michele, R.; Mercati, F.; D’Onghia, A.M.; Carra, A. Different cell types affect the transition from juvenile to mature phase in Citrus plants regenerated through somatic embryogenesis. Plants 2022, 11, 1811. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Accession ID | G1R/G1MP | Explant Type | Regeneration Medium |
|---|---|---|---|---|
| S. concinna | S-CNR-014/MP | - | - | |
| S-CNR-014/1 | 1 | Ov | T5 | |
| S. fasciata × forskaoliana | S-CNR-074/MP | - | - | |
| S-CNR-074/1 | 1 | Ov | T4 | |
| S-CNR-074/2 | 1 | Ov | T4 | |
| S-CNR-074/3 | 1 | Ov | T4 | |
| S. forskaoliana | S-CNR-036/MP | - | - | |
| S-CNR-036/1 | 1 | Ant/Fil | T5 | |
| S-CNR-036/2 | 1 | Ant/Fil | T5 | |
| S-CNR-036/3 | 1 | Ov | T5 | |
| S. elliptica | S-CNR-103/MP | - | - | |
| S-CNR-103/1 | 1 | Ov | T16 | |
| S-CNR-103/2 | 1 | Sti/Sty | T5 | |
| S-CNR-103/3 | 1 | Sti/Sty | T5 | |
| S. parva | S-CNR-054/MP | |||
| S-CNR-054/1 | 0,6 | Sti/Sty | T16 | |
| S-CNR-054/2 | 0,6 | Ov | T16 | |
| S. pearsonii | S-CNR-058/MP | |||
| S-CNR-058/1 | 1 | Ant/Fil | T16 | |
| S-CNR-058/2 | 1 | Ant/Fil | T16 | |
| S-CNR-058/3 | 1 | Sti/Sty | T16 | |
| S. caulescens | S-CNR-088/MP | - | - | |
| S-CNR-088/1 | 1 | Sti/Sty | T16 |
| PGR Combinations | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 µM CPPU + 5 µM 2,4-D (T4) | 20 µM NOA + 4 µM TDZ (T5) | 10 µM NOA + 4.4 µM BA (T16) | ||||||||
| Ov | Sti/Sty | Ant/Fil | Ov | Sti/Sty | Ant/Fil | Ov | Sti/Sty | Ant/Fil | ||
| Genotype | Accessions ID | Embryogenic Explants (%) | ||||||||
| S. concinna | S-CNR-014 | 0 | 0 | 16 ± 7.5 d | 40 ± 6.3 c | 0 | 0 | 12 ± 4.9 d,e | 0 | 0 |
| S. fasciata × forskaoliana | S-CNR-074 | 16.8 ± 7.5 d,e | 4 ± 1.9 e | 16 ± 7.5 d,e | 100 a | 60 ± 17 b | 8 ± 4.9 d,e | 0 | 0 | 0 |
| S. forskaoliana | S-CNR-036 | 40 ± 6.3 c | 0 | 60 ± 14.1b | 40 ± 6.3 c | 0 | 44 ± 4 c | 0 | 0 | 0 |
| S. elliptica | S-CNR-103 | 0 | 0 | 0 | 0 | 60 ± 14 b | 10 ± 1.1 d | 20 ± 2.3 d | 0 | 30 ± 10 c,d |
| S. parva | S-CNR-054 | 20 ± 8.9 d | 0 | 25 ± 5 d | 0 | 24 ± 11.6 d | 32 ± 18.5 c | 0 | 6.7 ± 0.4 e | 0 |
| S. pearsonii | S-CNR-058 | 0 | 0 | 6.7 ± 1.5 e | 20 ± 11.5 d | 33.3 ± 6.7 c | 0 | 10 ± 1.8 e | 70 ± 10 b | 40 ± 3 c |
| S. caulescens | S-CNR-088 | 24 ± 4 d | 30 ± 5.7 c | 25 ± 5 d | 73.3 ± 6.7 b | 40 ± 11.6 c | 35 ± 5 c | 20 ± 6.3 d | 5 ± 1.2 e | 0 |
| Genotype | Accession ID | Embryogenic Explants (%) |
|---|---|---|
| S. concinna | S-CNR-014 | 13.3 ± 0.3 c |
| S. fasciata × forskaoliana | S-CNR-074 | 18.5 ± 1.1 b |
| S. forskaoliana | S-CNR-036 | 28.9 ± 0.5 a |
| S. elliptica | S-CNR-103 | 9.7 ± 0.7 d |
| S. parva | S-CNR-054 | 31.1 ± 0.6 a |
| S. pearsonii | S-CNR-058 | 20 ± 1.1 b |
| S. caulescens | S-CNR-088 | 27.2 ± 0.5 a |
| Media | Embryogenic Response (%) | Explant type | Embryogenic Response (%) |
|---|---|---|---|
| T4 | 8.8 + 0.1 b | Ov | 11.5 + 0.6 b |
| T5 | 18.8 + 0.2 a | Sti/Sty | 4.1 + 0.4 c |
| T16 | 19.1 + 0.5 a | Ant/Fil | 13.8 + 0.3 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catalano, C.; Carra, A.; Carimi, F.; Motisi, A.; Sajeva, M.; Butler, A.; Lucretti, S.; Giorgi, D.; Farina, A.; Abbate, L. Somatic Embryogenesis and Flow Cytometric Assessment of Nuclear Genetic Stability for Sansevieria spp.: An Approach for In Vitro Regeneration of Ornamental Plants. Horticulturae 2023, 9, 138. https://doi.org/10.3390/horticulturae9020138
Catalano C, Carra A, Carimi F, Motisi A, Sajeva M, Butler A, Lucretti S, Giorgi D, Farina A, Abbate L. Somatic Embryogenesis and Flow Cytometric Assessment of Nuclear Genetic Stability for Sansevieria spp.: An Approach for In Vitro Regeneration of Ornamental Plants. Horticulturae. 2023; 9(2):138. https://doi.org/10.3390/horticulturae9020138
Chicago/Turabian StyleCatalano, Caterina, Angela Carra, Francesco Carimi, Antonio Motisi, Maurizio Sajeva, Alan Butler, Sergio Lucretti, Debora Giorgi, Anna Farina, and Loredana Abbate. 2023. "Somatic Embryogenesis and Flow Cytometric Assessment of Nuclear Genetic Stability for Sansevieria spp.: An Approach for In Vitro Regeneration of Ornamental Plants" Horticulturae 9, no. 2: 138. https://doi.org/10.3390/horticulturae9020138
APA StyleCatalano, C., Carra, A., Carimi, F., Motisi, A., Sajeva, M., Butler, A., Lucretti, S., Giorgi, D., Farina, A., & Abbate, L. (2023). Somatic Embryogenesis and Flow Cytometric Assessment of Nuclear Genetic Stability for Sansevieria spp.: An Approach for In Vitro Regeneration of Ornamental Plants. Horticulturae, 9(2), 138. https://doi.org/10.3390/horticulturae9020138