


Antifungal Activity of Plant Waste Extracts against Phytopathogenic Fungi: Allium sativum Peels Extract as a Promising Product Targeting the Fungal Plasma Membrane and Cell Wall
 ,
,  , , , ,
, , , ,  and
and
Abstract
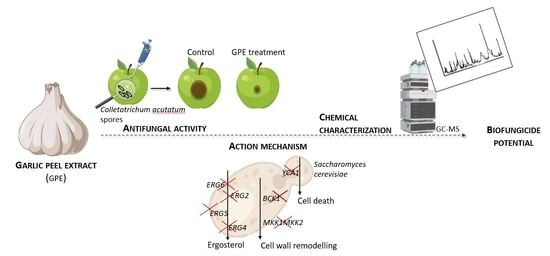
1. Introduction
2. Materials and Methods
2.1. Microorganisms, Culture Media and Growth Conditions
2.2. Plant By-Products and Extracts Preparation
2.3. Screening of Extracts Antifungal In Vitro Activity
2.4. Evaluation of Extract Antifungal Using an Ex Situ Apple Assay
2.5. Cell Viability Assays
2.6. Chemical Characterization
2.7. Statistical Analysis
3. Results
3.1. Screening of In Vitro Antifungal Activity of Plant By-Products
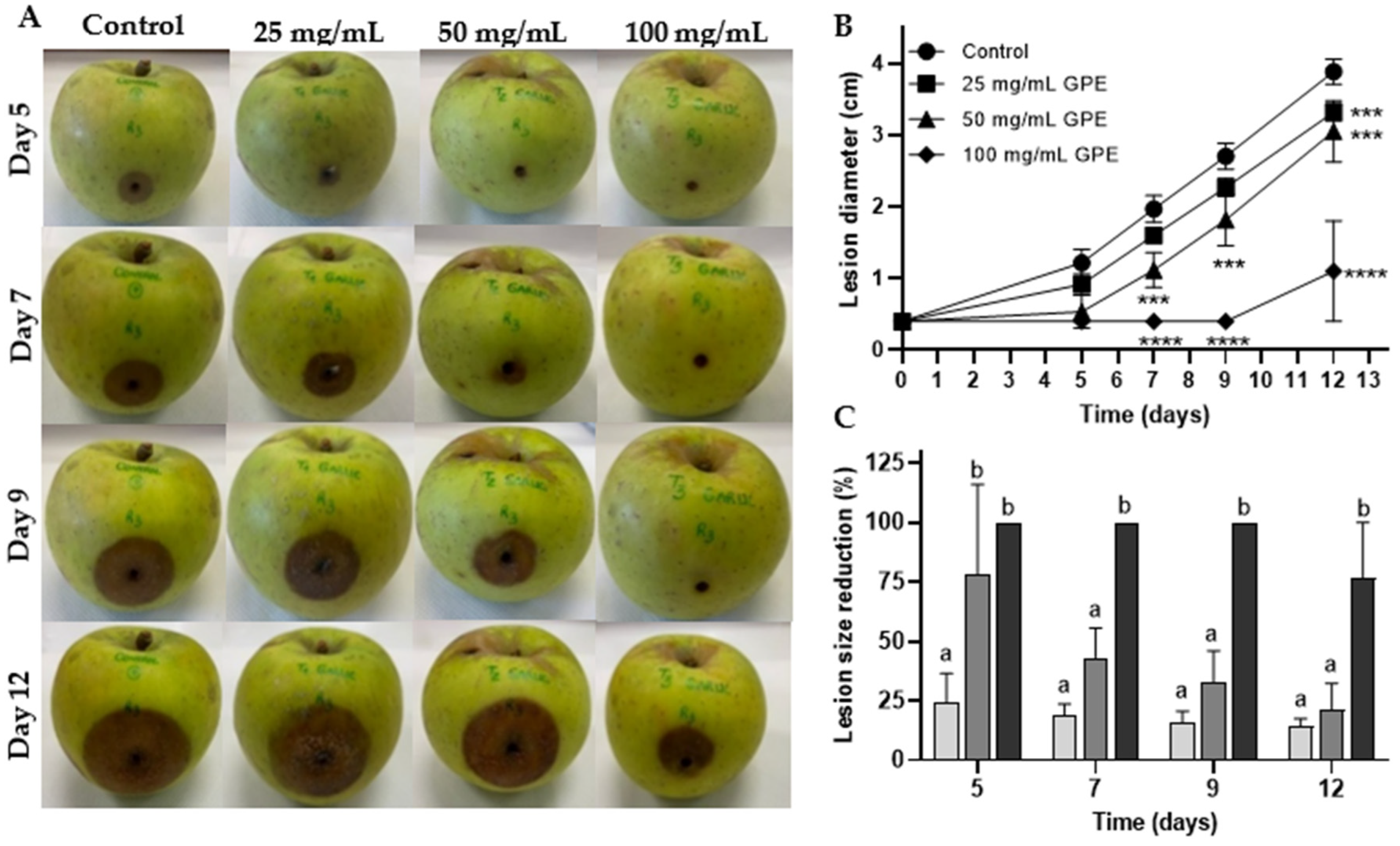
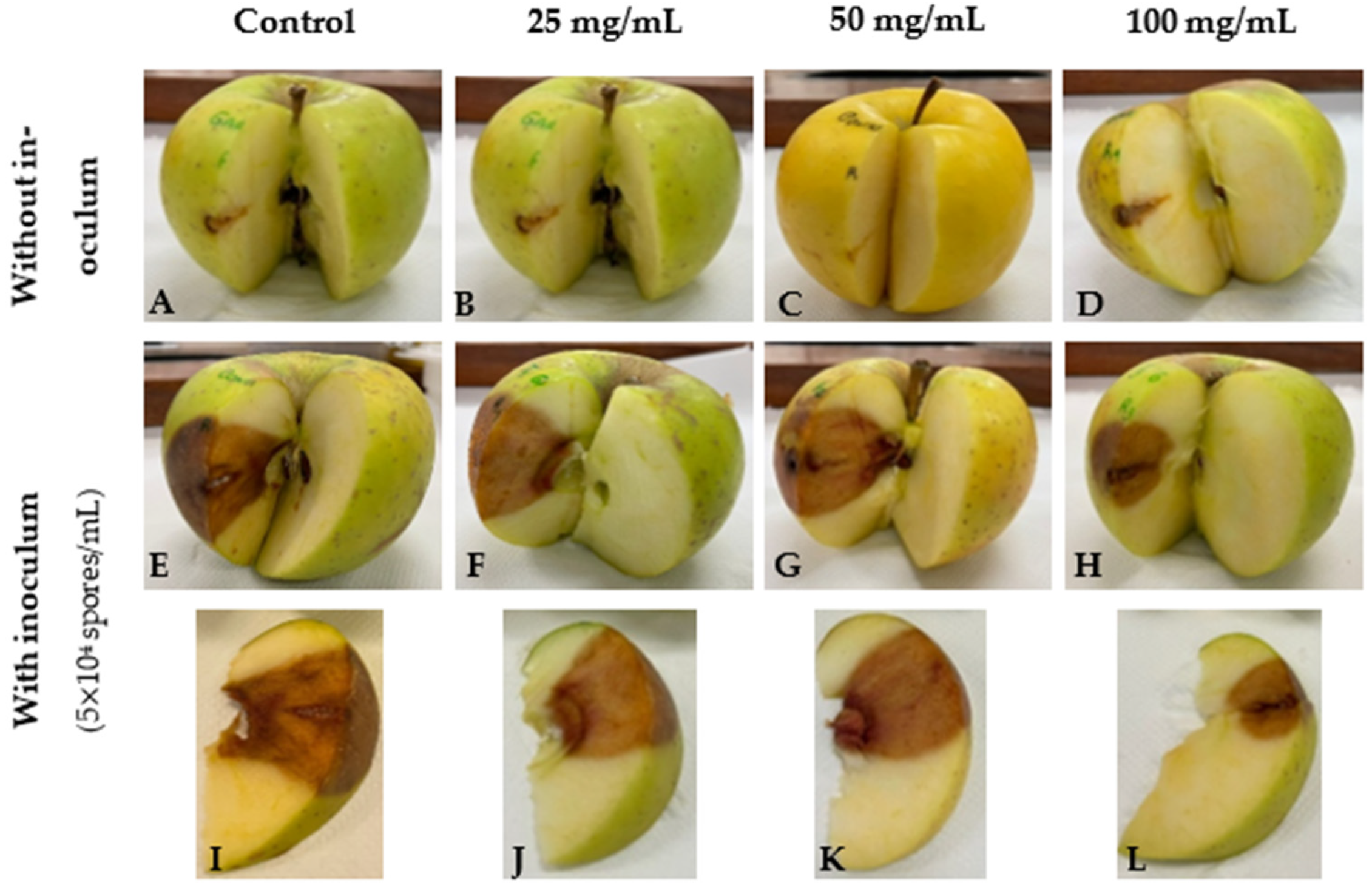
3.2. GPE as a Biocontrol Agent for C. acutatum Proliferation
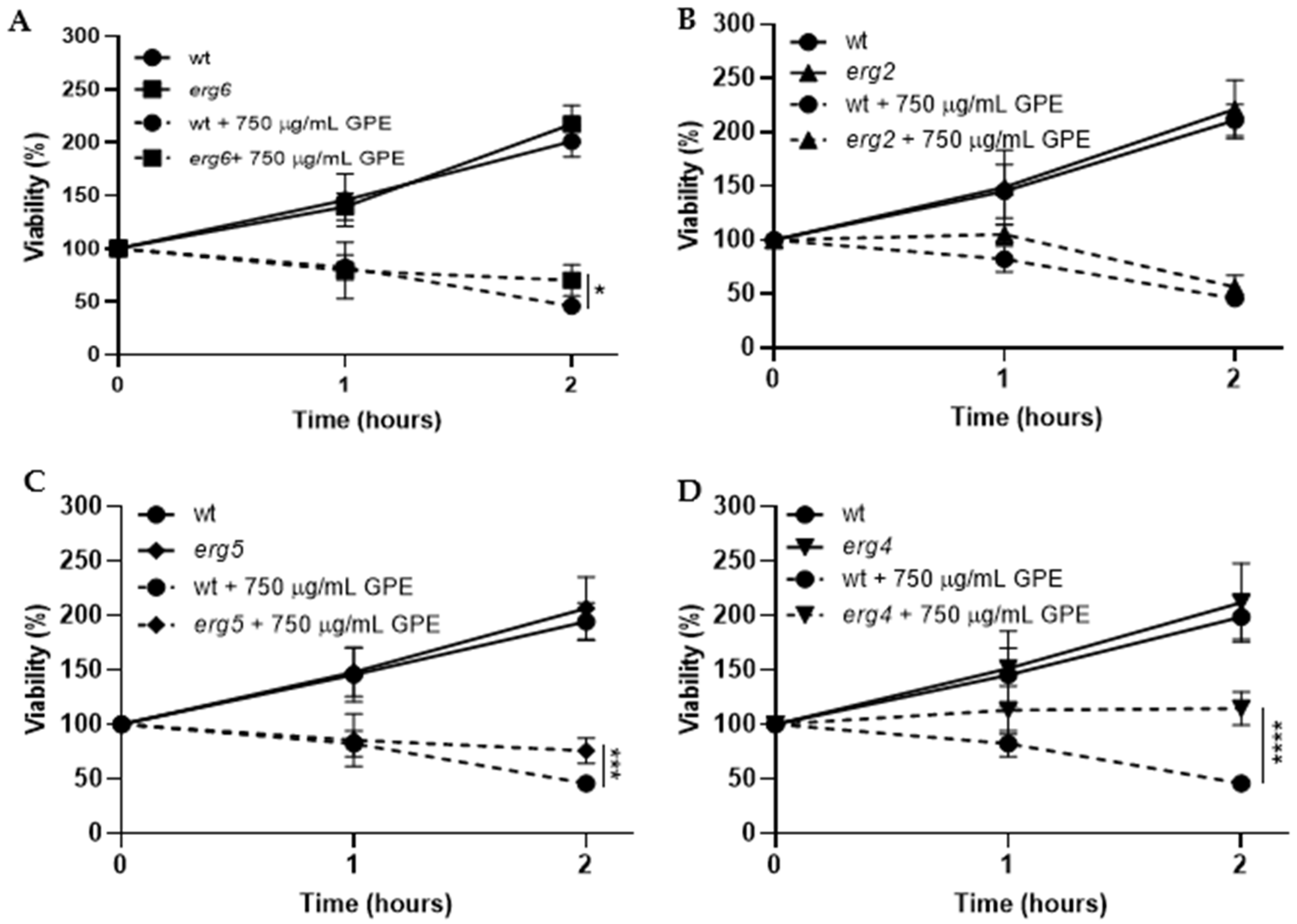
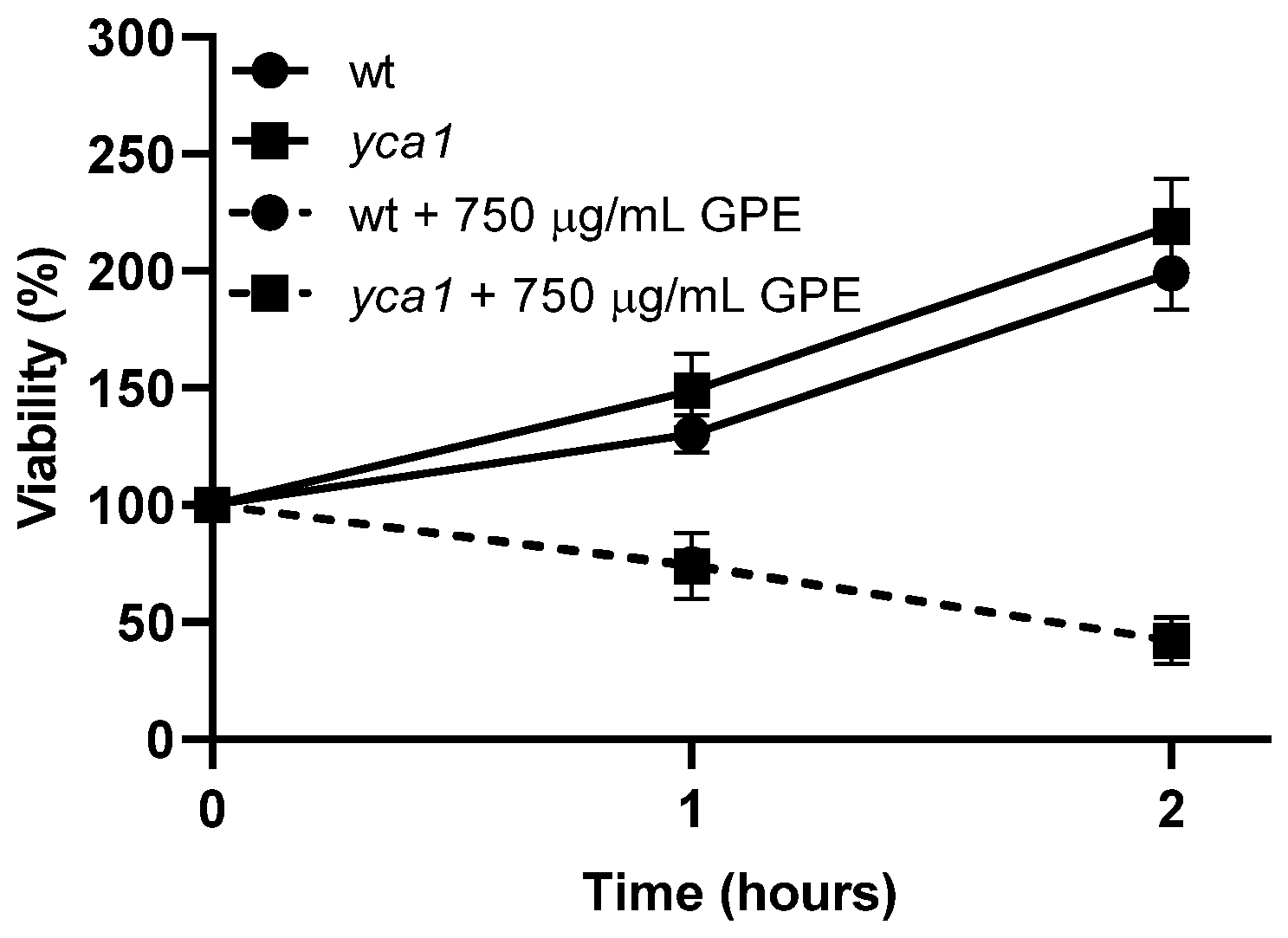
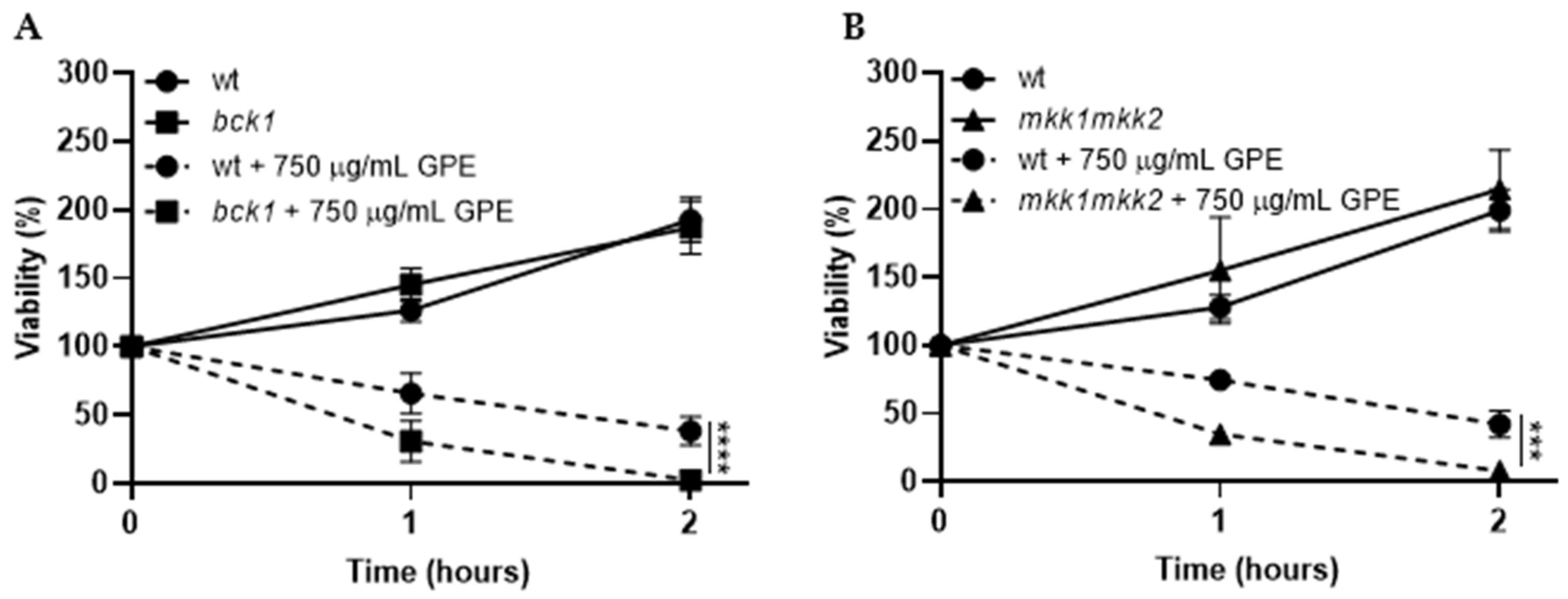
3.3. Toxicity Mechanism of GPE in S. cerevisiae Fungal Model
3.4. Chemical Characterization of GPE
3.4.1. Vibrational Characterization by ATR-FTIR
3.4.2. Phytochemicals Identified by GC–MS
4. Discussion
4.1. Phytochemical Profile
4.2. Antimicrobial Activity
4.3. Toxicity Mechanism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The Future of Food and Agriculture: Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Bajželj, B.; Richards, K.S.; Allwood, J.M.; Smith, P.; Dennis, J.S.; Curmi, E.; Gilligan, C.A. Importance of food-demand management for climate mitigation. Nat. Clim. Chang. 2014, 4, 924–929. [Google Scholar] [CrossRef]
- EUROSTAT. Waste Statistics. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Waste_statistics&oldid=527341 (accessed on 20 November 2022).
- Tomer, S.; Suyal, D.C.; Goel, R. Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Bailey, R.; Benton, T.G.; Challinor, A.; Elliott, J.; Gustafson, D.; Hiller, B.; Jones, A.; Jahn, M.; Kent, C.; Lewis, K.; et al. Extreme Weather and Resilience of the Global Food System; Final Project Report from the UK-US Taskforce on Extreme Weather and Global Food System Resilience; The Global Food Security Programme: London, UK, 2015. [Google Scholar]
- Baltazar, M.T.; Dinis-Oliveira, R.J.; de Lourdes Bastos, M.; Tsatsakis, A.M.; Duarte, J.A.; Carvalho, F. Pesticides exposure as etiological factors of Parkinson’s disease and other neurodegenerative diseases-A mechanistic approach. Toxicol. Lett. 2014, 230, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Cerejeira, M.J.; Viana, P.; Batista, S.; Pereira, T.; Silva, E.; Valério, M.J.; Silva, A.; Ferreira, M.; Silva-Fernandes, A.M. Pesticides in Portuguese surface and ground waters. Water Res. 2003, 37, 1055–1063. [Google Scholar] [CrossRef]
- Meena, R.S.; Kumar, S.; Datta, R.; Lal, R.; Vijayakumar, V.; Brtnicky, M.; Sharma, M.P.; Yadav, G.S.; Jhariya, M.K.; Jangir, C.K.; et al. Impact of agrochemicals on soil microbiota and management: A review. Land 2020, 9, 34. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Narasimhamurthy, R.K.; Joshi, M.B.; Dsouza, H.S.; Mumbrekar, K.D. Pesticides exposure-induced changes in brain metabolome: Implications in the pathogenesis of neurodegenerative disorders. Neurotox. Res. 2022, 40, 1539–1552. [Google Scholar] [CrossRef]
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. 2019, 94, 135–155. [Google Scholar] [CrossRef]
- Ranson, H.; Lissenden, N. Insecticide resistance in African anopheles mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- Van den Berg, J.; Greyvenstein, B.; du Plessis, H. Insect resistance management facing African smallholder farmers under climate change. Curr. Opin. Insect Sci. 2022, 50, 100894. [Google Scholar] [CrossRef]
- Gwinn, K.D. Bioactive Natural Products in Plant Disease Control, 1st ed.; Elsevier, B.V., Ed.; Elsevier: Gurgaon, India, 2018; Volume 56. [Google Scholar] [CrossRef]
- Czaja, K.; Góralczyk, K.; Struciński, P.; Hernik, A.; Korcz, W.; Minorczyk, M.; Łyczewska, M.; Ludwicki, J.K. Biopesticides—Towards increased consumer safety in the European Union. Pest Manag. Sci. 2015, 71, 3–6. [Google Scholar] [CrossRef]
- Hans, A.L.; Sanxena, S. Plant bioprospecting for biopesticides and bioinsecticides. In Bioprospecting of Plant Biodiversity for Industrial Molecules; Upadhyay, S.K., Singh, S.P., Eds.; John Wiley & Sons Ltd: Oxford, UK, 2021; pp. 335–344. [Google Scholar] [CrossRef]
- Sánchez-Hernández, E.; Buzón-Durán, L.; Lorenzo-Vidal, B.; Martín-Gil, J.; Martín-Ramos, P. Physicochemical characterization and antimicrobial activity against Erwinia amylovora, Erwinia vitivora, and Diplodia seriata of a light purple Hibiscus syriacus L. cultivar. Plants 2021, 10, 1876. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, J.; Liu, Y.Q. Inhibitory activity of dihydrosanguinarine and dihydrochelerythrine against phytopathogenic fungi. Nat. Prod. Res. 2011, 25, 1082–1089. [Google Scholar] [CrossRef]
- Ramírez-Pelayo, C.; Martínez-Quiñones, J.; Gil, J.; Durango, D. Coumarins from the peel of citrus grown in Colombia: Composition, elicitation and antifungal activity. Heliyon 2019, 5, e01937. [Google Scholar] [CrossRef] [PubMed]
- Kosina, P.; Gregorova, J.; Gruz, J.; Vacek, J.; Kolar, M.; Vogel, M.; Roos, W.; Naumann, K.; Simanek, V.; Ulrichova, J. Phytochemical and antimicrobial characterization of Macleaya cordata herb. Fitoterapia 2010, 81, 1006–1012. [Google Scholar] [CrossRef]
- Ntalli, N.; Parlapani, A.B.; Tzani, K.; Samara, M.; Boutsis, G.; Dimou, M.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Thymus citriodorus (Schreb) Botanical Products as Ecofriendly Nematicides with Bio-Fertilizing Properties. Plants 2020, 9, 202. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Pereira, L.; Cunha, A.; Almeida-Aguiar, C. Portuguese propolis from Caramulo as a biocontrol agent of the apple blue mold. Food Control 2022, 139, 109071. [Google Scholar] [CrossRef]
- Loebler, M.; Sánchez, C.; Maurício, E.M.; Diogo, E.; Santos, M.; Vasilenko, P.; Cruz, A.S.; Mendes, B.; Gonçalves, M.; Duarte, M.P. Potential application of propolis extracts to control the growth of Stemphylium vesicarium in ’Rocha’pear. Appl. Sci. 2020, 10, 1990. [Google Scholar] [CrossRef]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on fruit crops: A “complex” challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef]
- Simons, V.; Morrissey, J.P.; Latijnhouwers, M.; Csukai, M.; Cleaver, A.; Yarrow, C.; Osbourn, A. Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2006, 50, 2732–2740. [Google Scholar] [CrossRef] [PubMed]
- Troppens, D.M.; Dmitriev, R.I.; Papkovsky, D.B.; O’Gara, F.; Morrissey, J.P. Genome-wide investigation of cellular targets and mode of action of the antifungal bacterial metabolite 2,4-diacetylphloroglucinol in Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 322–334. [Google Scholar] [CrossRef]
- Kodedová, M.; Sychrová, H. Changes in the sterol composition of the plasma membrane affect membrane potential, salt tolerance and the activity of multidrug resistance pumps in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0139306. [Google Scholar] [CrossRef]
- Ludovico, P.; Sousa, M.J.; Silva, M.T.; Leão, C.; Côrte-Real, M. Saccharomyces cerevisiae commits to a programmed cell death process in response to acetic acid. Microbiology 2001, 147, 2409–2415. [Google Scholar] [CrossRef]
- de Castro, P.A.; Savoldi, M.; Bonatto, D.; Barros, M.H.; Goldman, M.H.S.; Berretta, A.A.; Goldman, G.H. Molecular characterization of propolis-induced cell death in Saccharomyces cerevisiae. Eukaryot. Cell 2011, 10, 398–411. [Google Scholar] [CrossRef]
- Guaragnella, N.; Pereira, C.; Sousa, M.J.; Antonacci, L.; Passarella, S.; Côrte-Real, M.; Marra, E.; Giannattasio, S. YCA1 participates in the acetic acid induced yeast programmed cell death also in a manner unrelated to its caspase-like activity. FEBS Lett. 2006, 580, 6880–6884. [Google Scholar] [CrossRef]
- Wong, A.H.H.; Yan, C.; Shi, Y. Crystal structure of the yeast metacaspase Yca1. J. Biol. Chem. 2012, 287, 29251–29259. [Google Scholar] [CrossRef]
- Madeo, F.; Herker, E.; Maldener, C.; Wissing, S.; Lächelt, S.; Herlan, M.; Fehr, M.; Lauber, K.; Sigrist, S.J.; Wesselborg, S.; et al. A caspase-related protease regulates apoptosis in yeast. Mol. Cell 2002, 9, 911–917. [Google Scholar] [CrossRef]
- Severin, F.F.; Meer, M.V.; Smirnova, E.A.; Knorre, D.A.; Skulachev, V.P. Natural causes of programmed death of yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 1350–1353. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Lorberg, A.; Schmitz, H.P.; Jacoby, J.J. The protein kinase C-mediated MAP kinase pathway involved in the maintenance of cellular integrity in Saccharomyces cerevisiae. Mol. Microbiol. 1999, 32, 671–680. [Google Scholar] [CrossRef]
- Süfer, Ö.; Bozok, F. Determination of volatile components and antioxidnt activity of essential oil obtained from Kastamonu garlic by microwave-assisted clevenger system. Gida J. Food 2019, 44, 22–30. [Google Scholar] [CrossRef]
- Harris, J.C.; Cottrell, S.L.; Plummer, S.; Lloyd, D. Antimicrobial properties of Allium sativum (garlic). Appl. Microbiol. Biotechnol. 2001, 57, 282–286. [Google Scholar] [CrossRef]
- Ariga, T.; Seki, T. Antithrombotic and anticancer effects of garlic-derived sulfur compounds: A review. Biofactors 2006, 26, 93–103. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Portz, D.; Stitz, M.; Anwar, A.; Schneider, T.; Jacob, C.; Schlaich, N.L.; Slusarenko, A.J. Allicin disrupts the cell’s electrochemical potential and induces apoptosis in yeast. Free Radic. Biol. Med. 2010, 49, 1916–1924. [Google Scholar] [CrossRef]
- Lee, H.H.; Jeong, J.-W.; Hong, S.H.; Park, C.; Kim, B.W.; Choi, Y.H. Diallyl trisulfide suppresses the production of lipopolysaccharide-induced inflammatory mediators in BV2 microglia by decreasing the NF-κB pathway activity associated with toll-like receptor 4 and CXCL12/CXCR4 Pathway Blockade. J. Cancer Prev. 2018, 23, 134–140. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, S.X.; Ho, S.H. Bioactivities of methyl allyl disulfide and diallyl trisulfide from essential oil of garlic to two species of stored-product pests, Sitophilus zeamais (Coleoptera: Curculionidae) and Tribolium castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2000, 93, 537–543. [Google Scholar] [CrossRef]
- Curtis, H.; Noll, U.; Störmann, J.; Slusarenko, A.J. Broad-spectrum activity of the volatile phytoanticipin allicin in extracts of garlic (Allium sativum L.) against plant pathogenic bacteria, fungi and Oomycetes. Physiol. Mol. Plant Pathol. 2004, 65, 79–89. [Google Scholar] [CrossRef]
- Ji, X.; Li, J.; Meng, Z.; Dong, S.; Zhang, S.; Qiao, K. Inhibitory effect of allicin against Meloidogyne incognita and Botrytis cinerea in tomato. Sci. Hortic. (Amst.) 2019, 253, 203–208. [Google Scholar] [CrossRef]
- Sharma, R.L.; Ahir, R.R.; Yadav, S.L.; Sharma, P.; Ghasolia, R.P. Effect of nutrients and plant extracts on Alternaria blight of tomato caused by Alternaria Alternata. J. Plant Dis. Prot. 2021, 128, 951–960. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Dawood, D.H.; Elsebai, M.F.; Mira, A.; Taher, M.A. Control of dry rot and resistance induction in potato tubers against Fusarium sambucinum using red onion peel extract. Postharvest Biol. Technol. 2023, 195, 112119. [Google Scholar] [CrossRef]
- Behiry, S.I.; Okla, M.K.; Alamri, S.A.; EL-Hefny, M.; Salem, M.Z.M.; Alaraidh, I.A.; Ali, H.M.; Al-Ghtani, S.M.; Monroy, J.C.; Salem, A.Z.M. Antifungal and antibacterial activities of musa. Processes 2019, 7, 215. [Google Scholar] [CrossRef]
- T-Thienprasert, N.P.; T-Thienprasert, J.; Ruangtong, J.; Jaithon, T.; Srifah Huehne, P.; Piasai, O. Large scale synthesis of green synthesized zinc oxide nanoparticles from banana peel extracts and their inhibitory effects against Colletotrichum sp., isolate KUFC 021, causal agent of anthracnose on dendrobium orchid. J. Nanomater. 2021, 2021, 5625199. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Salem, M.Z.M.; Kordy, A.M.; Salem, A.Z.M.; Behiry, S.I. Antiviral, antifungal, and insecticidal activities of Eucalyptus bark extract: HPLC analysis of polyphenolic compounds. Microb. Pathog. 2020, 147, 104383. [Google Scholar] [CrossRef]
- Alfredsen, G.; Solheim, H.; Slimestad, R. Antifungal effect of bark extracts from some European tree species. Eur. J. For. Res. 2008, 127, 387–393. [Google Scholar] [CrossRef]
- Jung, K.; Moon, S.; Lee, U. Furfural from pine needle extract inhibits the growth of a plant pathogenic fungus, Alternaria mali. Korean Soc. Mycol. 2007, 35, 39–43. [Google Scholar] [CrossRef]
- Hernández, A.; Ruiz-Moyano, S.; Galván, A.I.; Merchán, A.V.; Pérez Nevado, F.; Aranda, E.; Serradilla, M.J.; Córdoba, M.d.G.; Martín, A. Anti-fungal activity of phenolic sweet orange peel extract for controlling fungi responsible for post-harvest fruit decay. Fungal Biol. 2021, 125, 143–152. [Google Scholar] [CrossRef]
- Liu, Y.; Benohoud, M.; Galani Yamdeu, J.H.; Gong, Y.Y.; Orfila, C. Green extraction of polyphenols from citrus peel by-products and their antifungal activity against Aspergillus flavus. Food Chem. 2021, 12, 100144. [Google Scholar] [CrossRef]
- Khalifa, I.; Barakat, H.; El-Mansy, H.A.; Soliman, S.A. Improving the shelf-life stability of apple and strawberry fruits applying chitosan-incorporated olive oil processing residues coating. Food Packag. Shelf Life 2016, 9, 10–19. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Badolati, G.; Chiappetta, A.; Picci, N.; Muzzalupo, R. In vitro antifungal activity of olive (Olea europaea) leaf extracts loaded in chitosan nanoparticles. Front. Bioeng. Biotechnol. 2020, 8, 151. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Amin, B.H.; Baka, Z.A. Efficiency of pomegranate (Punica granatum L.) peels extract as a high potential natural tool towards Fusarium dry rot on potato tubers. Postharvest Biol. Technol. 2016, 111, 256–263. [Google Scholar] [CrossRef]
- Mari, M.; Guidarelli, M.; Martini, C.; Spadoni, A. First report of Colletotrichum acutatum causing bitter rot on apple in Italy. Plant Dis. 2012, 96, 144. [Google Scholar] [CrossRef]
- Sutton, B.C. Compendium of apple and pear diseases. In The American Phytopathological Society; Aldwinckle, H.S., Jones, A.L., Eds.; APS Press: St. Paul, MN, USA, 1990; pp. 15–16. [Google Scholar]
- Chen, Y.; Fu, D.; Wang, W.; Gleason, M.L.; Zhang, R.; Liang, X.; Sun, G. Diversity of Colletotrichum species causing apple bitter rot and glomerella leaf spot in China. J. Fungi 2022, 8, 740. [Google Scholar] [CrossRef] [PubMed]
- de Alvindia, D.G.; Mangoba, M.A.A. Bioactivities of Allium longicuspis regel against anthracnose of mango caused by Colletotrichum gloeosporioides (Penz.). Sci. Rep. 2020, 10, 11367. [Google Scholar] [CrossRef] [PubMed]
- Van den Bosch, F.; Paveley, N.; Shaw, M.; Hobbelen, P.; Oliver, R. The dose rate debate: Does the risk of fungicide resistance increase or decrease with dose? Plant Pathol. 2011, 60, 597–606. [Google Scholar] [CrossRef]
- Hu, M.J.; Grabke, A.; Schnabel, G. Investigation of the Colletotrichum gloeosporioides species complex causing peach anthracnose in South Carolina. Plant Dis. 2015, 99, 797–805. [Google Scholar] [CrossRef]
- Daniel, C.K.; Lennox, C.L.; Vries, F.A. In vivo application of garlic extracts in combination with clove oil to prevent postharvest decay caused by Botrytis cinerea, Penicillium expansum and Neofabraea alba on apples. Postharvest Biol. Technol. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Hosseini, S.; Amini, J.; Saba, M.K.; Karimi, K.; Pertot, I. Preharvest and postharvest application of garlic and rosemary essential oils for controlling anthracnose and quality assessment of strawberry fruit during cold storage. Front. Microbiol. 2020, 11, 1855. [Google Scholar] [CrossRef]
- Khaliq, G.; Abbas, H.T.; Ali, I.; Waseem, M. Aloe vera gel enriched with garlic essential oil effectively controls anthracnose disease and maintains postharvest quality of banana fruit during storage. Hortic. Environ. Biotechnol. 2019, 60, 659–669. [Google Scholar] [CrossRef]
- Fromtling, R.A. Overview of medically important antifungal azole derivatives. Clin. Microbiol. Rev. 1988, 1, 187–217. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Shalini, K.; Kumar, N.; Drabu, S.; Sharma, P.K. Advances in synthetic approach to and antifungal activity of triazoles. Beilstein J. Org. Chem. 2011, 7, 668–677. [Google Scholar] [CrossRef]
- Ogita, A.; Yutani, M.; Fujita, K.I.; Tanaka, T. Dependence of vacuole disruption and independence of potassium ion efflux in fungicidal activity induced by combination of amphotericin B and allicin against Saccharomyces cerevisiae. J. Antibiot. 2010, 63, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Jahanshiri, Z.; Shams-Ghahfarokhi, M.; Asghari-Paskiabi, F.; Saghiri, R.; Razzaghi-Abyaneh, M. α-Bisabolol inhibits Aspergillus fumigatus Af239 growth via affecting microsomal∆24-sterol methyltransferase as a crucial enzyme in ergosterol biosynthesis pathway. World J. Microbiol. Biotechnol. 2017, 33, 55. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.; Donaghey, F.; Helming, K.; McCarthy, M.; Rogers, S.; Austriaco, N. Deletion of AIF1 but not of YCA1/MCA1 protects Saccharomyces cerevisiae and Candida albicans cells from caspofungin-induced programmed cell death. Microb. Cell 2014, 1, 58–63. [Google Scholar] [CrossRef]
- Reinoso-Martín, C.; Schüller, C.; Schuetzer-Muehlbauer, M.; Kuchler, K. The yeast protein kinase C cell integrity pathway mediates tolerance to the antifungal drug caspofungin through activation of Slt2p mitogen-activated protein kinase signaling. Eukaryot. Cell 2003, 2, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Chan, K.; Cheng, L. Octyl gallate as an intervention catalyst to augment antifungal efficacy of caspofungin. J 2018, 1, 19–28. [Google Scholar] [CrossRef]
- Valiante, V.; Jain, R.; Heinekamp, T.; Brakhage, A.A. The MpkA MAP kinase module regulates cell wall integrity signaling and pyomelanin formation in Aspergillus fumigatus. Fungal Genet. Biol. 2009, 46, 909–918. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytopathogenic Fungi | ||||
|---|---|---|---|---|
| Plant by-Products Extracts (750 µg/mL) | D. corticola | B. cinerea | C. nymphaeae | P. cinnamomi |
| Banana peels | + | ++ | - | - |
| Eucalyptus bark | + | + | + | - |
| Garlic peels | +++ | ++ | +++ | +++ |
| Lemon peels | ++ | ++ | - | ++ |
| Olive leaves | + | +++ | - | - |
| Onion peels | ++ | ++ | +++ | +++ |
| Orange peels | ++ | ++ | - | ++ |
| Pine bark | ++ | + | + | ++ |
| Pine needles | ++ | +++ | + | - |
| Pomegranate peels | - | ++ | - | - |
| Potato peels | ++ | ++ | - | NT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, A.; Sánchez-Hernández, E.; Noversa, J.; Cunha, A.; Cortez, I.; Marques, G.; Martín-Ramos, P.; Oliveira, R. Antifungal Activity of Plant Waste Extracts against Phytopathogenic Fungi: Allium sativum Peels Extract as a Promising Product Targeting the Fungal Plasma Membrane and Cell Wall. Horticulturae 2023, 9, 136. https://doi.org/10.3390/horticulturae9020136
Teixeira A, Sánchez-Hernández E, Noversa J, Cunha A, Cortez I, Marques G, Martín-Ramos P, Oliveira R. Antifungal Activity of Plant Waste Extracts against Phytopathogenic Fungi: Allium sativum Peels Extract as a Promising Product Targeting the Fungal Plasma Membrane and Cell Wall. Horticulturae. 2023; 9(2):136. https://doi.org/10.3390/horticulturae9020136
Chicago/Turabian StyleTeixeira, Ana, Eva Sánchez-Hernández, João Noversa, Ana Cunha, Isabel Cortez, Guilhermina Marques, Pablo Martín-Ramos, and Rui Oliveira. 2023. "Antifungal Activity of Plant Waste Extracts against Phytopathogenic Fungi: Allium sativum Peels Extract as a Promising Product Targeting the Fungal Plasma Membrane and Cell Wall" Horticulturae 9, no. 2: 136. https://doi.org/10.3390/horticulturae9020136
APA StyleTeixeira, A., Sánchez-Hernández, E., Noversa, J., Cunha, A., Cortez, I., Marques, G., Martín-Ramos, P., & Oliveira, R. (2023). Antifungal Activity of Plant Waste Extracts against Phytopathogenic Fungi: Allium sativum Peels Extract as a Promising Product Targeting the Fungal Plasma Membrane and Cell Wall. Horticulturae, 9(2), 136. https://doi.org/10.3390/horticulturae9020136