
Morpho-Physiological and Transcriptional Regulation of Root System under Saline Conditions in Nymphaea Plants

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Salt Treatment
2.2. Plant Growth and Root Measurement
2.3. Transcriptome Analysis of Roots
2.4. ICP-MS Analysis of Na+ and K+ Contents
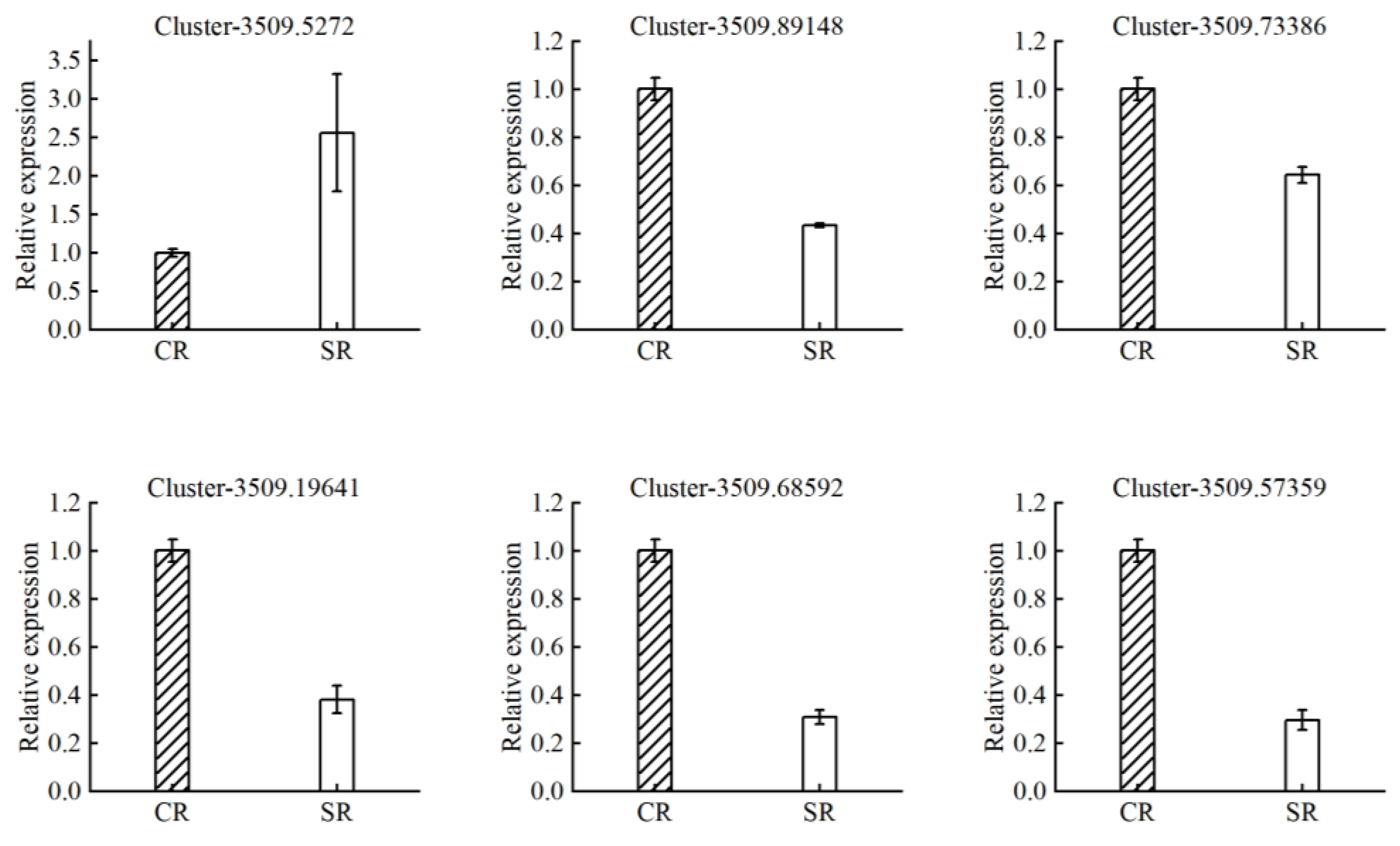
2.5. Quantitative Real-Time PCR
3. Results
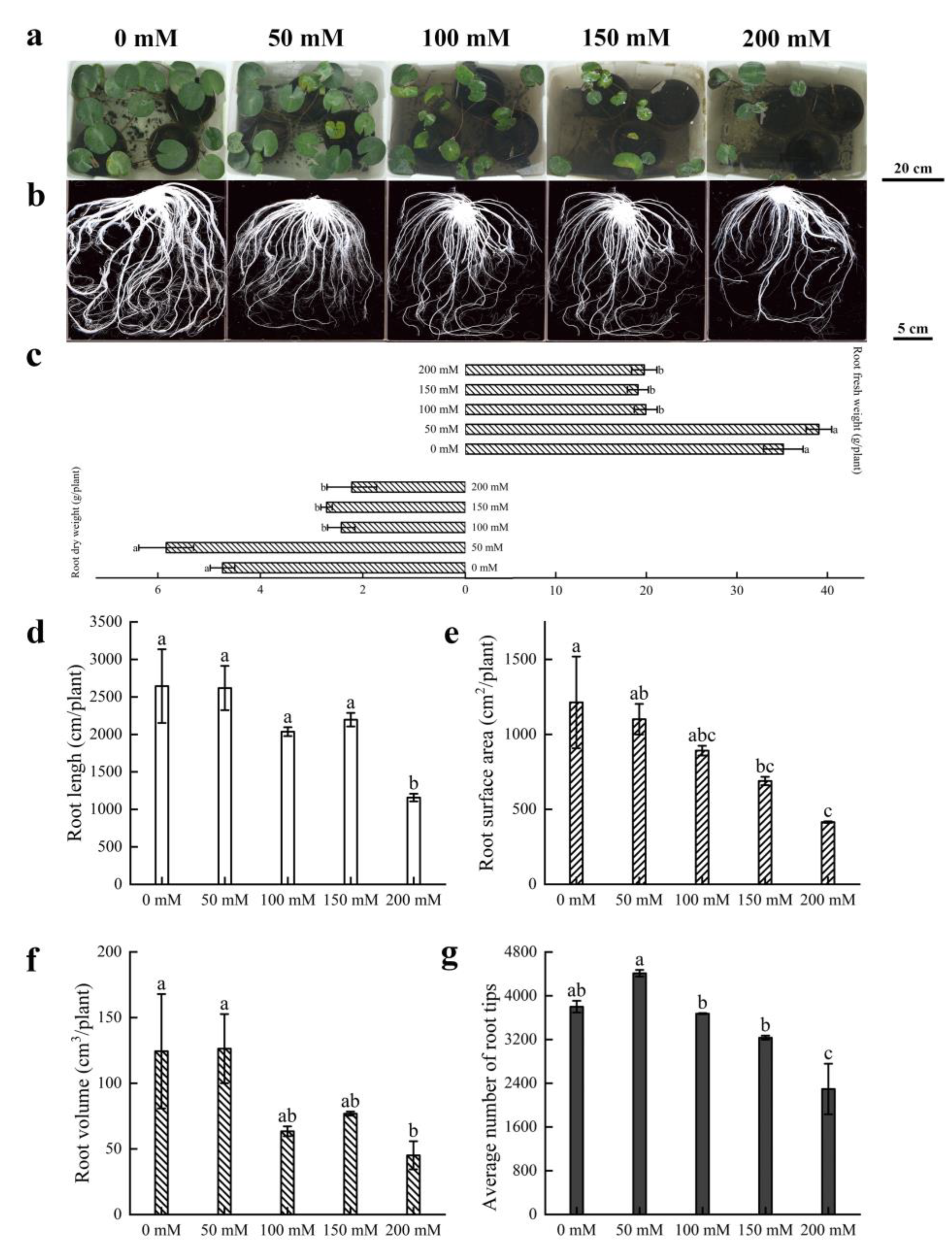
3.1. Morphological Characterization of Water Lily Plants
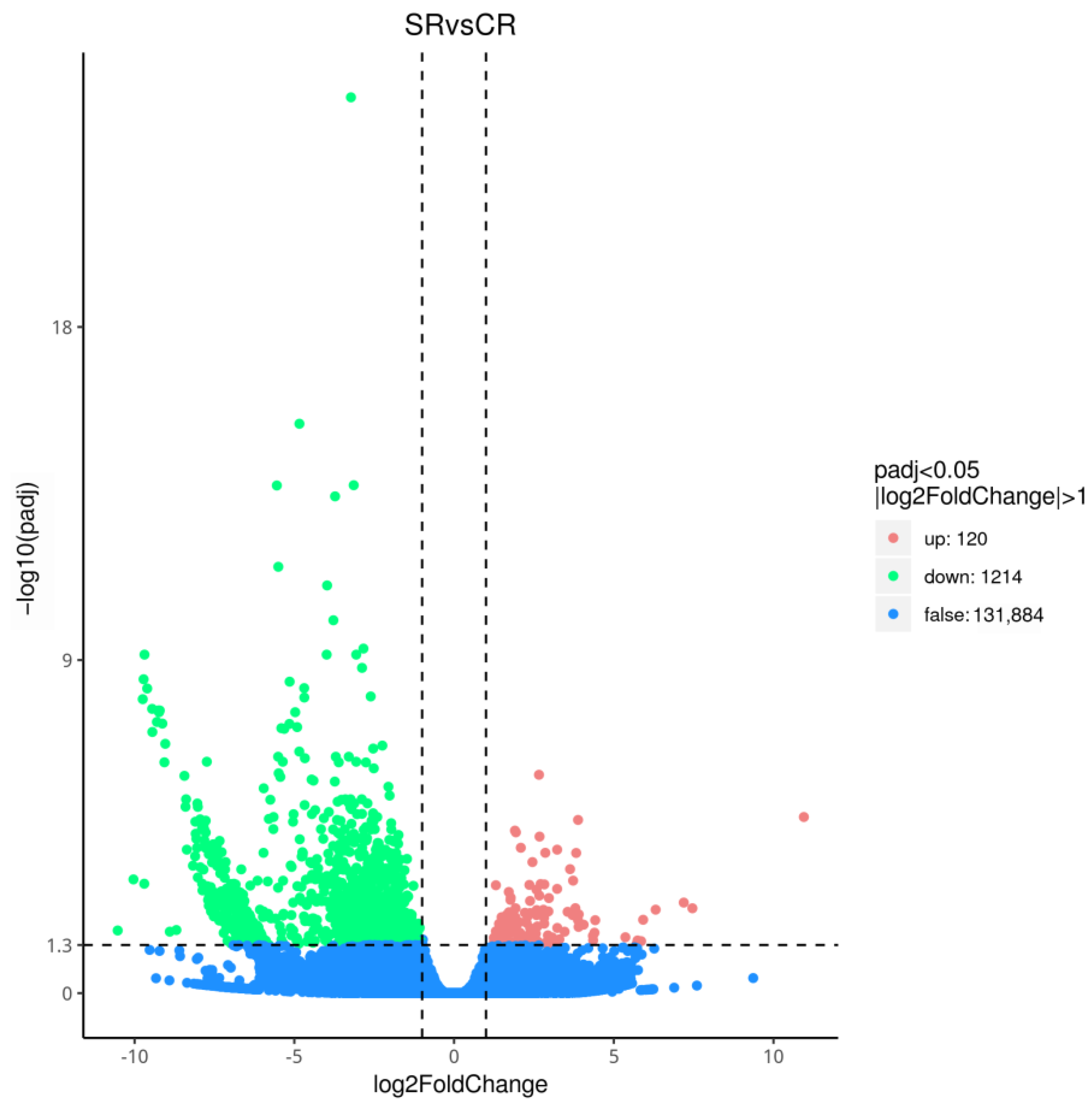
3.2. Analysis of RNA-Seq Data
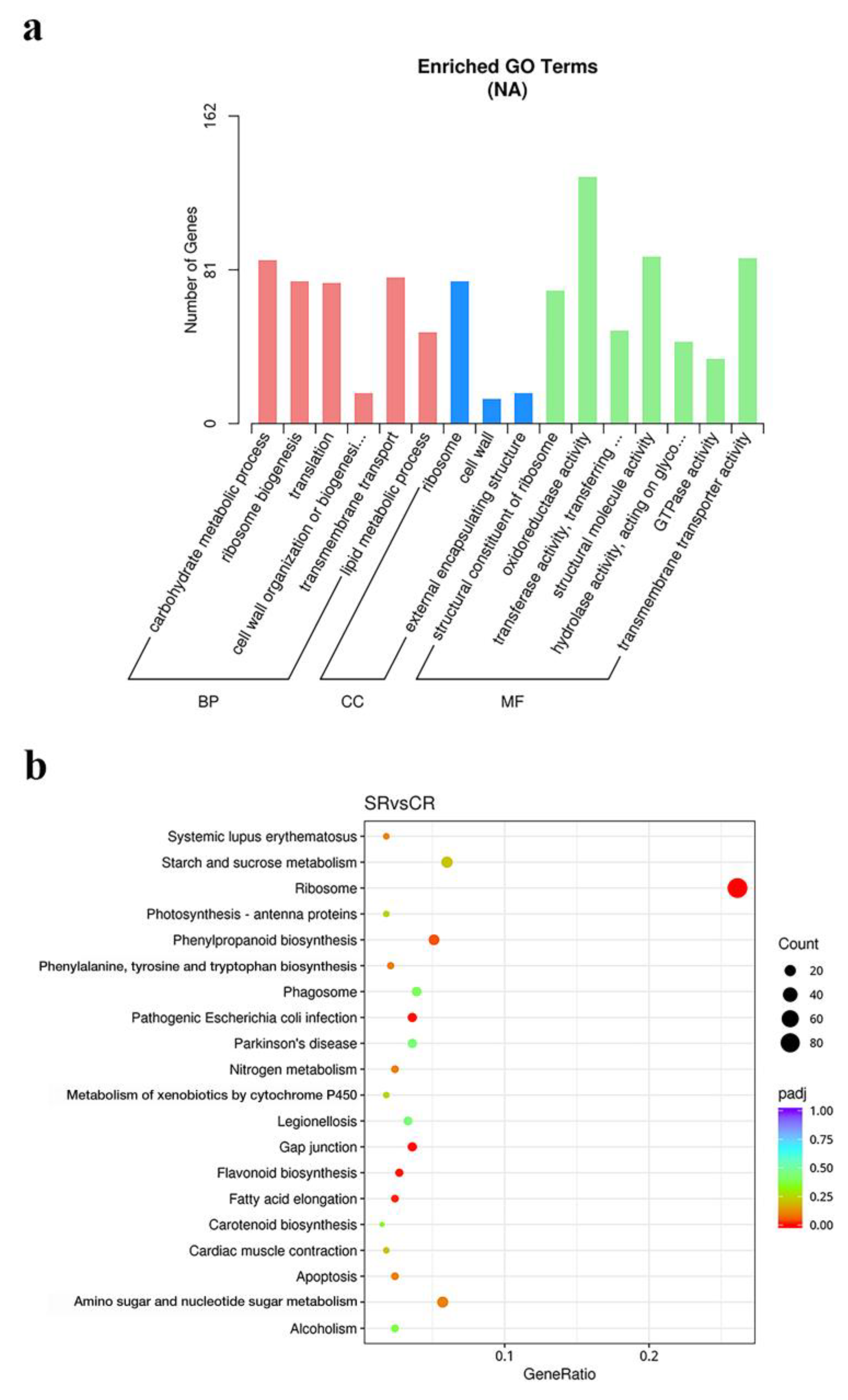
3.3. Pathway Enrichment Analyses
3.4. Analysis of Genes Related to Ion Channels and Transporters
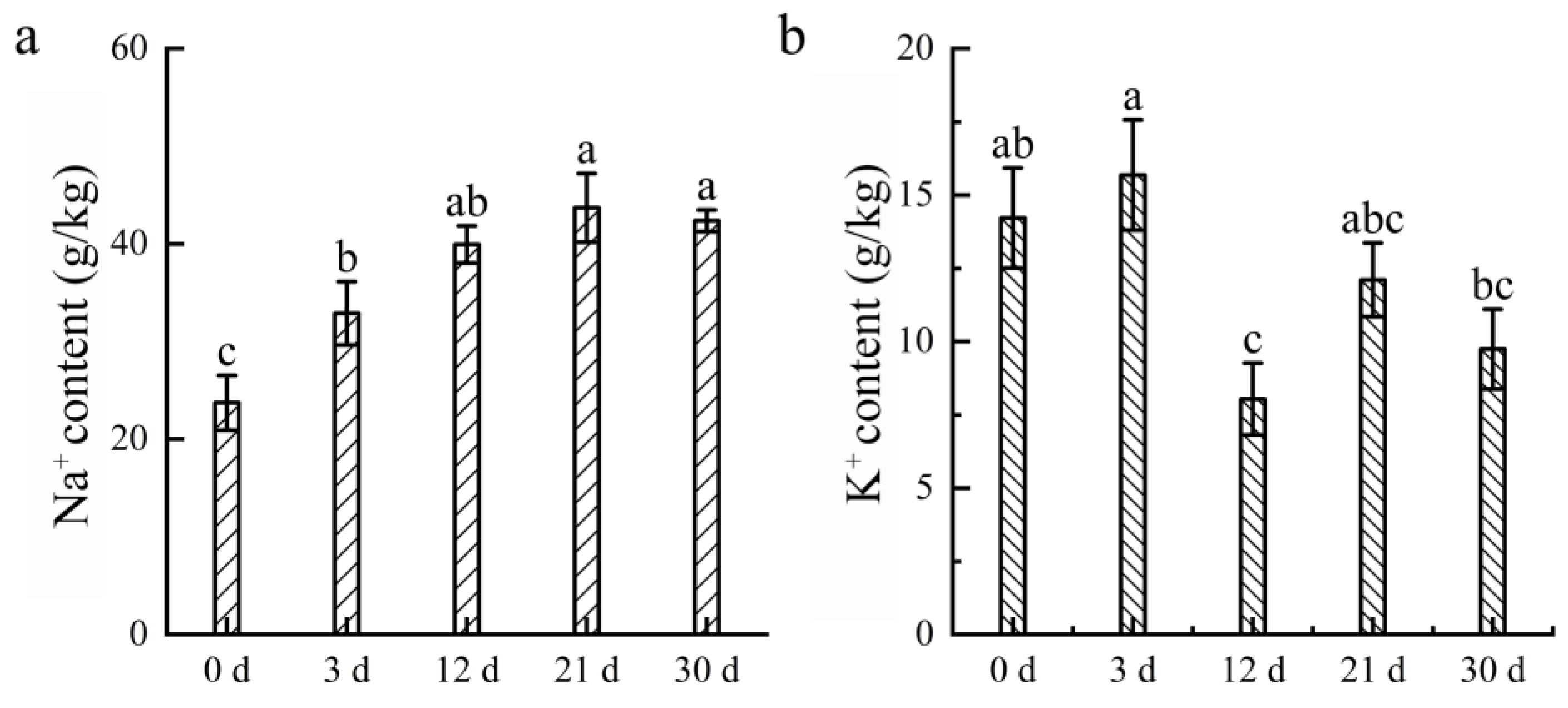
3.5. Analysis of Na+ and K+ in Water Lily Roots
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of Plant Growth-Promoting Rhizobacteria (PGPR) in Stimulating Salinity Stress Defense in Plants: A Review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef]
- Parida, A.; Das, A. Salt Tolerance and Salinity Effects on Plants: A Review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Abiala, M.A.; Abdelrahman, M.; Burritt, D.J.; Tran, L.-S.P. Salt Stress Tolerance Mechanisms and Potential Applications of Legumes for Sustainable Reclamation of Salt-Degraded Soils. Land Degrad. Dev. 2018, 29, 3812–3822. [Google Scholar] [CrossRef]
- Mushtaq, Z.; Faizan, S.; Gulzar, B. Salt Stress, Its Impacts on Plants and the Strategies Plants Are Employing against It: A Review. J. Appl. Biol. Biotechnol. 2020, 8, 81–91. [Google Scholar] [CrossRef]
- Sunarpi, H.T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.; Leung, H.; Hattori, K. Enhanced Salt Tolerance Mediated by AtHKT1 Transporter-Induced Na+ Unloading from Xylem Vessels to Xylem Parenchyma Cells. Plant J. 2005, 44, 928–938. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, S.; Hu, Y.; Wu, F.; Hu, Q.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L. The Role of a Potassium Transporter OsHAK5 in Potassium Acquisition and Transport from Roots to Shoots in Rice at Low Potassium Supply Levels. Plant Physiol. 2014, 166, 945-U757. [Google Scholar] [CrossRef]
- Zhang, K.; Tang, J.; Wang, Y.; Kang, H.; Zeng, J. The Tolerance to Saline-Alkaline Stress Was Dependent on the Roots in Wheat. Physiol. Mol. Biol. Plants 2020, 26, 947–954. [Google Scholar] [CrossRef]
- Chatenet, P.; Froissard, D.; Cook-Moreau, J.; Hourdin, P.; Ghestem, A.; Botineau, M.; Haury, J. Populations of Myriophyllum alterniflorum L. as Bioindicators of Pollution in Acidic to Neutral Rivers in the Limousin Region. Hydrobiologia 2006, 570, 61–65. [Google Scholar] [CrossRef]
- Bennett, S.; Barrett-Lennard, E.; Colmer, T. Salinity and Waterlogging as Constraints to Saltland Pasture Production: A Review. Agric. Ecosyst. Environ. 2009, 129, 349–360. [Google Scholar] [CrossRef]
- Setter, T.; Waters, I.; Sharma, S.; Singh, K.; Kulshreshtha, N.; Yaduvanshi, N.; Ram, P.; Singh, B.; Rane, J.; McDonald, G. Review of Wheat Improvement for Waterlogging Tolerance in Australia and India: The Importance of Anaerobiosis and Element Toxicities Associated with Different Soils. Ann. Bot. 2009, 103, 221–235. [Google Scholar] [CrossRef]
- Babourina, O.; Rengel, Z. Ion Transport in Aquatic Plants. In Waterlogging Signalling and Tolerance in Plants; Mancuso, S., Shabala, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 221–238. ISBN 978-3-642-10304-9. [Google Scholar]
- Garcia-Sanchez, M.; Jaime, M.; Ramos, A.; Sanders, D.; Fernandez, J. Sodium-Dependent Nitrate Transport at the Plasma Membrane of Leaf Cells of the Marine Higher Plant Zostera marina L. Plant Physiol. 2000, 122, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Linares-Rueda, A.; Garcia-Sanchez, M.; Fernandez, J. Physiological Evidence for a Sodium-Dependent High-Affinity Phosphate and Nitrate Transport at the Plasma Membrane of Leaf and Root Cells of Zostera marina L. J. Exp. Bot. 2005, 56, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.-K. The Arabidopsis thaliana Salt Tolerance Gene SOS1 Encodes a Putative Na+/H+ Antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Garciadeblas, B.; Haro, R.; Benito, B. Cloning of Two SOS1 Transporters from the Seagrass Cymodocea nodosa. SOS1 Transporters from Cymodocea and Arabidopsis Mediate Potassium Uptake in Bacteria. Plant Mol. Biol. 2007, 63, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Qiao, G.; Qiu, W.; Yu, D.; Zhou, S.; Shen, Y.; Yu, G.; Jiang, J.; Han, X.; Liu, M. Molecular Breeding of Water Lily: Engineering Cold Stress Tolerance into Tropical Water Lily. Hortic. Res. 2018, 5, 73. [Google Scholar] [CrossRef]
- Luo, H.; Chen, S.; Jiang, J.; Chen, Y.; Chen, F.; Teng, N.; Yin, D.; Huang, C. The Expression of Floral Organ Identity Genes in Contrasting Water Lily Cultivars. Plant Cell Rep. 2011, 30, 1909–1918. [Google Scholar] [CrossRef]
- Hinojosa-Garro, D.; Mason, C.; Underwood, G. Macrophyte Assemblages in Ditches of Coastal Marshes in Relation to Land-Use, Salinity and Water Quality. Fundam. Appl. Limnol. 2008, 172, 325–337. [Google Scholar] [CrossRef]
- Julkowska, M.; Koevoets, I.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.; Keurentjes, J.; Korte, A.; Haring, M.; de Boer, G. Genetic Components of Root Architecture Remodeling in Response to Salt Stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef]
- Korver, R.A.; van den Berg, T.; Meyer, A.J.; Galvan-Ampudia, C.S.; ten Tusscher, K.H.W.J.; Testerink, C. Halotropism Requires Phospholipase D Zeta 1-Mediated Modulation of Cellular Polarity of Auxin Transport Carriers. Plant Cell Environ. 2020, 43, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, X.; Wang, H.; Pan, T.; Wang, Y.; Xu, Y.; Xu, C.; Yang, Z. Genetic Control of Root Plasticity in Response to Salt Stress in Maize. Theor. Appl. Genet. 2021, 134, 1475–1492. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, T.; Zhang, W.; Li, X. SOS3 Mediates Lateral Root Development under Low Salt Stress through Regulation of Auxin Redistribution and Maxima in Arabidopsis. New Phytol. 2011, 189, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Saqib, M.; Zorb, C.; Rengel, Z.; Schubert, S. The Expression of the Endogenous Vacuolar Na+/H+ Antiporters in Roots and Shoots Correlates Positively with the Salt Resistance of Wheat (Triticum aestivum L.). PLANT Sci. 2005, 169, 959–965. [Google Scholar] [CrossRef]
- Sarasketa, A.; Gonzalez-Moro, M.; Gonzalez-Murua, C.; Marino, D. Exploring Ammonium Tolerance in a Large Panel of Arabidopsis thaliana Natural Accessions. J. Exp. Bot. 2014, 65, 6023–6033. [Google Scholar] [CrossRef]
- Tolay, I. The Impact of Different Zinc (Zn) Levels on Growth and Nutrient Uptake of Basil (Ocimum basilicum L.) Grown under Salinity Stress. PLoS ONE 2021, 16, e0246493. [Google Scholar] [CrossRef] [PubMed]
- Coelho, D.G.; Miranda, R.D.; Paula-Marinho, S.D.; de Carvalho, H.H.; Prisco, J.T.; Gomes, E. Ammonium Nutrition Modulates K+ and N Uptake, Transport and Accumulation during Salt Stress Acclimation of Sorghum Plants. Arch. Agron. Soil Sci. 2020, 66, 1991–2004. [Google Scholar] [CrossRef]
- Chebbi, M.; Amdouni, T.; Ben, A.S.; Msilini, N.; Lachaal, M.; Ouerghi, Z. Does the Source of Nitrogen Affect the Response of Fenugreek Plants to Saline Stress? Agrochimica. 2017, 61, 296–316. [Google Scholar] [CrossRef]
- Hessini, K.; Jeddi, K.; Siddique, K.; Cruz, C. Drought and Salinity: A Comparison of Their Effects on the Ammonium-Preferring Species Spartina Alterniflora. Physiol. Plant 2021, 172, 431–440. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Clean Reads | Clean Bases | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| CR1 | 22,947,598 | 22,451,618 | 6.7G | 0.03 | 97.58 | 93.25 | 48.55 |
| CR2 | 22,741,980 | 22,228,034 | 6.7G | 0.03 | 97.72 | 93.55 | 47.91 |
| CR3 | 22,409,365 | 21,936,861 | 6.6G | 0.03 | 97.63 | 93.25 | 48.17 |
| SR1 | 23,281,962 | 21,788,747 | 6.5G | 0.03 | 97.86 | 93.75 | 46.52 |
| SR2 | 22,502,300 | 22,344,183 | 6.7G | 0.03 | 97.89 | 93.92 | 47.56 |
| SR3 | 20,460,810 | 19,022,924 | 5.7G | 0.03 | 97.99 | 94.03 | 43.76 |
| Gene ID | log2FC | p-Value | Description |
|---|---|---|---|
| Downregulated | |||
| Cluster-3509.89148 | −3.11 | 2.67 × 10−3 | sodium transporter HKT1 |
| Cluster-3509.60818 | −4.40 | 9.59 × 10−3 | potassium transporter 5 |
| Cluster-3509.73386 | −2.68 | 5.13 × 10−3 | potassium transporter 5-like |
| Cluster-3509.79079 | −1.72 | 4.51 × 10−2 | potassium transporter 5-like |
| Cluster-3509.78332 | −1.58 | 3.22 × 10−2 | K+ uptake permease 4 |
| Cluster-3509.41717 | −1.34 | 1.31 × 10−2 | boron transporter 2 |
| Cluster-3509.64063 | −1.47 | 2.36 × 10−4 | boron transporter 2 |
| Cluster-3509.65281 | −2.94 | 2.04 × 10−4 | anion transporter 3, chloroplastic |
| Cluster-3509.82188 | −1.25 | 3.10 × 10−2 | organic cation/carnitine transporter 7-like |
| Cluster-3509.72310 | −1.05 | 4.78 × 10−2 | cation-transporting P-type ATPase |
| Cluster-3509.19641 | −2.02 | 7.55 × 10−3 | ABC transporter G family member 36 |
| Cluster-3509.96046 | −3.61 | 9.84 × 10−4 | ABC transporter G family member |
| Cluster-3509.61142 | −1.33 | 3.79 × 10−2 | ABC transporter G family member 29 |
| Cluster-3509.60854 | −5.93 | 8.54 × 10−3 | ABC transporter G family member 21 |
| Cluster-3509.98196 | −2.31 | 1.13 × 10−2 | ABC transporter I family member 17-like |
| Cluster-3509.82719 | −1.14 | 3.20 × 10−2 | ABC transporter I family member 6, chloroplastic |
| Cluster-3509.68592 | −2.68 | 1.59 × 10−2 | ammonium transporter 1 member 1-like |
| Cluster-3509.60927 | −2.84 | 2.33 × 10−2 | ammonium transporter 2-like |
| Cluster-3509.57872 | −2.78 | 1.36 × 10−3 | ammonium transporter 1 member 1-like |
| Cluster-3509.78326 | −4.61 | 4.17 × 10−3 | ammonium transporter 3 member 1-like |
| Cluster-3509.57359 | −3.13 | 8.47 × 10−5 | high-affinity nitrate transporter 2.1 |
| Cluster-3509.63374 | −3.31 | 1.47 × 10−4 | high-affinity nitrate transporter-activating protein 2.1-like |
| Cluster-3509.66911 | −3.23 | 6.24 × 10−25 | urea-proton symporter DUR3 |
| Cluster-3509.51839 | −2.80 | 6.07 × 10−3 | urea-proton symporter DUR3 |
| Cluster-3509.81777 | −1.57 | 3.48 × 10−2 | proton-dependent oligopeptide transporter family |
| Cluster-3509.18628 | −6.08 | 5.55 × 10−3 | proton-dependent oligopeptide transporter family |
| Cluster-3509.40878 | −1.70 | 7.53 × 10−3 | oligopeptide transporter 4 |
| Cluster-3509.77846 | −1.43 | 4.91 × 10−2 | peptide transporter |
| Cluster-3509.97247 | −1.53 | 3.45 × 10−2 | amino acid transporter |
| Cluster-3509.81032 | −1.55 | 6.73 × 10−3 | amino acid/polyamine transporter I |
| Cluster-3509.22635 | −1.91 | 3.95 × 10−2 | cationic amino acid transporter 1 |
| Upregulated | |||
| Cluster-3509.5272 | 3.56 | 6.35 × 10−3 | aluminum-activated malate transporter |
| Cluster-3509.5272 | 3.56 | 6.35 × 10−3 | aluminum-activated malate transporter |
| Cluster-3509.69281 | 2.29 | 7.37 × 10−3 | NRT1/PTR FAMILY 7.2 |
| Cluster-3509.41673 | 1.51 | 2.56 × 10−2 | amino acid transporter AVT6A-like |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Du, F.; Huang, Q.; Gao, X.; Zhang, Z.; Cui, J.; Chang, Y.; Liu, X.; Yao, D. Morpho-Physiological and Transcriptional Regulation of Root System under Saline Conditions in Nymphaea Plants. Horticulturae 2023, 9, 132. https://doi.org/10.3390/horticulturae9020132
Chen S, Du F, Huang Q, Gao X, Zhang Z, Cui J, Chang Y, Liu X, Yao D. Morpho-Physiological and Transcriptional Regulation of Root System under Saline Conditions in Nymphaea Plants. Horticulturae. 2023; 9(2):132. https://doi.org/10.3390/horticulturae9020132
Chicago/Turabian StyleChen, Shaozhou, Fengfeng Du, Qianhao Huang, Xiaojing Gao, Zhiyuan Zhang, Jian Cui, Yajun Chang, Xiaojing Liu, and Dongrui Yao. 2023. "Morpho-Physiological and Transcriptional Regulation of Root System under Saline Conditions in Nymphaea Plants" Horticulturae 9, no. 2: 132. https://doi.org/10.3390/horticulturae9020132
APA StyleChen, S., Du, F., Huang, Q., Gao, X., Zhang, Z., Cui, J., Chang, Y., Liu, X., & Yao, D. (2023). Morpho-Physiological and Transcriptional Regulation of Root System under Saline Conditions in Nymphaea Plants. Horticulturae, 9(2), 132. https://doi.org/10.3390/horticulturae9020132