
Evolution of Seed Dispersal Modes in the Orchidaceae: Has the Vanilla Mystery Been Solved?
 ,
,
Abstract
1. Background
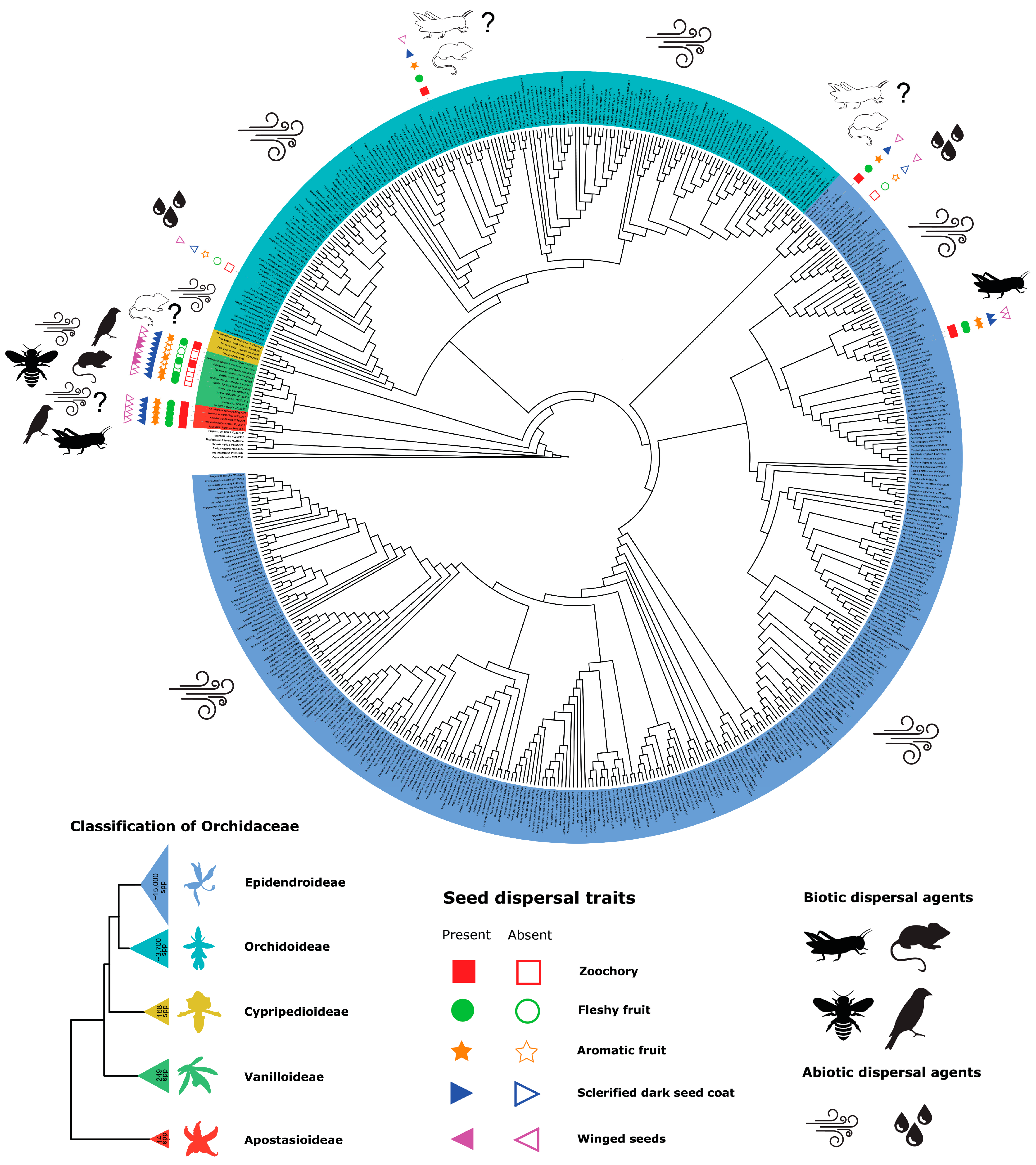
2. Animal-Mediated Seed Dispersal in Orchidaceae
3. Animal-Mediated Seed Dispersal in Vanilla
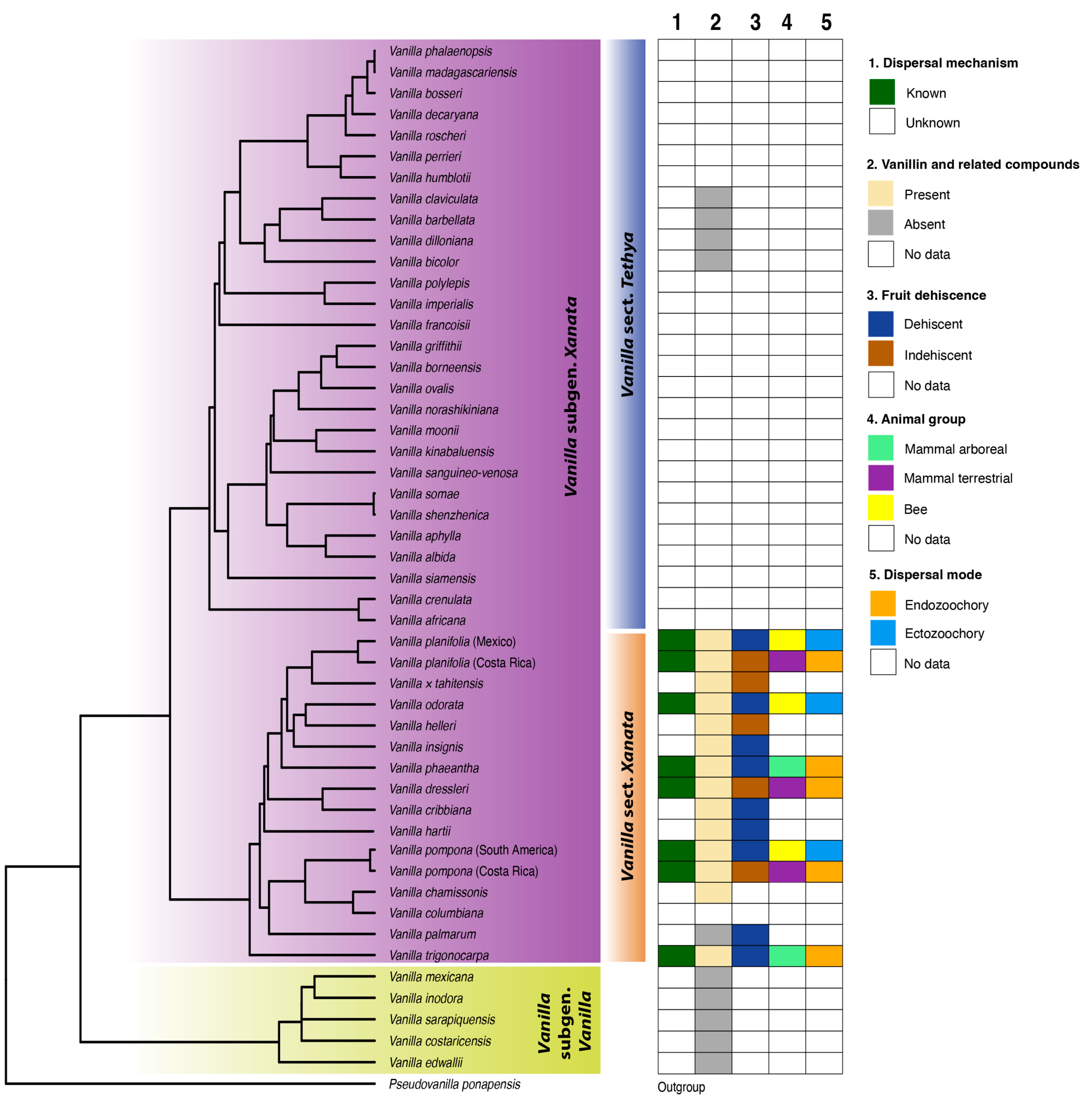
3.1. Vanilla Fruit Trait Diversity
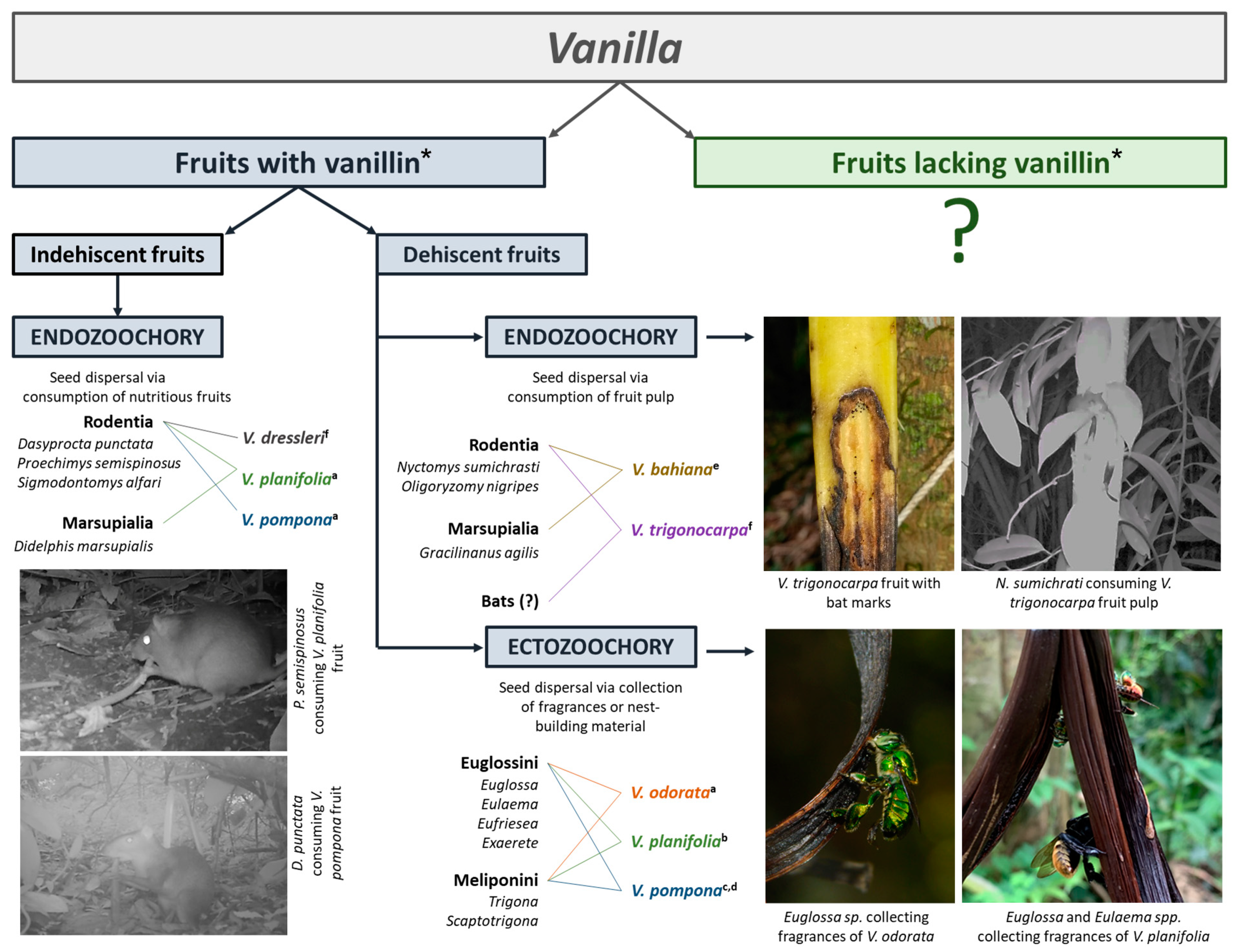
3.2. Linking Vanilla Fruit Traits with Dispersal Modes
3.2.1. Evidence for Ectozoochory
3.2.2. Evidence for Endozoochory
3.3. Flexible Dispersal Modes in Vanilla: A Gene–Environment-Induced Regulation?
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soto Arenas, M.A.; Dressler, R.L. A revision of the Mexican and Central American species of Vanilla Plumier ex Miller with a characterization of their ITS region of the nuclear ribosomal DNA. Lankesteriana 2010, 9, 285–354. [Google Scholar] [CrossRef]
- Cameron, K.M. Vanilla Orchids: Natural History and Cultivation, 1st ed.; Timber Press Inc.: Portland, OR, USA, 2011. [Google Scholar]
- Karremans, A.P.; Chinchilla, I.F.; Rojas-Alvarado, G.; Cedeño-Fonseca, M.; Damián, A.; Léotard, G. A reappraisal of Neotropical Vanilla. With a note on taxonomic inflation and the importance of alpha taxonomy in biological studies. Lankesteriana 2020, 20, 395–497. [Google Scholar] [CrossRef]
- Havkin-Frenkel, D.; Belanger, F.C. Handbook of Vanilla Science and Technology, 2nd ed.; Blackwell Publishing Ltd.: Ames, IA, USA, 2018. [Google Scholar]
- Faostat. Food and Agriculture Organization of the United Nations. 2021. Available online: https://www.fao.org/faostat/ (accessed on 15 June 2023).
- Besse, P.; Da Silva, D.; Bory, S.; Grisoni, M.; Le Bellec, F.; Duval, M.F. RAPD genetic diversity in cultivated vanilla: Vanilla planifolia, and relationships with V. tahitensis and V. pompona. Plant Sci. J. 2004, 167, 379–385. [Google Scholar] [CrossRef]
- Bory, S.; Grisoni, M.; Duval, M.F.; Besse, P. Biodiversity and preservation of vanilla: Present state of knowledge. Genet. Resour. Crop Evol. 2008, 55, 551–571. [Google Scholar] [CrossRef]
- Hernández, H.J.; Lubinsky, P. Cultivation systems. In Vanilla, 1st ed.; Odoux, E., Grisoni, M., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2010; pp. 75–95. [Google Scholar]
- Flanagan, N.S.; Mosquera-Espinosa, A.T. An integrated strategy for the conservation and sustainable use of native Vanilla species in Colombia. Lankesteriana 2016, 16, 201–218. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather. Clim. Extremes 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Bramel, P.; Frey, F. Global Strategy for the Conservation and Use of Vanilla Genetic Resources; Global Crop Diversity Trust: Bonn, Germany, 2021. [Google Scholar]
- Duval, M.F.; Bory, S.; Andrzejewski, S.; Grisoni, M.; Besse, P.; Causse, S.; Charon, C.; Dron, M.; Odoux, E.; Wong, M. Diversité Génétique des Vanilliers Dans Leurs Zones de Dispersion Secondaire. Les Actes BRG 2006, 6, 181–196. [Google Scholar]
- Schlüter, P.M.; Arenas, M.A.S.; Harris, S.A. Genetic variation in Vanilla planifolia (Orchidaceae). Econ. Bot. 2007, 61, 328–336. [Google Scholar] [CrossRef]
- Sreedhar, R.V.; Venkatachalam, L.; Roohie, K.; Bhagyalakshmi, N. Molecular analyses of Vanilla planifolia cultivated in India using RAPD and ISSR markers. OSB 2007, 1, 29–33. [Google Scholar]
- Minoo, D.; Jayakumar, V.N.; Veena, S.S.; Vimala, J.; Basha, A.; Saji, K.V.; Nirmal Babu, K.; Peter, K.V. Genetic variations and interrelationships in Vanilla planifolia and few related species as expressed by RAPD polymorphism. Genet. Resour. Crop. Evol. 2008, 55, 459–470. [Google Scholar] [CrossRef]
- Verma, P.C.; Chakrabarty, D.; Jena, S.N.; Mishra, D.K.; Singh, P.K.; Sawant, S.V.; Tuli, R. The extent of genetic diversity among Vanilla species: Comparative results for RAPD and ISSR. Ind. Crops. Prod. 2009, 29, 581–589. [Google Scholar] [CrossRef]
- Hu, Y.; Resende, M.F., Jr.; Bombarely, A.; Brym, M.; Bassil, E.; Chambers, A.H. Genomics-based diversity analysis of Vanilla species using a Vanilla planifolia draft genome and Genotyping-By-Sequencing. Sci. Rep. 2019, 9, 3416. [Google Scholar] [CrossRef]
- Chambers, A.; Cibrián-Jaramillo, A.; Karremans, A.P.; Martinez, D.M.; Hernandez-Hernandez, J.; Brym, M.; Resende, M.F.R.; Moloney, R.; Sierra, S.N.; Hasing, T.; et al. Genotyping-By-Sequencing diversity analysis of international Vanilla collections uncovers hidden diversity and enables plant improvement. Plant Sci. 2021, 311, 111019. [Google Scholar] [CrossRef] [PubMed]
- Ellestad, P.; Pérez-Farrera, M.A.; Buerki, S. Genomic insights into cultivated Mexican Vanilla planifolia reveal high levels of heterozygosity stemming from hybridization. Plants 2022, 11, 2090. [Google Scholar] [CrossRef]
- Favre, F.; Jourda, C.; Grisoni, M.; Piet, Q.; Rivallan, R.; Dijoux, J.-B.; Hascoat, J.; Lepers-Andrzejewski, S.; Besse, P.; Charron, C. A genome-wide assessment of the genetic diversity, evolution and relationships with allied species of the clonally propagated crop Vanilla planifolia Jacks. ex Andrews. Genet. Resour. Crop. Evol. 2022, 69, 2125–2139. [Google Scholar] [CrossRef]
- Anderson, J.D.; Gastelbondo, M.; Chambers, A.H. Diagnostic KASP markers differentiate Vanilla planifolia, V. odorata, V. pompona, and their hybrids using leaf or cured pod tissues. Mol. Biol. Rep. 2023, 50, 707–717. [Google Scholar] [CrossRef]
- Azofeifa-Bolaños, J.B.; Gigant, L.R.; Nicolás-García, M.; Pignal, M.; Tavares-González, F.B.; Hágsater, E.; Salazar-Chávez, G.A.; Reyes-Lopez, D.; Archila-Morales, F.L.; García-García, J.A.; et al. A new Vanilla species from Costa Rica closely related to V. planifolia (Orchidaceae). Eur. J. Taxon. 2017, 284, 1–26. [Google Scholar] [CrossRef]
- Grisoni, M.; Nany, F. The beautiful hills: Half a century of vanilla (Vanilla planifolia Jacks. ex Andrews) breeding in Madagascar. Genet. Resour. Crop Evol. 2021, 68, 1691–1708. [Google Scholar] [CrossRef]
- Divakaran, M.; Babu, K.N.; Peter, K.V. Conservation of Vanilla species, in vitro. Sci. Hortic. 2006, 110, 175–180. [Google Scholar] [CrossRef]
- Lubinsky, P.; Bory, S.; Hernández Hernández, J.; Kim, S.C.; Gómez-Pompa, A. Origins and dispersal of cultivated vanilla (Vanilla planifolia Jacks [Orchidaceae]). Econ. Bot. 2008, 62, 127–138. [Google Scholar] [CrossRef]
- Li, J.; Demesyeux, L.; Brym, M.; Chambers, A.H. Development of species-specific molecular markers in Vanilla for seedling selection of hybrids. Mol. Biol. Rep. 2020, 47, 1905–1920. [Google Scholar] [CrossRef] [PubMed]
- Soto Arenas, M.A. Filogeografia y Recursos Genéticos de las Vainillas de México. 1999. Available online: http://www.conabio.gob.mx/institucion/proyectos/resultados/InfJ101.pdf (accessed on 2 November 2023).
- Gigant, R.; Bory, S.; Grisoni, M.; Besse, P. Biodiversity and evolution in the Vanilla. In The Dynamical Processes of Biodiversity; Case Studies of Evolution and Spatial, Distribution, Grillo, O., Venora, G., Eds.; IntechOpen: Rijeka, Croatia, 2011; pp. 1–26. [Google Scholar]
- Goettsch, B.; Urquiza-Haas, T.; Koleff, P.; Gasman, F.A.; Aguilar-Meléndez, A.; Alavez, V.; Alejandre-Iturbide, G.; Cuevas, F.A.; Pérez, C.A.; Carr, J.A.; et al. Extinction risk of Mesoamerican crop wild relatives. Plants People Planet 2021, 3, 775–795. [Google Scholar] [CrossRef]
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef] [PubMed]
- Lubinsky, P.; Van Dam, M.; Van Dam, A. Pollination of Vanilla and evolution in Orchidaceae. Lindleyana 2006, 75, 926–929. [Google Scholar]
- Householder, E.; Janovec, J.; Balarezo, M.A.; Huinga, M.J.; Wells, J.; Valega, R. Diversity, natural history, and conservation of Vanilla (Orchidaceae) in Amazonian wetlands of Madre de Dios, Peru. J. Bot. Res. Inst. Tex. 2010, 4, 227–243. [Google Scholar]
- Karremans, A.P.; Lehmann, C.P. A highly threatened new species of Vanilla from Costa Rica. Lindleyana 2018, 87, 304–307. [Google Scholar]
- Karremans, A.P.; Pillco Huarcaya, R.; Watteyn, C.; Cedeño-Fonseca, M.; Chinchilla, I.F.; Rojas-Alvarado, G.; López-Morales, M.; Whitworth, A.; Rodríguez-Herrera, B.; Ramírez, S.; et al. Irresistibly fragrant: A key innovation for the long-distance dispersal of wild Vanilla seeds. In Proceedings of the 6th Scientific Conference on Andean Orchids (IISCAO), Medellin, Colombia, 6–8 August 2019. [Google Scholar]
- Karremans, A.P.; Pillco Huarcaya, R.; Watteyn, C.; Cedeño-Fonseca, M.; Chinchilla, I.F.; Rojas-Alvarado, G.; López-Morales, M.; Whitworth, A.; Rodríguez-Herrera, B.; Ramírez, S.; et al. Irresistibly fragrant: The mysterious ecology of Vanilla. In Proceedings of the 7th International Orchid Conservation Congress (IOCC), London, UK, 29 May 2019. [Google Scholar]
- Karremans, A.P.; Pillco Huarcaya, R.; Watteyn, C.; Cedeño-Fonseca, M.; Chinchilla, I.F.; Rojas-Alvarado, G.; López-Morales, M.; Whitworth, A.; Rodríguez-Herrera, B.; Ramírez, S.; et al. Irresistiblemente fragante: La misteriosa ecología de Vanilla. In Proceedings of the 5th Meeting of Mexican Orchidology, Merida, Mexico, 21 November 2019. [Google Scholar]
- Pansarin, E.R. Unravelling the enigma of seed dispersal in Vanilla. Plant Biol. 2021, 23, 974–980. [Google Scholar] [CrossRef]
- Watteyn, C.; Scaccabarozzi, D.; Muys, B.; van der Schueren, N.; van Meerbeek, K.; Guizar Amador, M.F.; Ackerman, J.D.; Cedeño Fonseca, M.; Chinchilla, I.F.; Reubens, B.; et al. Trick or Treat? Pollinator attraction in Vanilla pompona (Orchidaceae). Biotropica 2021, 54, 268–274. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Suetsugu, K. Mammal-mediated seed dispersal in Vanilla: Its rewards and clues to the evolution of fleshy fruits in orchids. Ecology 2022, 103, e3701. [Google Scholar] [CrossRef]
- Karremans, A.P. Demystifying Orchid Pollination: Stories of Sex, Lies and Obsession; Kew Publishing: Richmond, UK, 2023; p. 440. [Google Scholar]
- Karremans, A.P.; Bogarín, D.; Otárola, M.F.; Sharma, J.; Watteyn, C.; Warner, J.; Herrera, B.R.; Chinchilla, I.F.; Carman, E.; Valerio, E.R.; et al. First evidence for multimodal animal seed dispersal in orchids. Curr. Biol. 2023, 33, 364–371. [Google Scholar] [CrossRef]
- Watteyn, C.; Scaccabarozzi, D.; Muys, B.; Reubens, B.; Ackerman, J.D.; Fernández Otárola, M.; Guizar Amador, M.F.; Karremans, A.P. Sweet as Vanilla hartii: Evidence for nectar rewards in Vanilla (Orchidaceae) flowers. Flora 2023, 303, 152294. [Google Scholar] [CrossRef]
- Webb, C.J. The selection of pollen and seed dispersal in plants. Plant Species Biol. 1998, 13, 57–67. [Google Scholar] [CrossRef]
- Dressler, R.L. The Orchids: Natural History and Classification; Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Dressler, R.L. Phylogeny and Classification of the Orchid Family, 1st ed.; Dioscorides Press: Portland, OR, USA, 1993. [Google Scholar]
- Dixon, K.W.; Pate, J.S.; Kuo, J. The Western Australian subterranean orchid Rhizanthella gardneri Rogers, the underground orchid of Western Australia. In Orchid Biology, Reviews and Perspectives 5; Timber Press: Portland, OR, USA, 1990; pp. 37–62. [Google Scholar]
- Cameron, K.M.; Chase, M.W. Seed morphology of the vanilloid orchids. Lindleyana 1998, 13, 148–169. [Google Scholar]
- Molvray, M.; Chase, M.W. Seed Morphology. In Genera Orchidacearum; Pridgeon, A.M., Criib, P.J., Chase, M.W., Ramusen, F.N., Eds.; General Introduction, Apastosiodea, Cypripedioideae; Oxford University Press: Oxford, UK, 1999; pp. 59–66. [Google Scholar]
- Kocyan, A.; Endress, P.K. Floral structure and development of Apostasia and Neuwiedia (Apostasioideae) and their relationships to other Orchidaceae. Int. J. Plant Sci. 2001, 162, 847–867. [Google Scholar] [CrossRef]
- Cameron, K.M. Vanilloideae. In Genera Orchidacearum; Pridgeon, A.M., Cribb, P.J., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2003; Volume 3, pp. 281–334. [Google Scholar]
- Barthlott, W.; Große-Veldmann, B.; Korotkova, N. Orchid seed diversity. Englera 2014, 32, 1–245. [Google Scholar]
- Yang, C.-K.; Lee, Y.-I. The seed development of a mycoheterotrophic orchid, Cyrtosia javanica Blume. Bot. Stud. 2014, 55, 44. [Google Scholar] [CrossRef]
- Suetsugu, K.; Kawakita, A.; Kato, M. Avian seed dispersal in a mycoheterotrophic orchid Cyrtosia septentrionalis. Nat. Plants 2015, 1, 15052. [Google Scholar] [CrossRef]
- Suetsugu, K. Seed dispersal in the mycoheterotrophic orchid Yoania japonica: Further evidence for endozoochory by camel crickets. Plant Biol. 2018, 20, 707–712. [Google Scholar] [CrossRef]
- Suetsugu, K. Independent recruitment of a novel seed dispersal system by camel crickets in achlorophyllous plants. N. Phytol. 2018, 217, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, K.A. Novel seed dispersal mode of Apostasia nipponica could provide some clues to the early evolution of the seed dispersal system in Orchidaceae. Evol. Lett. 2020, 4, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.Y.; Liu, J.; Luo, F.; Lee, Y.I. Seed dispersal in Neuwiedia singapureana: Novel evidence for avian endozoochory in the earliest diverging clade in Orchidaceae. Bot. Stud. 2021, 62, 3. [Google Scholar] [CrossRef] [PubMed]
- Kurzweil, H. Seed morphology in Southern African Orchidoideae (Orchidaceae). Plant Syst. Evol. 1993, 185, 229–247. [Google Scholar] [CrossRef]
- Gosper, C.R.; Stansbury, C.D.; Vivian-Smith, G. Seed dispersal of fleshy-fruited invasive plants by birds: Contributing factors and management options. Divers. Distrib. 2005, 11, 549–558. [Google Scholar] [CrossRef]
- Duan, Q.; Goodale, E.; Quan, R.-C. Bird fruit preferences match the frequency of fruit colours in tropical Asia. Sci. Rep. 2014, 4, 5627. [Google Scholar] [CrossRef] [PubMed]
- Bouetard, A.; Lefuvre, P.; Gigant, R.; Bory, S.; Pignal, M.; Besse, P.; Grisoni, M. Evidence of transoceanic dispersion of the genus Vanilla based on plastid DNA phylogenetic analysis. Mol. Phylogenet. Evol. 2010, 55, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, T.A.; Furtado, M.S.C.; Pereira, W.C.; Barbarena, F.F.V.A. Vanilla bahiana Hoehne (Orchidaceae): Studies on fruit development and new perspectives into crop improvement for the Vanilla planifolia group. Biota Neotrop. 2019, 19, e20180696. [Google Scholar] [CrossRef]
- Koch, A.K.; de Fraga, C.N.; dos Santos, J.U.M.; Ilkiu-Borges, A.L. Taxonomic notes on Vanilla (Orchidaceae) in the Brazilian Amazon, and the description of a new species. Syst. Bot. 2013, 38, 975–981. [Google Scholar] [CrossRef]
- Pramanik, D.; Becker, A.; Roessner, C.; Rupp, O.; Bogarín, D.; Pérez-Escobar, O.A.; Dirks-Mulder, A.; Droppert, K.; Kocyan, A.; Smets, E.; et al. Evolution and development of fruits of Erycina pusilla and other orchid species. PLoS ONE 2023, 18, e0286846. [Google Scholar] [CrossRef]
- Mühlhausen, A.; Polster, A.; Theißen, G.; Mummenhoff, K. Evolution of fruit dehiscence in Brassicaceae–Examples from Aethionema and Lepidium. Acta Hortic. 2010, 867, 207–220. [Google Scholar] [CrossRef]
- Mühlhausen, A.; Lenser, T.; Mummenhoff, K.; Theißen, G. Evidence that an evolutionary transition from dehiscent to indehiscent fruits in Lepidium (Brassicaceae) was caused by a change in the control of valve margin identity genes. Plant J. 2013, 73, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Gramzow, L.; Klupsch, K.; Fernández-Pozo, N.; Hölzer, M.; Marz, M.; Rensing, S.A.; Theißen, G. Comparative transcriptomics identifies candidate genes involved in the evolutionary transition from dehiscent to indehiscent fruits in Lepidium (Brassicaceae). BMC Plant Biol. 2022, 22, 340. [Google Scholar] [CrossRef]
- Lozano Rodríguez, M.A.; Rodríguez, M.L.; Canche, J.M.P.; García, R.A.M.; Cabrera, C.R.C. Visit frequency of Euglossine bees (Hymenoptera: Apidae) to mature fruits of Vanilla planifolia (Orchidaceae). Acta Bot. Mex. 2022, 129, e2001. [Google Scholar] [CrossRef]
- Madison, M. Vanilla beans and bees. Mo. Bot. Gard. bull. 1981, 8, 8. [Google Scholar]
- Nemésio, A.; Seixas, D.P.; Rasmussen, C. Trigona pallens (Fabricius, 1798) (Hymenoptera: Apidae) strongly attracted to vanillin in northeastern Peru. Braz. J. Biol. 2013, 73, 677–678. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roubik, D.W.; de Camargo, J.M.F. The Panama microplate, island studies and relictual species of Melipona (Melikerria) (Hymenoptera: Apidae: Meliponini). Syst. Entomol. 2013, 37, 189–199. [Google Scholar] [CrossRef]
- Wallace, H.M.; Trueman, S.J. Dispersal of Eucalyptus torelliana seeds by the resin-collecting stingless bee, Trigona carbonaria. Oecologia 1995, 104, 12–16. [Google Scholar] [CrossRef]
- Bacelar-Lima, C.G.; Freire, D.C.B.; Coletto-Silva, A.; Costa, K.B.D.; Laray, J.P.B.; Vilas-Boas, H.C.; Carvalho-Zilse, G.A. Melitocory of Zygia racemosa (Ducke) Barneby & Grimes by Melipona seminigra merrillae Cockerell 1919 and Melipona compressipes manaosensis Schwarz 1932 (Hymenoptera, Meliponina) in Central Amazon, Brazil. Acta Amazon. 2006, 36, 343–348. [Google Scholar]
- Nunez, C.V.; de Oliveira, M.L.; Lima, R.D.; Diaz, I.E.C.; Sargentini, E., Jr.; Pereira, O.L., Jr.; Araújo, L.M. Chemical analyses confirm a rare case of seed dispersal by bees. Apidologie 2008, 39, 618–626. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Baumann, A.M.; Wallace, H.M.; Brooks, P.; Schmitt, T. The chemistry of an unusual seed dispersal mutualism: Bees use a complex set of olfactory cues to find their partner. Anim. Behav. 2014, 98, 41–51. [Google Scholar] [CrossRef]
- Flanagan, N.S.; Otero, J.T.; Molineros, F.H.; Mosquera-Espinosa, A.T.; Vásquez, E.; Garnica, S.; Rosero, D.; Parra, S.; Martínez, C.; Palma, R. Aprovechamiento Sostenible de Recursos Biológicos Nativos del Distrito de Manejo Integrado de Atuncela; Pontificia Universidad Javeriana Cali: Cali, Colombia, 2012; p. 11. [Google Scholar]
- Hartwig, G. The Tropical World: A Popular Scientificaccount of the Natural History of the Animal and Vegetablekingdoms in the Equatorial Regions; Spottiswoode and Co.: London, UK, 1863; p. 201. [Google Scholar]
- Pohl, J.B.E. Reise Im Innern Von Brasilien; Nabu Press: Vienna, Austria, 1832; pp. 416–417. [Google Scholar]
- Pérez-Silva, A.; Nicolás-García, M.; Petit, T.; Dijoux, J.B.; de los Ángeles Vivar-Vera, M.; Besse, P.; Grisoni, M. Quantification of the aromatic potential of ripe fruit of Vanilla planifolia (Orchidaceae) and several of its closely and distantly related species and hybrids. Eur. Food Res. Technol. 2021, 247, 1489–1499. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Ditta, G.S.; Eshed, Y.; Savidge, B.; Bowman, J.L.; Yanofsky, M.F. SHATTERPROOF MADS-box genes control seed dispersal in Arabidopsis. Nature 2000, 404, 766–770. [Google Scholar] [CrossRef]
- Tamayo-García, R.; Narváez-Zapata, J.A.; Ku-González, A.; Aguilar-Espinosa, M.; Gutiérrez-Pacheco, L.C.; Rivera-Madrid, R. Gene expression analysis during the fruit development in dehiscent and indehiscent Bixa orellana L. accessions. Physiol. Mol. Biol. Plants 2022, 28, 709–718. [Google Scholar] [CrossRef]
- Li, X.R.; Deb, J.; Kumar, S.V.; Østergaard, L. Temperature modulates tissue-specification program to control fruit dehiscence in Brassicaceae. Mol. Plant. 2018, 11, 598–606. [Google Scholar] [CrossRef]
- Giovannoni, J.J. Genetic regulation of fruit development and ripening. Plant Cell 2004, 16, S170–S180. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2013, 65, 4561–4575. [Google Scholar] [CrossRef]
- Dirks-Mulder, A.; Ahmed, I.; uit het Broek, M.; Krol, L.; Menger, N.; Snier, J.; van Winzum, A.; de Wolf, A.; van’t Wout, M.; Zeegers, J.J.; et al. Morphological and molecular characterization of orchid fruit development. Front. Plant Sci. 2019, 10, 137. [Google Scholar] [CrossRef]
- Pramanik, D.; Spaans, M.; Kranenburg, T.; Bogarin, B.; Heijungs, R.; Lens, F.; Smets, E.; Gravendeel, B. Inflorescence lignification of natural species and horticultural hybrids of Phalaenopsis orchids. Sci. Hortic. 2022, 295, 110845. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genus [Subfamily] | Species | Seed Dispersal Mechanism | Fleshy/Aromatic Fruit | Sclerified Seed Coat/Winged | Dispersal Agent | References |
|---|---|---|---|---|---|---|
| Apostasia [Apostasioideae] | Zoochory | Yes/most | Yes/No | Crickets | [49,50,52] | |
| Apostasia [Apostasioideae] | A. nipponica | Zoochory | Yes/Yes | Yes/No | Crickets | [57] |
| Neuwiedia [Apostasioideae] | N. grifithii | Anemochory? | No/No | Yes/No | Wind? | [49,50,52] |
| Neuwiedia [Apostasioideae] | N. veratrifolia | Anemochory? | No/No | Yes/No | Wind? | [49,50,52] |
| Neuwiedia [Apostasioideae] | N. borneensis | Zoochory | Yes/Yes | Yes/No | Birds? | [49,50,52] |
| Neuwiedia [Apostasioideae] | N. singapureana | Zoochory | Yes/Yes | Yes/No | Birds | [58] |
| Neuwiedia [Apostasioideae] | N. zolingeri var. javanica | Zoochory | Yes/Yes | Yes/No | Birds? | [49,50,52] |
| Selenipedium [Cypripedioideae] | S. chica | Zoochory? | Yes/Yes | Yes/No | Unknown animals | [31,46,49,51] |
| Clematepistephium [Vanilloideae] | Anemochory? | No/No | No?/Yes | Wind? | [48,52] | |
| Cyrtosia [Vanilloideae] | Zoochory | Yes/Yes | Yes/No | Birds | [53] | |
| Cyrtosia [Vanilloideae] | C. septentrionalis | Zoochory | Yes/Yes | Yes/No | Frugivorous birds | [48,52,54] |
| Epistephium [Vanilloideae] | Anemochory? | No/No | Yes/Yes | Wind? | [45,48,52] | |
| Erythrorchis [Vanilloideae] | Anemochory? | No/No | Yes/Yes | Wind? | [48,52] | |
| Eriaxis [Vanilloideae] | Anemochory? | No/No | No?/Yes | Wind? | [48,52] | |
| Galeola [Vanilloideae] | Anemochory? | No/No | Yes/Yes | Wind? | [45,48,52] | |
| Pseudovanilla [Vanilloideae] | Anemochory? | No/No | Yes/Yes | Wind? | [48,52] | |
| Vanilla [Vanilloideae] | Zoochory | Yes/Yes | Yes/No | Bees, Crickets, Mammals | [42] | |
| Rhizanthella [Orchidoideae] | Zoochory? | Yes/Yes | Yes/No | Mammals? | [47,52] | |
| Palmorchis [Epidendroideae] | Zoochory? | Yes/Yes | Yes/No | Unknown animals | [31,46,49,51,52] | |
| Yoania [Epidendroideae] | Y. amagiensis | Zoochory | Yes/Yes | Yes/No | Crickets | [55,56] |
| Yoania [Epidendroideae] | Y. japonica | Zoochory | Yes/Yes | Yes/No | Crickets | [55,56] |
| Disa [Epidendroideae] | D. uniflora | Hydrochory | No/No | No/No | Streams | [52,59] |
| Epipactis [Epidendroideae] | E. gigantea | Hydrochory | No/No | No/No | Streams | [52] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karremans, A.P.; Watteyn, C.; Scaccabarozzi, D.; Pérez-Escobar, O.A.; Bogarín, D. Evolution of Seed Dispersal Modes in the Orchidaceae: Has the Vanilla Mystery Been Solved? Horticulturae 2023, 9, 1270. https://doi.org/10.3390/horticulturae9121270
Karremans AP, Watteyn C, Scaccabarozzi D, Pérez-Escobar OA, Bogarín D. Evolution of Seed Dispersal Modes in the Orchidaceae: Has the Vanilla Mystery Been Solved? Horticulturae. 2023; 9(12):1270. https://doi.org/10.3390/horticulturae9121270
Chicago/Turabian StyleKarremans, Adam P., Charlotte Watteyn, Daniela Scaccabarozzi, Oscar A. Pérez-Escobar, and Diego Bogarín. 2023. "Evolution of Seed Dispersal Modes in the Orchidaceae: Has the Vanilla Mystery Been Solved?" Horticulturae 9, no. 12: 1270. https://doi.org/10.3390/horticulturae9121270
APA StyleKarremans, A. P., Watteyn, C., Scaccabarozzi, D., Pérez-Escobar, O. A., & Bogarín, D. (2023). Evolution of Seed Dispersal Modes in the Orchidaceae: Has the Vanilla Mystery Been Solved? Horticulturae, 9(12), 1270. https://doi.org/10.3390/horticulturae9121270