
Effect of Pre-Germination Temperature Regime on Pollen Germination and Fruit Set in Pear, Pyrus bretschneiderilia

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Configuration of Pollen Suspension and Determination of Pollen Germination Rate
2.2. Experiments on the Effect of Incubation Time on the Pollen Germination Rate
2.3. Experiment on the Effect of Pollen Suspension Temperature on the Pollen Germination Rate
2.4. Experiment on the Effect of Pollen Suspension (in the Tank) Pre-Germination Temperature on Fruit Set and Fruit Size
2.5. Data Processing
3. Results
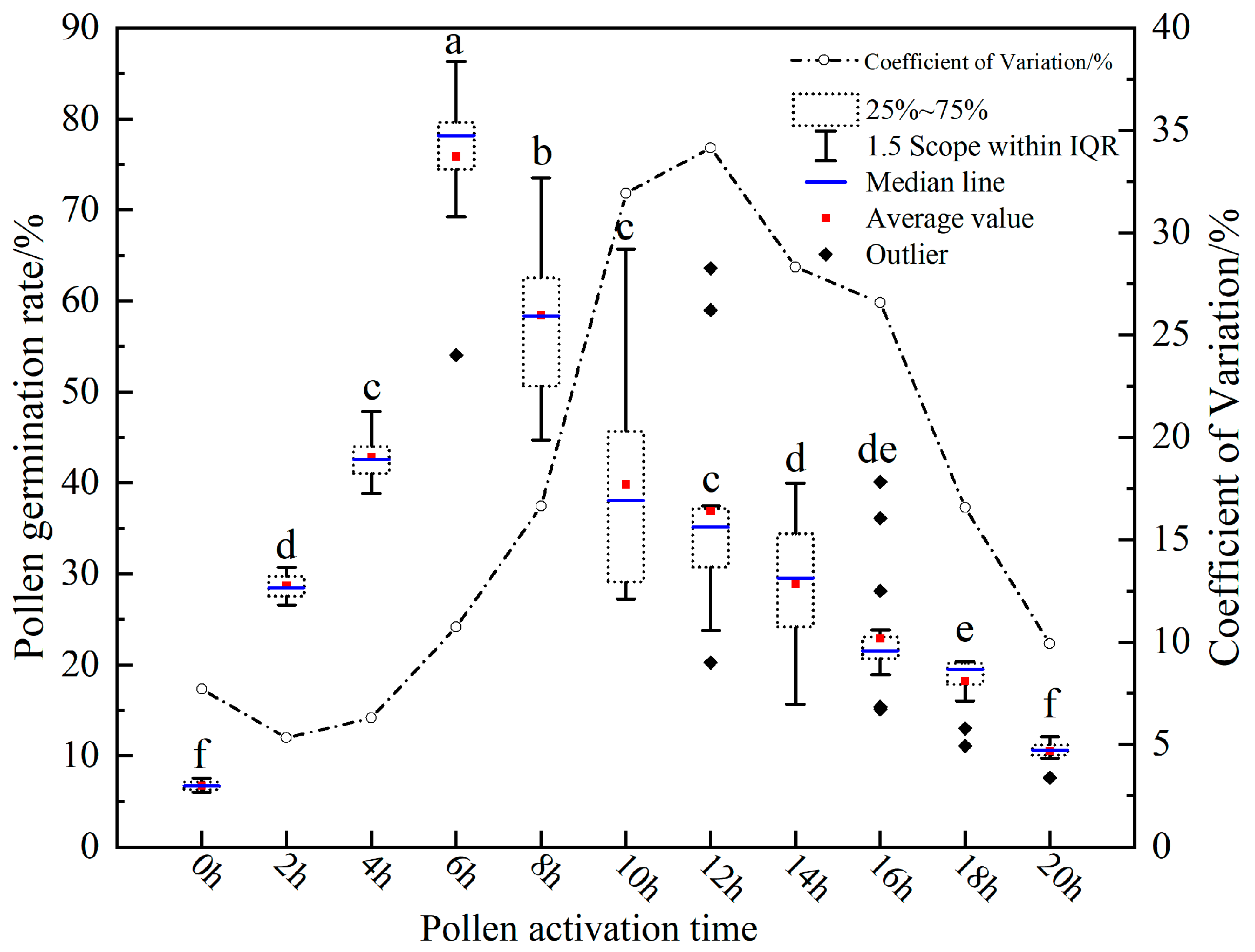
3.1. Results of Incubation Duration on Pollen Germination Rate
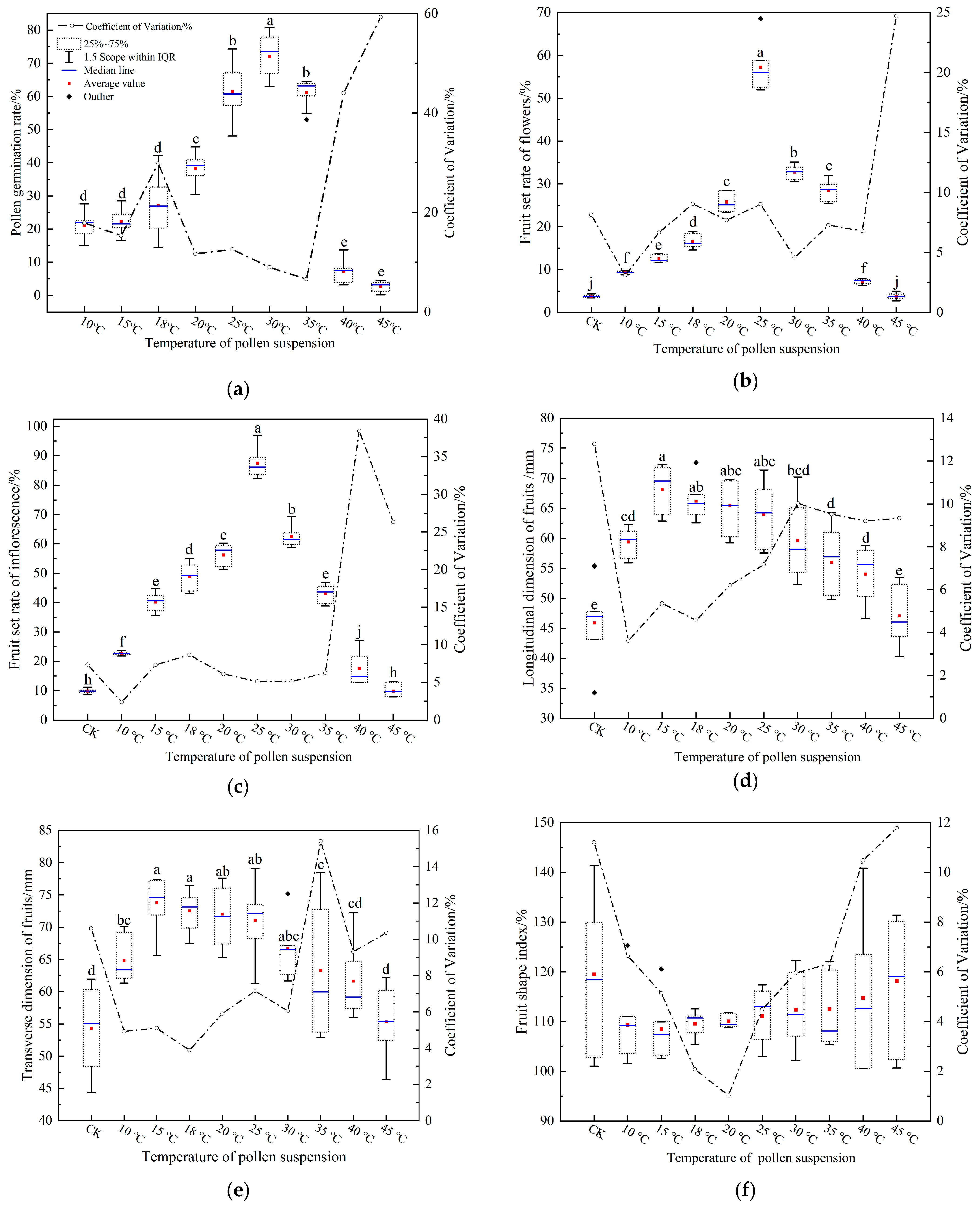
3.2. Effect of Suspension (in the Tank) Temperature on the Germination Rate of Pollen
3.3. Effect of Pollen Suspension Temperature on Fruit Set
3.4. Effect of Pollen Suspension Temperature on Fruit Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garibaldi, L.A.; Carvalheiro, L.G.; Leonhardt, S.D.; Aizen, M.A.; Blaauw, B.R.; Isaacs, R.; Kuhlmann, M.; Kleijn, D.; Klein, A.M.; Kremen, C.; et al. From Research to Action: Enhancing Crop Yield through Wild Pollinators. Front. Ecol. Environ. 2014, 12, 439–447. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatrizfonseca, V.; Ngo, H.T.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; Vanbergen, A.J. Summary for policymakers of the thematic assessment on pollinators, pollination and food production. Biota Neotrop. 2016, 16, e20160101. [Google Scholar] [CrossRef]
- Vasilakakis, M.; Porlingis, I.C. Effect of temperature on pollen germination, pollen tube growth, effective pollination period, and fruit set of pear. Hortscience 1985, 20, 733–735. [Google Scholar] [CrossRef]
- Ren, Z.; Zhao, Y.; Liang, H.; Tao, Z.; Tang, H.; Zhang, H.; Wang, H. Pollination Ecology in China from 1977 to 2017. Plant Divers. 2018, 40, 172–180. [Google Scholar] [CrossRef]
- Ryalls, J.M.W.; Langford, B.; Mullinger, N.J.; Bromfield, L.M.; Nemitz, E.; Pfrang, C.; Girling, R.D. Anthropogenic Air Pollutants Reduce Insect-Mediated Pollination Services. Environ. Pollut. 2022, 297, 118847. [Google Scholar] [CrossRef]
- Yousefi, M.; Mohd Rafie, A.S.; Abd Aziz, S.; Azrad, S.; Razak, A.b.A. Introduction of Current Pollination Techniques and Factors Affecting Pollination Effectiveness by Elaeidobius kamerunicus in Oil Palm Plantations on Regional and Global Scale: A Review. S. Afr. J. Bot. 2020, 132, 171–179. [Google Scholar] [CrossRef]
- Reitmayer, C.M.; Ryalls, J.M.W.; Farthing, E.; Jackson, C.W.; Girling, R.D.; Newman, T.A. Acute Exposure to Diesel Exhaust Induces Central Nervous System Stress and Altered Learning and Memory in Honey Bees. Sci. Rep. 2019, 9, 5793. [Google Scholar] [CrossRef]
- Wurz, A.; Grass, I.; Tscharntke, T. Hand Pollination of Global Crops—A Systematic Review. Basic Appl. Ecol. 2021, 56, 299–321. [Google Scholar] [CrossRef]
- Shi, F.; Jiang, Z.; Ma, C.; Zhu, Y.; Liu, Z. Controlled Parameters of Targeted Pollination Depositionby Air-Liquid Nozzle. Nongye Jixie Xuebao/Trans. Chin. Soc. Agric. Mach. 2019, 50, 115–124. [Google Scholar] [CrossRef]
- Mokdad, M.; Bouhafs, M.; Lahcene, B.; Mokdad, I. Ergonomic Practices in Africa: Date Palm Work in Algeria as an Example. Work 2019, 62, 657–665. [Google Scholar] [CrossRef]
- Beltrán, R.; Valls, A.; Cebrián, N.; Zornoza, C.; García Breijo, F.; Reig Armiñana, J.; Garmendia, A.; Merle, H. Effect of temperature on pollen germination for several rosaceae species: Influence of freezing conservation time on germination patterns. PeerJ 2019, 7, e8195. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Davenport, T.L. Apple Pollination: A Review. Sci. Hortic. 2013, 162, 188–203. [Google Scholar] [CrossRef]
- Jalca Zambrano, I.; García Cruzatty, L.C.; Castro Olaya, J.; Villamar Torres, R.; Guachambala Cando, M. Condiciones Óptimas Para Almacenamiento Del Polen de Ochroma Pyramidale. Bosque (Valdivia) 2019, 40, 227–233. [Google Scholar] [CrossRef]
- Jia, W.; Wang, Y.; Guo, Y.; Wang, Z.; Qi, Q.; Yan, S.; Liu, H.; He, S. Characterization of Pollen Germination and Storage of Paeonia Ludlowii. Linye Kexue/Sci. Silvae Sin. 2021, 57, 82–92. [Google Scholar] [CrossRef]
- Mortazavi, S.M.H.; Arzani, K.; Moieni, A. Optimizing Storage and in Vitro Germination of Date Palm (Phoenix dactylifera) Pollen. J. Agric. Sci. Technol. 2010, 12, 181–189. [Google Scholar]
- Soares, T.L.; de Jesus, O.N.; dos Santos-Serejo, J.A.; de Oliveira, E.J. In Vitro Pollen Germination and Pollen Viability in Passion Fruit (Passiflora spp.). Rev. Bras. Frutic. 2013, 35, 1116–1126. [Google Scholar] [CrossRef]
- Wolukau, J.N.; Zhang, S.; Xu, G.; Chen, D. The effect of temperature, polyamines and polyamine synthesis inhibitor on in vitro pollen germination and pollen tube growth of prunus mume. Sci. Hortic. 2004, 99, 289–299. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Y.; Lombardini, L. In Vitro Viability and Germination of Carya Illinoinensis Pollen under Different Storage Conditions. Sci. Hortic. 2020, 275, 109662. [Google Scholar] [CrossRef]
- Sanzol, J.; Rallo, P.; Herrero, M. Stigmatic Receptivity Limits the Effective Pollination Period in “Agua de Aranjuez” Pear. J. Am. Soc. Hortic. Sci. 2003, 128, 458–462. [Google Scholar] [CrossRef]
- Filho, J.G.S.; Torres, A.C.; Nascimento, W.M.; Boiteux, L.S. Tomato Hybrid Seed Production Using Stored Pollen Grains: Temperature and Storage Time Interactions and Their Effects on Pollen Viability. Acta Hortic. 2019, 1249, 201–204. [Google Scholar] [CrossRef]
- Yoder, K.; Yuan, R.; Combs, L.; Byers, R.; Mcferson, J.; Schmidt, T. Effects of Temperature and the Combination of Liquid Lime Sulfur and Fish Oil on Pollen Germination, Pollen Tube Growth, and Fruit Set in Apples. HortScience 2009, 44, 1277–1283. [Google Scholar] [CrossRef]
- Ferreira, M.d.S.; Soares, T.L.; Costa, E.M.R.; da Silva, R.L.; de Jesus, O.N.; Junghans, T.G.; Souza, F.V.D. Optimization of Culture Medium for the in Vitro Germination and Histochemical Analysis of Passiflora spp. Pollen Grains. Sci. Hortic. 2021, 288, 110298. [Google Scholar] [CrossRef]
- Araújo de Oliveira, A.C.; da Silva Lédo, A.; Polek, M.L.; Krueger, R.; Shepherd, A.; Volk, G.M. Optimization of in Vitro Germination and Cryopreservation Conditions for Preserving Date Palm Pollen in the USDA National Plant Germplasm System. Plant Cell Tissue Organ Cult. 2021, 144, 223–232. [Google Scholar] [CrossRef]
- He, J.G.; Cheng, Y.D.; Guan, J.F.; Ge, W.Y.; Zhao, Z. Changes of chlorogenic acid content and its synthesis-associated genes expression in xuehua pear fruit during development. J. Integr. Agric. 2017, 16, 471–477. [Google Scholar] [CrossRef]
- Song, Y.S.; Lee, S.H.; Cho, J.A.; Moon, C.; Seo, D.J.; Jung, W.J. Expression and degradation patterns of chitinase purified from xuehuali (Pyrus bretschneiderilia) pollen. Int. J. Biol. Macromol. Struct. Funct. Interact. 2018, 107 Pt A, 446–452. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Z.; Han, H.; Jiang, Y.; He, X.; Liu, Y.; Wang, D.; Guo, X.; Liang, Z. Influence of Different Liquid Spray Pollination Parameters on Pollen Activity of Fruit Trees—Pear Liquid Spray Pollination as an Example. Horticulturae 2023, 9, 350. [Google Scholar] [CrossRef]
- Sakamoto, D.; Hayama, H.; Ito, A.; Kashimura, Y.; Moriguchi, T.; Nakamura, Y. Spray Pollination as a Labor-Saving Pollination System in Japanese Pear (Pyrus pyrifolia (Burm.f.) Nakai): Development of the Suspension Medium. Sci. Hortic. 2009, 119, 280–285. [Google Scholar] [CrossRef]
- Gao, Y.-B.; Wang, C.-L.; Wu, J.-Y.; Zhou, H.-S.; Jiang, X.-T.; Wu, J.; Zhang, S.-L. Low temperature inhibits pollen tube growth by disruption of both tip-localized reactive oxygen species and endocytosis in pyrus bretschneideri rehd. Plant Physiol. Biochem. 2014, 74C, 255–262. [Google Scholar] [CrossRef]
- Herrero, M.P.; Johnson, R.R. High temperature stress and pollen viability of maize1. Crop Sci. 1980, 20, 796–800. [Google Scholar] [CrossRef]
- Sukhvibul, N.; Hetherington, S.; Whiley, A.; Smith, M.; Vithanage, V. Effect of temperature on pollen germination, pollen tube growth and seed development in mango (Mangifera indica L.). J. Hortic. Sci. Biotechnol. 2000, 75, 214–222. [Google Scholar] [CrossRef]
- Sukhvibul, N.; Whiley, A.W.; Vithanage, V.; Smith, M.K.; Doogan, V.J.; Hetherington, S.E. Effect of temperature on pollen germination and pollen tube growth of four cultivars of mango (Mangifera indica L.). J. Hortic. Sci. Biotechnol. 2000, 75, 214–222. [Google Scholar] [CrossRef]
- Gianni, T.; Vania, M. Artificial Pollination in Kiwifruit and Olive Trees. In Pollination in Plants; IntechOpen Limited: London, UK, 2018. [Google Scholar]
- de Oliveira da Silva, L.F.; Zambon, C.R.; Pio, R.; de Oliveira, A.F.; Gonçalves, E.D. Establishment of growth medium and quantification of pollen grains of olive cultivars in Brazil’s subtropical areas. Bragantia 2015, 75, 26–32. [Google Scholar] [CrossRef]
- Shaukat, A.K.; Perveen, A. In vitro pollen germination capacity of Citrullus lanatus L. (cucurbitaceae). Pak. J. Bot. 2010, 42, 681–684. [Google Scholar]
- Kuroki, K.; Takemura, Y.; Mingfeng, J.; Marumori, H.; Teratani, N.; Matsumoto, K.; Matsumoto, T.; Tamura, F. Pear pollen selection using higher germination properties at low temperatures and the effect on the fruit set and quality of Japanese pear cultivars. Sci. Hortic. 2017, 216, 200–204. [Google Scholar] [CrossRef]
- Sanzol, J.; Rallo, P.; Herrero, M. Asynchronous development of stigmatic receptivity in the pear (Pyrus communis; rosaceae) flower. Am. J. Bot. 2003, 90, 78–84. [Google Scholar] [CrossRef]
- Liu, X.; Pan, Y.; Liu, C.; Ding, Y.; Wang, X.; Cheng, Z.; Meng, H. Cucumber Fruit Size and Shape Variations Explored from the Aspects of Morphology, Histology, and Endogenous Hormones. Plants 2020, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Bramlage, W.J.; Weis, S.A.; Greene, D.W. Observations on the Relationships among Seed Number, Fruit Calcium, and Senescent Breakdown in Apples. HortScience 1990, 25, 351–353. [Google Scholar] [CrossRef]
- Sobral, M.; Larrinaga, A.R.; Guitián, J. Do Seed-Dispersing Birds Exert Selection on Optimal Plant Trait Combinations? Correlated Phenotypic Selection on the Fruit and Seed Size of Hawthorn (Crataegus monogyna). Evol. Ecol. 2010, 24, 1277–1290. [Google Scholar] [CrossRef]
- Maeda, H.; Akagi, T.; Tao, R. Quantitative Characterization of Fruit Shape and Its Differentiation Pattern in Diverse Persimmon (Diospyros kaki) Cultivars. Sci. Hortic. 2018, 228, 41–48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content (%) | Volume or Mass |
|---|---|---|
| Demineralized water | - | Fixed volume to 5 L |
| Sucrose | 10.00 | 0.50 kg |
| Xanthan gum | 0.02 | 1.00 g |
| Calcium gluconate | 0.05 | 2.50 g |
| Boric acid | 0.01 | 0.50 g |
| Sources of Variation for Different Experiments | Sum of Squares | DF | Mean Squares | F | Sig. |
|---|---|---|---|---|---|
| Pollen activation time and pollen germination rate | 54,635.567 | 10 | 5463.557 | 107.676 | 0.000 *** |
| Pollen suspension temperature (PST) and pollen germination rate | 45,095.424 | 8 | 5636.928 | 186.270 | 0.000 *** |
| PST and fruit set rate of flowers | 17,717.643 | 9 | 1968.627 | 410.933 | 0.000 *** |
| PST and fruit set rate of inflorescences | 40,174.583 | 9 | 4463.843 | 362.652 | 0.000 *** |
| PST and fruit longitudinal dimension | 40,174.583 | 9 | 4463.843 | 362.652 | 0.000 *** |
| PST and fruit transverse dimension | 3074.531 | 9 | 341.615 | 10.741 | 0.000 *** |
| PST and fruit shape index | 906.996 | 9 | 100.777 | 1.238 | 0.290 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Liu, Z.; Han, H.; Qiao, B.; Li, Y.; He, X.; Liu, Y. Effect of Pre-Germination Temperature Regime on Pollen Germination and Fruit Set in Pear, Pyrus bretschneiderilia. Horticulturae 2023, 9, 1151. https://doi.org/10.3390/horticulturae9101151
Liu L, Liu Z, Han H, Qiao B, Li Y, He X, Liu Y. Effect of Pre-Germination Temperature Regime on Pollen Germination and Fruit Set in Pear, Pyrus bretschneiderilia. Horticulturae. 2023; 9(10):1151. https://doi.org/10.3390/horticulturae9101151
Chicago/Turabian StyleLiu, Limin, Ziyan Liu, Hu Han, Baiyu Qiao, Yangfan Li, Xiongkui He, and Yajia Liu. 2023. "Effect of Pre-Germination Temperature Regime on Pollen Germination and Fruit Set in Pear, Pyrus bretschneiderilia" Horticulturae 9, no. 10: 1151. https://doi.org/10.3390/horticulturae9101151
APA StyleLiu, L., Liu, Z., Han, H., Qiao, B., Li, Y., He, X., & Liu, Y. (2023). Effect of Pre-Germination Temperature Regime on Pollen Germination and Fruit Set in Pear, Pyrus bretschneiderilia. Horticulturae, 9(10), 1151. https://doi.org/10.3390/horticulturae9101151