
Tomato Chlorosis Virus (ToCV) Infection Induced the Resistance of Bemisia tabaci to Two Insecticides: Pyrethroids and Flupyradifurone

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Tomato, Cotton and B. tabaci
2.2. Pyrethroids and Flupyradifurone Bioassay of Viruliferous and Non-Viruliferous B. tabaci MED
2.3. Metabolic Enzyme Assays
2.4. Expression Analysis of Detoxification Enzyme Related Genes CYP6CX4, GSTs2 and LOC109038667
2.5. Statistical Analyses
3. Results
3.1. Bioassays
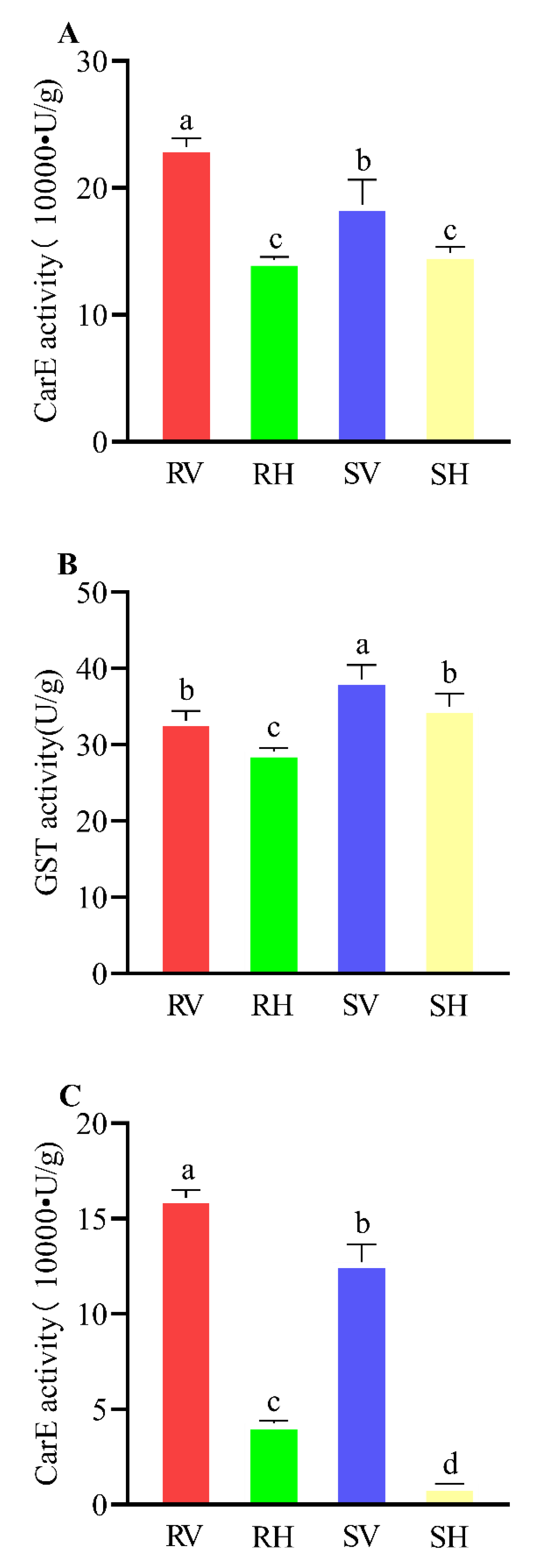
3.2. Activities of Metabolic Enzymes
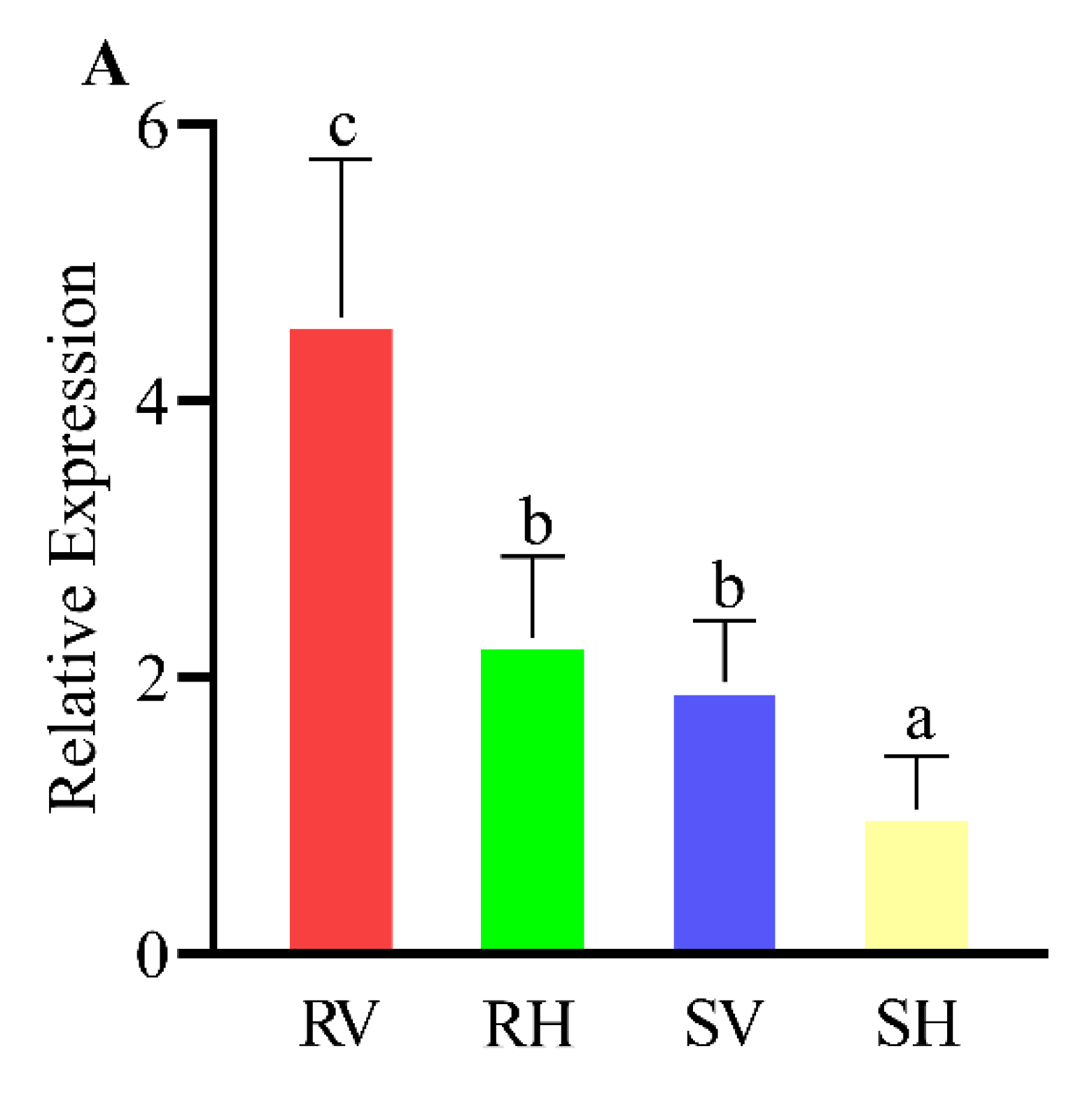
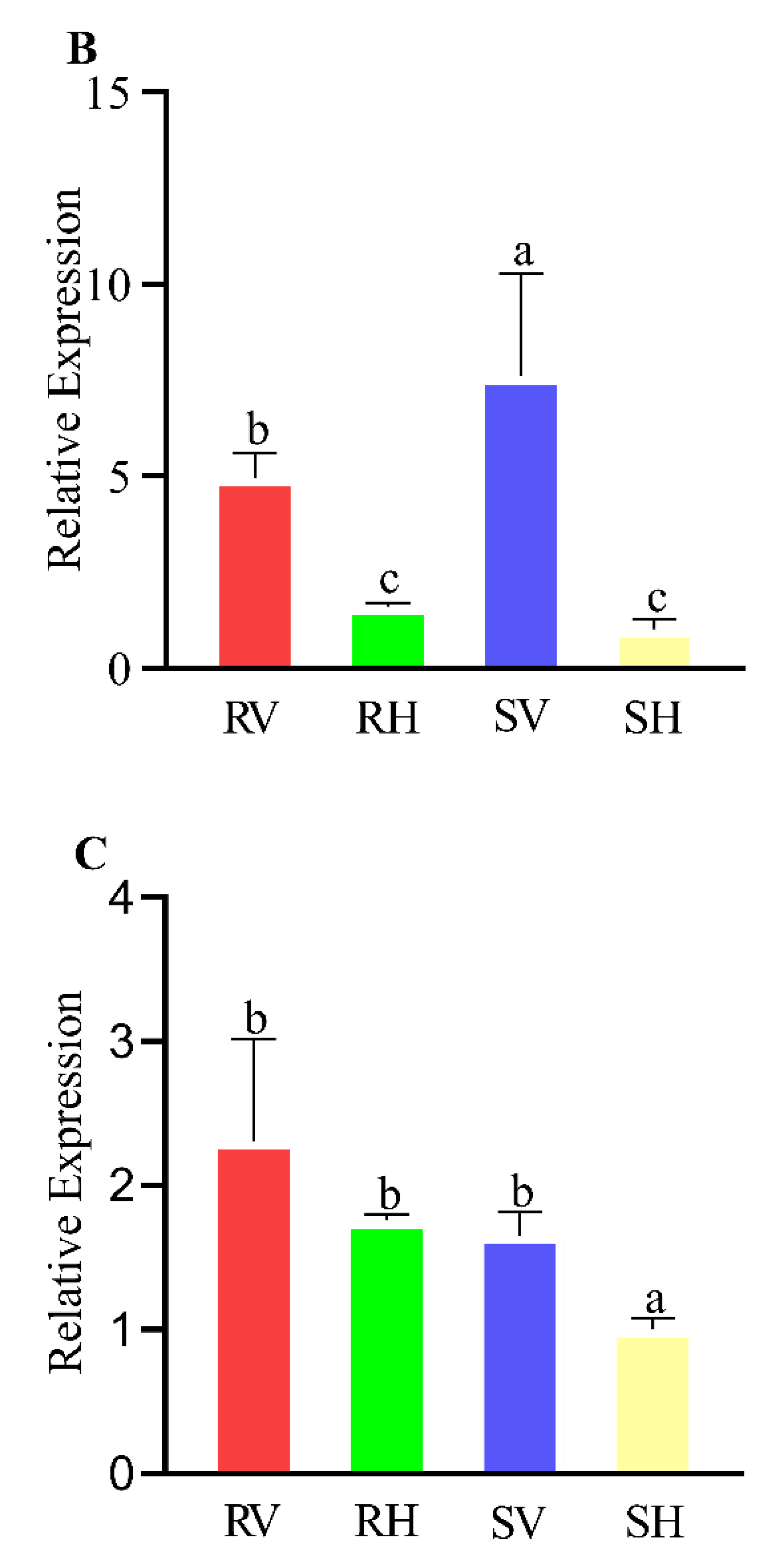
3.3. Expression Patterns of Detoxification-Related P450, GST and CarE
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wisler, G.C.; Li, R.H.; Liu, H.Y.; Lowry, D.S.; Duffus, J.E. Tomato chlorosis virus: A new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopathology 1998, 88, 402–409. [Google Scholar] [CrossRef]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef]
- Accotto, G.P.; Vaira, A.M.; Vecchiati, M.; Sialer, M.M.F.; Gallitelli, D.; Davino, M. First report of Tomato chlorosis virus in Italy. Plant Dis. 2001, 85, 1208. [Google Scholar] [CrossRef]
- Segev, L.; Wintermantel, W.M.; Polston, J.E.; Lapidot, M. First report of Tomato chlorosis virus in Israel. Plant Dis. 2004, 88, 1160. [Google Scholar] [CrossRef]
- Hirota, T.; Natsuaki, T.; Murai, T.; Nishigawa, H.; Niibori, K.; Goto, K.; Hartono, S.; Suastika, G.; Okuda, S. Yellowing disease of tomato caused by Tomato chlorosis virus newly recognized in Japan. J. Gen. Plant Pathol. 2010, 76, 168–171. [Google Scholar] [CrossRef]
- Arruabarrena, A.; Rubio, L.; González-Arcos, M.; Maeso, D.; Fonseca, M.E.N.; Boiteux, L.S. First report of Tomato chlorosis virus infecting tomato crops in Uruguay. Plant Dis. 2014, 98, 1445. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Hamed, A.A.; Moriones, E.; Navas-Castillo, J. First report of Tomato chlorosis virus infecting tomato in Sudan. Plant Dis. 2011, 95, 1592. [Google Scholar] [CrossRef]
- Zhao, R.N.; Wang, R.; Wang, N.; Fan, Z.F.; Zhou, T.; Shi, Y.C.; Chai, M. First report of Tomato chlorosis virus in China. Plant Dis. 2013, 97, 1123. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Wisler, G.C. Vector specificity, host range, and genetic diversity of Tomato chlorosis virus. Plant Dis. 2006, 90, 814–819. [Google Scholar] [CrossRef]
- Wu, T.; Yang, D.Q. Biological characteristics and prevention and treatment of Bemisia tabaci. Hunan Agric. Sci. 2005, 01, 56–58. [Google Scholar]
- Zheng, H.X.; Xie, W.; Fu, B.L.; Xiao, S.; Tan, X.; Ji, Y.; Cheng, J.X.; Wang, R.; Liu, B.M.; Yang, X. Annual analysis of field-evolved insecticide resistance in Bemisia tabaci across China. Pest Manag. Sci. 2021, 77, 2990–3001. [Google Scholar] [CrossRef]
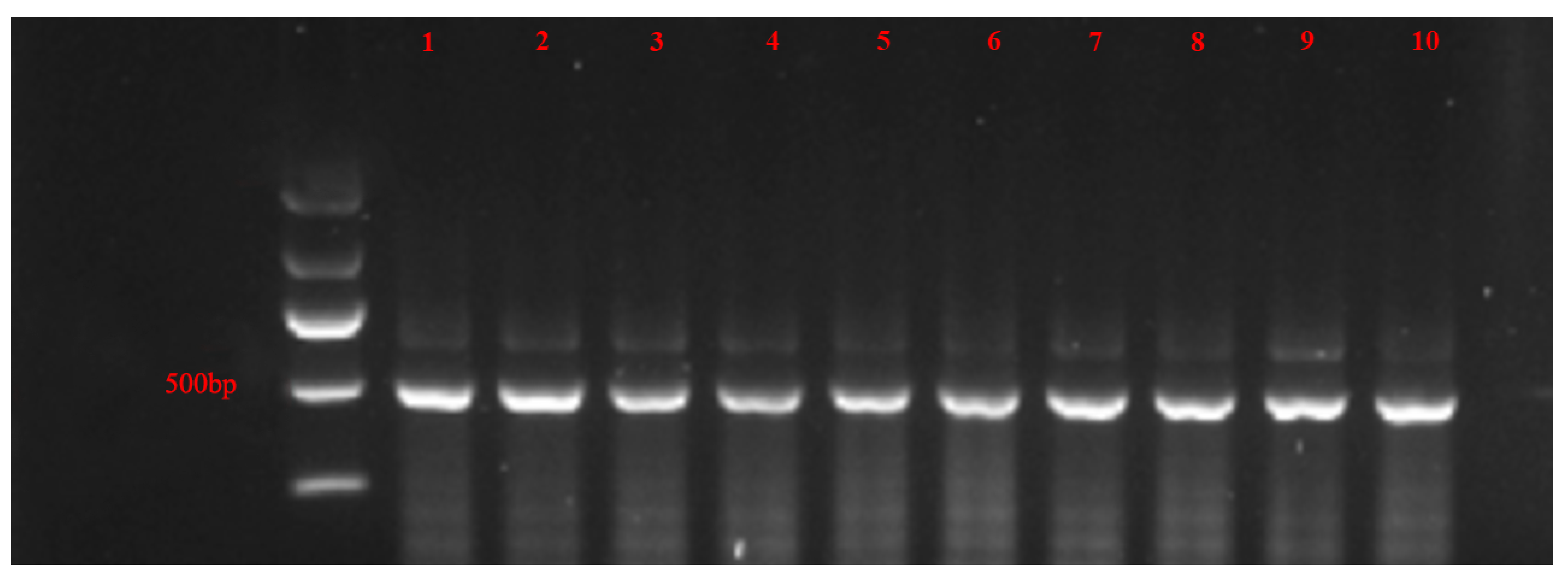
- Shi, X.B.; Tang, X.; Zhang, X.; Zhang, D.Y.; Li, F.; Yan, F.; Zhang, Y.J.; Zhou, X.G.; Liu, Y. Transmission efficiency, preference and behavior of Bemisia tabaci MEAM1 and MED under the influence of Tomato chlorosis virus. Front. Plant Sci. 2018, 8, 2271. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I.; Ahmad, Z.; Denholm, I. Cotton whitefly (Bemisia tabaci) resistance to organophosphate and pyrethroid insecticides in Pakistan. Pest Manag. Sci. 2002, 58, 203–208. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Chi, G.T.; Zhang, J.L. The progress of detoxification enzyme systems and resistance of insecticide. J. Hebei Agric. Univ. 2002, 25, 193–195. [Google Scholar]
- Wang, R.; Wang, J.D.; Zhang, J.S.; Che, W.N.; Feng, H.L.; Luo, C. Characterization of flupyradifurone resistance in the whitefly Bemisia tabaci Mediterranean (Q biotype). Pest Manag. Sci. 2020, 76, 4286–4292. [Google Scholar] [CrossRef]
- Li, X.C.; Schuler, M.R.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- He, Y.X.; Huang, J.; Yang, X.J.; Weng, Q.Y. Pyrethroid resistance mechanisms in Bemisia tabaci (Gennadius). Acta Entomol. Sin. 2007, 50, 241–247. [Google Scholar]
- Orílio, A.F.; Fortes, I.M.; Navas-Castillo, J. Infectious cDNA clones of the crinivirus Tomato chlorosis virus are competent for systemic plant infection and whitefly-transmission. Virology 2014, 464, 365–374. [Google Scholar] [CrossRef]
- Khasdan, V.; Levin, I.; Rosner, A. DNA markers for identifying biotypes B and Q of Bemisia tabaci (Hemiptera: Aleyrodidae) and studying population dynamics. Bull. Entomol. Res. 2005, 95, 605–613. [Google Scholar] [CrossRef]
- Chu, D.; Wan, F.H.; Zhang, Y.J. Change in the biotype composition of Bemisia tabaci in Shandong Province of China from 2005 to 2008. Environ. Entomol. 2010, 39, 1028–1036. [Google Scholar] [CrossRef]
- Chu, D.; Zhang, Y.J.; Wan, F.H. Cryptic Invasion of the Exotic Bemisia tabaci Biotype Q Occurred Widespread in Shandong Province of China. Fla. Entomol. 2010, 93, 203–207. [Google Scholar] [CrossRef]
- Liang, P.; Guo, Y.J.; Zhou, X.G.; Gao, X.W. Expression profiling in Bemisia tabaci under insecticide treatment: Indicating the necessity for custom reference gene selection. PLoS ONE 2014, 9, e87514. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Feng, Y.T.; Wu, Q.J.; Xu, B.Y.; Wang, S.L.; Chang, X.L.; Xie, W.; Zhang, Y.J. Fitness costs and morphological change of laboratory-selected thiamethoxam resistance in the B-type Bemisia tabaci (Hemiptera: Aleyrodidae). J. Appl. Entomol. 2009, 133, 466–472. [Google Scholar] [CrossRef]
- Xu, H.X.; He, X.C.; Zheng, X.S.; Yang, Y.J.; Lu, Z.X. Influence of rice black streaked dwarf virus on the ecological fitness of non-vector planthopper Nilaparvata lugens (Hemiptera: Delphacidae). Insect Sci. 2011, 25, 654–658. [Google Scholar]
- Chen, C.; Jiang, D.C.; Yang, H.; Jin, D.C. Effects of southern rice black streaked dwarf virus on defense enzymes in brown planthopper and white-blacked planthopper. J. Environ. Entomol. 2016, 38, 113–118. [Google Scholar]
- Meng, L.Q.; Li, D.D.; Su, D.; Zhao, H.Y.; Hu, Z.Q. Activities of protective and detoxifying enzymes in spring-grain aphid schizaphis graminum (Hemiptera: Aphididae) fed on wheat infected with barley yellow dwarf virus. J. Plant. Protect. 2019, 46, 707–708. [Google Scholar]
- Yan, M.H.; He, H.F.; Zhang, Z.L.; Zhang, B.B.; Zhu, C.Q.; Yan, W.L. Molecular Basis of Mutual Benefits Between Cucurbit Chlorotic Yellows Virus (CCYV) Transmission and Imidacloprid Resistance in Bemisia tabaci. 2022. Available online: https://www.researchsquare.com/article/rs-1469019/v1 (accessed on 5 November 2022). [CrossRef]
- Su, Q.; Preisser, E.L.; Zhou, X.M.; Xie, W.; Liu, B.M.; Wang, S.L.; Wu, Q.J.; Zhang, Y.J. Manipulation of host quality and defense by a plant virus improves performance of whitefly vectors. J. Econ. Entomol. 2015, 108, 11. [Google Scholar] [CrossRef]
- Li, J.; Ding, T.B.; Chi, H.; Chu, D. Effects of tomato chlorosis virus on the performance of its key vector, Bemisia tabaci, in China. J. Appl. Entomol. 2018, 142, 296–304. [Google Scholar] [CrossRef]
- Mayer, C.J.; Vilcinskas, A.; Gross, J. Phytopathogen lures its insect vector by altering host plant odor. J. Chem. Ecol. 2008, 34, 1045–1049. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Aim | Sequence (5′-3′) |
|---|---|---|
| C1-J-2195 | PCR | TTGATTTTTTGGTCATCCAGAAGT |
| R-BQ-2819 | CTGAATATCGRCGAGGCATTCC | |
| ToCV-3F | PCR | GGTTTGGATTTTGGTACTACATTCAGT |
| ToCV-3R | AAACTGCCTGCATGAAAAGTCTC | |
| Actain- B. tabaci-F | qPCR | ACCGCAAGATTCCATACCC |
| Actain- B. tabaci-R | TAGCCTTGTGCCAATTTCCG | |
| CYP6CX4-F | qPCR | TTGACAAACTTGCGGGGAACCTC |
| CYP6CX4-R | CACAGTCTTTCAGCGTCTCGT | |
| GSTs2-F | qPCR | ATGCCAAAAAATGTGGTCTCGT |
| GSTs2-R | AAATAGGTTGAAATAGCTTGACGC | |
| Gene-Loc109038667-F | qPCR | GCGTCACAACGATCACCTTC |
| Gene-Loc109038667-R | CCGAAGGAAAAGTCACGGAT |
| Pesticide | Stocks | LC50 (ppm) | 95% Confidence Interval |
|---|---|---|---|
| Bifenthrin (pyrethroids) | RV | 414.63 | 312.94~524.34 |
| RH | 359.37 | 276.64~447.69 | |
| SV | 22.55 | 17.77~27.74 | |
| SH | 8.21 | 6.14~10.65 | |
| Flupyradifurone | RV | 70.00 | 38.44~105.75 |
| RH | 51.74 | 36.96~66.85 | |
| SV | 26.34 | 14.68~38.30 | |
| SH | 19.90 | 14.20~25.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Huang, L.; Zhang, Z.; Zhang, Z.; Zhang, D.; Zhang, Y.; Shi, X.; Liu, Y. Tomato Chlorosis Virus (ToCV) Infection Induced the Resistance of Bemisia tabaci to Two Insecticides: Pyrethroids and Flupyradifurone. Horticulturae 2023, 9, 68. https://doi.org/10.3390/horticulturae9010068
Zhang J, Huang L, Zhang Z, Zhang Z, Zhang D, Zhang Y, Shi X, Liu Y. Tomato Chlorosis Virus (ToCV) Infection Induced the Resistance of Bemisia tabaci to Two Insecticides: Pyrethroids and Flupyradifurone. Horticulturae. 2023; 9(1):68. https://doi.org/10.3390/horticulturae9010068
Chicago/Turabian StyleZhang, Jisong, Liping Huang, Zhuo Zhang, Zhanhong Zhang, Deyong Zhang, Youjun Zhang, Xiaobin Shi, and Yong Liu. 2023. "Tomato Chlorosis Virus (ToCV) Infection Induced the Resistance of Bemisia tabaci to Two Insecticides: Pyrethroids and Flupyradifurone" Horticulturae 9, no. 1: 68. https://doi.org/10.3390/horticulturae9010068
APA StyleZhang, J., Huang, L., Zhang, Z., Zhang, Z., Zhang, D., Zhang, Y., Shi, X., & Liu, Y. (2023). Tomato Chlorosis Virus (ToCV) Infection Induced the Resistance of Bemisia tabaci to Two Insecticides: Pyrethroids and Flupyradifurone. Horticulturae, 9(1), 68. https://doi.org/10.3390/horticulturae9010068