
Baseline Sensitivity and Resistance of Botrytis cinerea to Penthiopyrad in Hebei Province, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Fungicides
2.3. Baseline-Sensitivity of B. cinerea to Penthiopyrad
2.3.1. Mycelial Radial Growth Assay
2.3.2. Conidia Germination Test
2.4. Detection of B. cinerea Resistance to Penthiopyrad
2.5. Sequencing of the sdhA, sdhB, sdhC and sdhD Genes
2.6. Correlation between Penthiopyrad and Three Other SDHI Fungicides
2.7. Statistical Analysis
3. Results
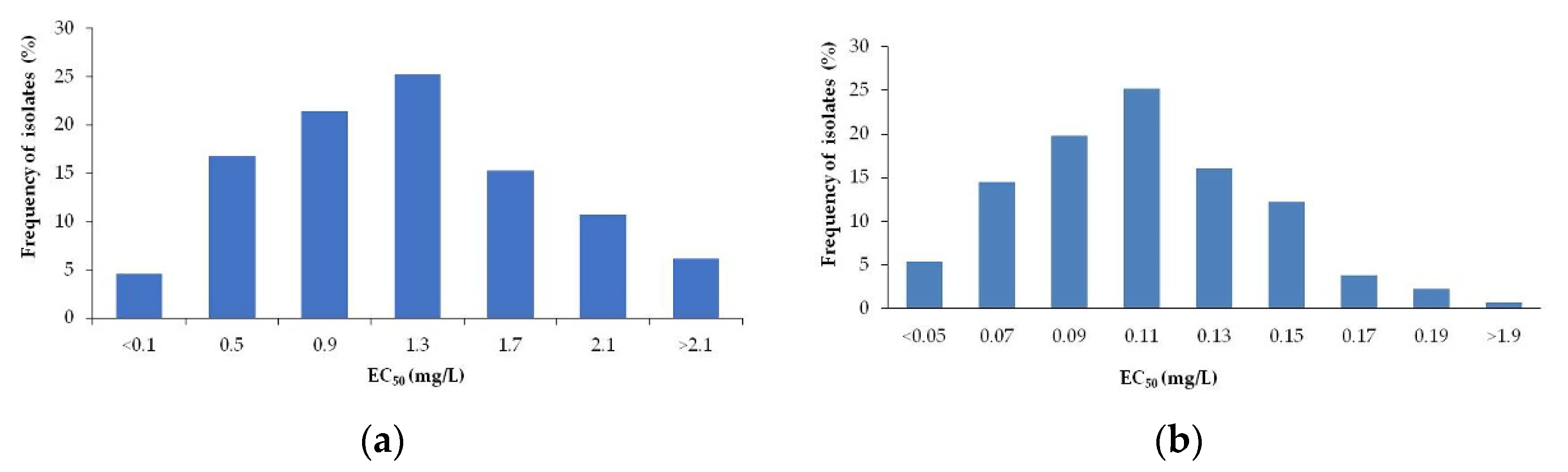
3.1. Sensitivity Baseline of B. cinerea to Penthiopyrad
3.1.1. On Mycelial Growth
3.1.2. On Conidia Germination
3.2. Resistance of B. cinerea to Penthiopyrad
3.3. Sequence Variation of the sdhA, sdhB, sdhC, and sdhD Genes
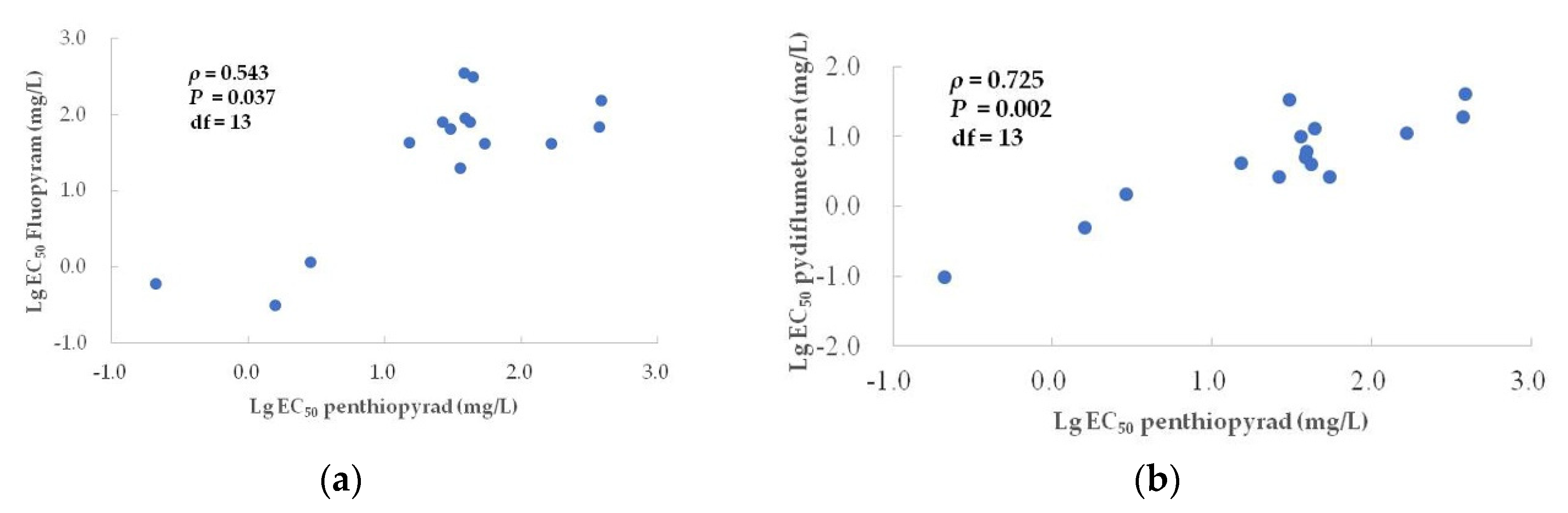
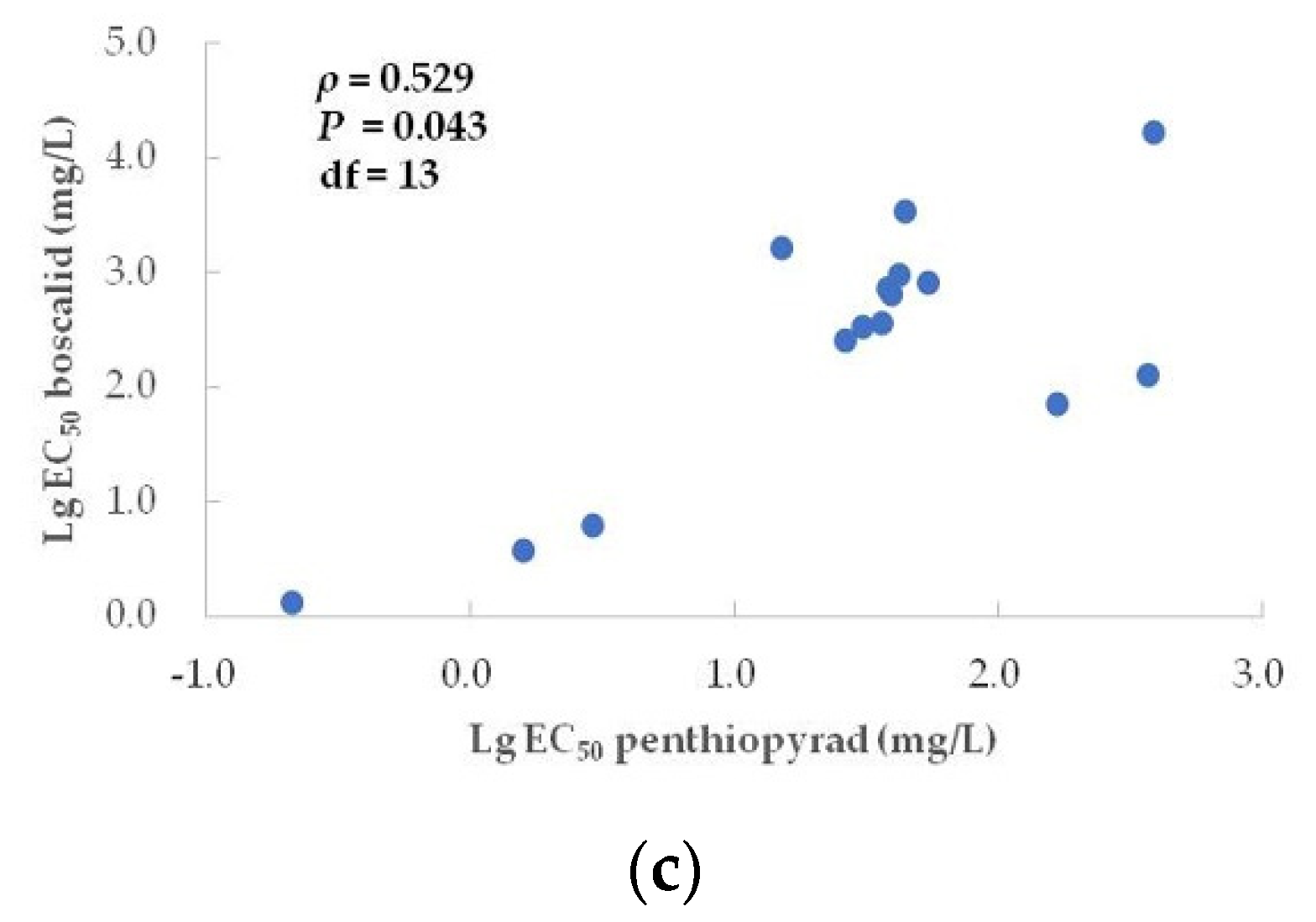
3.4. Correlation between Penthiopyrad and Other Botryticides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, F.; Li, N.; Li, G.Q.; Luo, C.X. Occurrence of Fungicide Resistance in Botrytis cinerea from Greenhouse Tomato in Hubei Province, China. Plant Dis. 2016, 100, 2414–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Che, Z.; Chen, G. Multiple-fungicide resistance to carbendazim, diethofencarb, procymidone, and pyrimethanil in field isolates of Botrytis cinerea from tomato in Henan Province, China. Crop Prot. 2016, 84, 56–61. [Google Scholar] [CrossRef]
- Zhao, J.; Bi, Q.; Wu, J.; Lu, F.; Han, X.; Wang, W. Occurrence and management of fungicide resistance in Botrytis cinerea on tomato from greenhouses in Hebei, China. J. Phytopathol. 2019, 167, 413–421. [Google Scholar] [CrossRef]
- Korolev, N.; Mamiev, M.; Zahavi, T.; Elad, Y. Screening of Botrytis cinerea isolates from vineyards in Israel for resistance to fungicides. Eur. J. Plant Pathol. 2010, 129, 591–608. [Google Scholar] [CrossRef]
- Kretschmer, M.; Hahn, M. Fungicide resistance and genetic diversity of Botrytis cinerea isolates from a vineyard in Germany. J. Plant Dis. Prot. 2008, 115, 214–219. [Google Scholar] [CrossRef]
- Liang, X.; Peng, Y.; Zou, L.; Wang, M.; Yang, Y.; Zhang, Y. Baseline sensitivity of penthiopyrad against Colletotrichum gloeosporioides species complex and its efficacy for the control of Colletotrichum leaf disease in rubber tree. Eur. J. Plant Pathol. 2020, 158, 965–974. [Google Scholar] [CrossRef]
- Nottensteiner, M.; Absmeier, C.; Zellner, M. QoI Fungicide Resistance Mutations in Alternaria solani and Alternaria alternata are Fully Established in Potato Growing Areas in Bavaria and Dual Resistance against SDHI Fungicides is Upcoming. Gesunde Pflanz. 2019, 71, 155–164. [Google Scholar] [CrossRef]
- Malandrakis, A.A.; Apostolidou, Z.A.; Louka, D.; Markoglou, A.; Flouri, F. Biological and molecular charac-terization of field isolates of Alternaria alternata with single or double resistance to respiratory complex II and III inhibitors. Eur. J. Plant Pathol. 2018, 152, 199–211. [Google Scholar] [CrossRef]
- Thomas, A.; Langston, J.D.B.; Stevenson, K.L. Baseline Sensitivity and Cross-Resistance to Succinate-Dehydrogenase-Inhibiting and Demethylation-Inhibiting Fungicides in Didymella bryoniae. Plant Dis. 2012, 96, 979–984. [Google Scholar] [CrossRef] [Green Version]
- Angelini, R.M.D.M.; Habib, W.; Rotolo, C.; Pollastro, S.; Faretra, F. Selection, characterization and genetic analysis of laboratory mutants of Botryotinia fuckeliana (Botrytis cinerea) resistant to the fungicide boscalid. Eur. J. Plant Pathol. 2010, 128, 185–199. [Google Scholar] [CrossRef]
- Hägerhäll, C. Succinate: Quinone oxidoreductases: Variations on a conserved theme. Biochim. Biophys. Acta 1997, 1320, 107–141. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Cui, K.; Song, Y.; Li, T.; Liu, N.; Mu, W.; Liu, F. Activity of the Novel Succinate Dehydrogenase Inhibitor Fungicide Pydiflumetofen Against SDHI-Sensitive and SDHI-Resistant Isolates of Botrytis cinerea and Efficacy Against Gray Mold. Plant Dis. 2020, 104, 2168–2173. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.N.; Kim, Y.K.; Xiao, C.L. Molecular characterization of boscalid resistance in field isolates of Botrytis ci-nerea from apple. Phytopathology 2011, 101, 986–995. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.W.; Wu, Z.W.; Bi, C.W.; Wang, J.X.; Zhao, F.F.; Gao, J.; Hou, Y.P.; Zhou, M.G. Molecular and bio-chemical characterization of pydiflumetofen-resistant mutants of Didymella bryoniae. J. Agric. Food Chem. 2020, 68, 9120–9130. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Fu, L.; Tan, H.; Jiang, J.; Che, Z.; Tian, Y.; Chen, G. Resistance to Boscalid in Botrytis cinerea From Greenhouse-Grown Tomato. Plant Dis. 2021, 105, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; He, L.; Li, T.; Mu, W.; Liu, F. Development of Boscalid Resistance in Botrytis cinerea and an Efficient Strategy for Resistance Management. Plant Dis. 2021, 105, 1042–1047. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, Z.; Chen, L.; He, L.; Lu, H.; Ren, Y.; Mu, W.; Liu, F. Baseline Sensitivity of Botrytis cinerea to the Succinate Dehydrogenase Inhibitor Isopyrazam and Efficacy of this Fungicide. Plant Dis. 2016, 100, 1314–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.-P.; Mao, X.-W.; Wang, J.-X.; Zhan, S.-W.; Zhou, M.-G. Sensitivity of Fusarium asiaticum to a novel succinate dehydrogenase inhibitor fungicide pydiflumetofen. Crop Prot. 2017, 96, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Zhang, X.; Zhao, J.; Wang, W.; Shang, L.; Ma, S.; Adzavon, Y.M.; Lu, F.; Weng, M.; Han, X.; et al. Combination of Suspension Array and Mycelial Growth Assay for Detecting Multiple-Fungicide Resistance in Botrytis cinerea in Hebei Province in China. Plant Dis. 2019, 103, 1213–1219. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, J.; Lu, F.; Bi, Q.; Yang, K.; Han, X.; Wang, W. Baseline-sensitivity of Botrytis cinerea to pydiflumetofen and its efficacy on tomato gray mold in Hebei Province, China. Crop Prot. 2022, 158, 105989. [Google Scholar] [CrossRef]
- Wu, J.; Xue, Z.L.; Miao, J.Q.; Zhang, F.; Gao, X.; Liu, X.L. Sensitivity of different developmental stages and re-sistance risk assessment of Phytophthora capsici to fluopicolide in China. Front. Microbiol. 2020, 11, 185. [Google Scholar]
- Lin, D.; Xue, Z.L.; Miao, J.Q.; Huang, Z.Q.; Liu, X.L. Activity and resistance assessment of a new OSBP inhibitor, R034-1, in and the detection of point mutations in PcORP1 that confer resistance. J. Agric. Food Chem. 2020, 68, 13651–13660. [Google Scholar] [CrossRef]
- Li, S.; Hou, Y.; Peng, D.; Meng, L.; Wang, J.; Zhou, M.; Chen, C. Baseline sensitivity and control efficacy of flutolanil in Rhizoctonia solani. Australas. Plant Pathol. 2014, 43, 313–320. [Google Scholar] [CrossRef]
- Zhang, C.Q.; Yuan, S.K.; Sun, H.Y.; Qi, Z.Q.; Zhou, M.G.; Zhu, G.N. Sensitivity of Botrytis cinerea from veg-etable greenhouses to boscalid. Plant Pathol. 2007, 56, 646–653. [Google Scholar] [CrossRef]
- Allen, P.J. Metabolic Aspects of Spore Germination in Fungi. Annu. Rev. Phytopathol. 1965, 3, 313–342. [Google Scholar] [CrossRef]
- Fan, F.; Hamada, M.S.; Li, N.; Li, G.Q.; Luo, C.X. Multiple Fungicide Resistance in Botrytis cinerea from Greenhouse Strawberries in Hubei Province, China. Plant Dis. 2017, 101, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Ortuno, D.; Perez-Garcia, A.; Chamorro, M.; Eduardo, D.L.P.; De Vicente, A.; Tores, J.A. Re-sistance to the SDHI fungicides boscalid, fluopyram, fluxapyroxad, and penthiopyrad in Botrytis cinerea from commercial strawberry fields in Spain. Plant Dis. 2017, 101, 1306–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroux, P.; Gredt, M.; Leroch, M.; Walker, A.S. Exploring mechanisms of resistance to respiratory inhibitors in field strains of Botrytis cinerea, the causal agent of gray mold. Appl. Environ. Microbiol. 2010, 76, 6615–6630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallik, I.; Arabiat, S.; Pasche, J.S.; Bolton, M.D.; Patel, J.S.; Gudmestad, N.C. Molecular Characterization and Detection of Mutations Associated with Resistance to Succinate Dehydrogenase-Inhibiting Fungicides in Alternaria solani. Phytopathology 2014, 104, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Outwater, C.A.; Proffer, T.J.; Rothwell, N.L.; Peng, J.; Sundin, G.W. Boscalid resistance in Blumeriella jaapii: Distribution, effect on field efficacy, and molecular characterization. Plant Dis. 2019, 103, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Amiri, A.; Zuniga, A.I.; Peres, N.A. Mutations in the membrane-anchored SdhC subunit affect fitness and sen-sitivity to succinate dehydrogenase inhibitors in Botrytis cinerea populations from multiple hosts. Phytopathology 2020, 110, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Veloukas, T.; Markoglou, A.N.; Karaoglanidis, G.S. Differential Effect of SdhB Gene Mutations on the Sensitivity to SDHI Fungicides in Botrytis cinerea. Plant Dis. 2013, 97, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Amiri, A.; Heath, S.M.; Peres, N.A. Resistance to fluopyram, fluxapyroxad, and penthiopyrad in Botrytis ci-nerea from strawberry. Plant Dis. 2014, 98, 532–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, R.W.; Entrop, A.-P.; Goertz, A.; Mehl, A. Status of Sensitivity of Northern German Botrytis Populations to the New Sdhi Fungicide Fluopyram Prior to its Release as a Commercial Fungicide. J. Plant Dis. Prot. 2015, 122, 81–90. [Google Scholar] [CrossRef]
- Veloukas, T.; Karaoglanidis, G.S. Biological activity of the succinate dehydrogenase inhibitor fluopyram against Botrytis cinerea and fungal baseline sensitivity. Pest Manag. Sci. 2011, 68, 858–864. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taget Gene | Primers | Sequence (5′→3′) | Amplicon Size (bp) | Annealing Temperature/°C |
|---|---|---|---|---|
| SdhA | SdhA-F SdhA-R | CTTCGCAATTGAAGATTTGGAAACGC ATCCATGCCCCTTTCGTGTCT | 2609 | 56 |
| SdhB | SdhB-F SdhB-R | ACCTACTCGCCCTATCCAAT AGACTTAGCAATAACCGCCC | 1004 | 54 |
| SdhC | SdhC-F SdhC-R | GCCAGATTTCCTTAGTCAG GCTGGACTCTGAATGTGAT | 1065 | 52 |
| SdhD | SdhD-F SdhD-R | ATGGCTTCATTCATCAAACCATC TTATGCGCGCCAAATTCTTTTG | 650 | 56 |
| Region | Number of Isolates | EC50 (mg/L) a | |||
|---|---|---|---|---|---|
| Mycelial Growth | Conidia Germination | ||||
| Range | Mean ± SD b | Range | Mean ± SD b | ||
| Handan | 21 | 0.090~2.456 | 1.098 ± 0.581 a | 0.055~0.154 | 0.098 ± 0.028 a |
| Xingtai | 18 | 0.301~2.080 | 0.944 ± 0.480 a | 0.051~0.128 | 0.101 ± 0.035 a |
| Shijiazhuang | 20 | 0.128~1.892 | 1.024 ± 0.595 a | 0.053~0.168 | 0.107 ± 0.036 a |
| Baoding | 26 | 0.069~2.550 | 1.128 ± 0.835 a | 0.034~0.142 | 0.107 ± 0.034 a |
| Cangzhou | 16 | 0.080~2.536 | 0.893 ± 0.543 a | 0.045~0.128 | 0.090 ± 0.025 a |
| Hengshui | 13 | 0.039~2.088 | 1.000 ± 0.506 a | 0.013~0.186 | 0.098 ± 0.047 a |
| Qinhuangdao | 17 | 0.071~2.134 | 1.176 ± 0.758 a | 0.039~0.286 | 0.098 ± 0.053 a |
| Total | 131 | 0.039~2.550 | 1.054 ± 0.633 | 0.013~0.286 | 0.101 ± 0.037 |
| Region | Number of Isolates | Isolate/Percentage of the Total Population/% | |||
|---|---|---|---|---|---|
| S | LR | MR | HR | ||
| Dingxing, Baoding | 32 | 24 (75.0) | 7 (21.9) | 1 (3.1) | 0 (0.0) |
| Xushui, Baoding | 18 | 14 (77.8) | 4 (22.2) | 0 (0.0) | 0 (0.0) |
| Weichang, chengde | 18 | 10 (55.6) | 7 (38.9) | 1 (5.6) | 0 (0.0) |
| Feixiang, Handan | 24 | 0 (0.0) | 4 (16.7) | 12 (50) | 8 (33.3) |
| Raoyang, Hengshui | 15 | 15 (100.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Gaoyi, Shijiazhuang | 19 | 2 (10.5) | 6 (31.6) | 4 (21.1) | 7 (36.8) |
| Kaiping, Tangshan | 27 | 0 (0.0) | 4 (14.8) | 6 (22.2) | 17 (63.0) |
| Guangzong, Xingtai | 34 | 0 (0.0) | 9 (26.5) | 13 (38.2) | 12 (35.3) |
| Total | 187 | 65 (34.8) | 39 (20.9) | 38 (20.3) | 45 (24.1) |
| Name of Isolates c | Phenotype | Mutation Type | |||
|---|---|---|---|---|---|
| sdhA | sdhB | sdhC | sdhD | ||
| 21BDDM6, 21CDCYW16 | S | - | - | - | - |
| 21CDLQ18 | S | - | - | G85A, I93V, M158V, V168I | - |
| 21CDCYW13, 21TS8, 21TS15, 21TS17, 21XTG17, 21XTG19 | LR | - | N230I | - | - |
| 21HDFH4, 21HDFH6, 21HDFH7, 21HDFH8, 21HDFH9, 21HDFH11, 21HDFH12, 21TS6 | MR | - | N230I | - | - |
| 21HDFH5, 21HDFH10, 21HDFH13, 21TS7, 21TS9, 21TS10, 21TS11, 21TS12, 21TS13, 21TS14, 21TS16, 21TS18, 21TS19, 21TS20 | HR | - | N230I | - | - |
| 21BDKG9, 21BDHE1 | LR | - | H272R | - | - |
| 21HDH1 | HR | - | H272R | - | - |
| 21BDL10, 21XTG29, 21XTG28 | LR | - | P225F | - | - |
| 21HDFH14, 21BDXS1, 21HDFH18 | LR | - | P225L | - | - |
| Isolates | Mutation Types | EC50/(mg/L) e | |||
|---|---|---|---|---|---|
| Penthiopyrad | Fluopyram | Pydiflumetofen | Boscalid | ||
| 21BDDM6 | - d | 2.874 ± 1.217 e | 1.140 ± 0.083 h | 1.482 ± 0.463 ef | 6.091 ± 1.715 c |
| 21CDCYW16 | - | 1.582 ± 0.301 e | 0.311 ± 0.025 h | 0.491 ± 0.052 f | 3.739 ± 2.182 c |
| 21CDLQ18 | - | 0.210 ± 0.090 e | 0.603 ± 0.108 h | 0.099 ± 0.034 f | 1.320 ± 0.188 c |
| 21BDHE1 | H272R | 41.82 ± 6.31 de | 79.15 ± 7.49 de | 3.999 ± 0.153 def | 938.26 ± 475.05 a |
| 21HDH1 | H272R | 383.06 ± 43.19 b | 152.45 ± 30.01 c | 39.76 ± 4.75 a | >500 z |
| 21BDKG9 | H272R | 15.00 ± 3.65 de | 42.58 ± 8.54 efg | 4.174 ± 0.250 cdef | >500 z |
| 21BDL10 | P225F | 26.30 ± 2.97 de | 79.70 ± 2.55 de | 2.604 ± 0.579 def | 255.99 ± 45.97 bc |
| 21XTG28 | P225F | 37.99 ± 5.67 de | 351.00 ± 39.30 a | 5.064 ± 0.513 cdef | >500 z |
| 21XTG29 | P225F | 43.92 ± 3.15 de | 308.99 ± 38.70 b | 13.19 ± 2.19 bc | >500 z |
| 21HDFH14 | P225L | 30.41 ± 7.46 de | 64.72 ± 16.13 def | 33.49 ± 7.11 a | 329.81 ± 87.59 bc |
| 21HDFH18 | P225L | 38.88 ± 11.07 de | 90.16 ± 9.40 d | 5.96 ± 0.56 cdef | >500 z |
| 21BDXS | P225L | 35.69 ± 5.60 de | 20.11 ± 2.22 gh | 9.838 ± 1.072 bcde | 356.82 ± 33.32 b |
| 21TS12 | N230I | 488.31 ± 63.38 a | 69.48 ± 5.82 def | 18.65 ± 1.34 b | 124.16 ± 5.31 bc |
| 21TS16 | N230I | 164.83 ± 14.57 c | 41.17 ± 10.68 fg | 10.93 ± 4.92 bcd | 70.50 ± 2.69 bc |
| 21TS6 | N230I | 54.21 ± 12.85 d | 41.46 ± 0.46 fg | 2.625 ± 0.046 def | 807.95 ± 66.67 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, Q.; Lu, F.; Yang, K.; Wu, J.; Zhang, S.; Han, X.; Wang, W.; Zhao, J. Baseline Sensitivity and Resistance of Botrytis cinerea to Penthiopyrad in Hebei Province, China. Horticulturae 2022, 8, 686. https://doi.org/10.3390/horticulturae8080686
Bi Q, Lu F, Yang K, Wu J, Zhang S, Han X, Wang W, Zhao J. Baseline Sensitivity and Resistance of Botrytis cinerea to Penthiopyrad in Hebei Province, China. Horticulturae. 2022; 8(8):686. https://doi.org/10.3390/horticulturae8080686
Chicago/Turabian StyleBi, Qiuyan, Fen Lu, Kexin Yang, Jie Wu, Shangqing Zhang, Xiuying Han, Wenqiao Wang, and Jianjiang Zhao. 2022. "Baseline Sensitivity and Resistance of Botrytis cinerea to Penthiopyrad in Hebei Province, China" Horticulturae 8, no. 8: 686. https://doi.org/10.3390/horticulturae8080686
APA StyleBi, Q., Lu, F., Yang, K., Wu, J., Zhang, S., Han, X., Wang, W., & Zhao, J. (2022). Baseline Sensitivity and Resistance of Botrytis cinerea to Penthiopyrad in Hebei Province, China. Horticulturae, 8(8), 686. https://doi.org/10.3390/horticulturae8080686