Abstract
Hibiscus syriacus is a highly ornamental flowering shrub widely grown in East Asia. Its abundant flower colors mainly reflect the accumulation of anthocyanins. Classifying H. syriacus petals and identifying the relationship between flower color and anthocyanins can provide references for flower color breeding. With eight cultivars of H. syriacus as experimental materials, the floral color was described using the Royal Horticultural Society Color Chart and the CIEL*a*b* coordinate. The anthocyanin in petals was qualitatively and quantitatively analyzed by using ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS), and the relationships between flower color and anthocyanin were analyzed by using stepwise regression analysis. We divided eight cultivars of H. syriacus into five color lines: red-purple, white, violet, purple, and blue-purple, and then into two groups: purple and red-purple. The results showed that the L* had a significant negative correlation with a* and C*. A total of 52 anthocyanin components were detected in the petals of H. syriacus, including cyanidin, delphinidin, malvidin, petunidin, peonidin, and pelargonidin derivatives. Cyanidin derivatives accounted for the highest proportion, and pelargonidin derivatives accounted for the lowest proportion of the total anthocyanins. Petals of ‘Pink Giant’ contained the largest number of types of anthocyanin components and the highest total anthocyanin content, while the petals of ‘Albus Single’ contained the smallest. Petunidin-3-O-glucoside showed a significant negative correlation with L* in the red-purple group and in all cultivars, whereas it showed a significant negative correlation with b* in the purple group. Delphinidin-3-O-(6-O-Malonyl-β-D-glucoside) exhibited a significant positive correlation with a* in the red-purple group, and delphinidin-3-O-glucoside showed a significant negative correlation with L* in the purple group. Therefore, our results suggest that changing the content of these three anthocyanin components may have the potential to alter the flower color. This research provides scientific guidance and a foundation for the molecular breeding of H. syriacus cultivars with new floral colors.
1. Introduction
Flower color is one of the most important ornamental traits of flowers, and it is also one of the key goals for the flower breeding. According to previous research, a wide variety of flower pigments could be generally divided into three groups, carotenoids, flavonoids, and alkaloids [1]. Carotenes and lutein in carotenoids provide petals and fruits with colors ranging from yellow to orange [2,3], while lycopene provides a red color [4]. Betalains cause the petals to be red-purple and yellow due to two different substances: betacyanin and betaxanthin [5]. However, although anthocyanins, chalcones, and some flavanols in flavonoids can play a role in flower pigments, anthocyanins are the most important group of water-soluble flavonoids because they produce a wide range of colors [1,6]. Currently, more than 700 anthocyanin components have been reported, which are the main substances affecting flower color [7].
Anthocyanins are generally identified and quantified by ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS) analysis, which is a labor-intensive and expensive method of analysis. It is cheaper and easier to estimate anthocyanin content by color space values measured by a colorimeter compared to using UPLC-MS. Moreover, color space values of many species, such as grape, strawberry, and chrysanthemum, were closely correlated to anthocyanin content [8,9,10]. Therefore, according to the relationship between anthocyanins and flower color, the anthocyanin content of H. syriacus can be estimated by measuring the color space value.
Currently, the CIEL*a*b* (Commission Internationale de l’Eclairage) coordinate is a very practical color space that quantifies luminance, green-red components, and blue-yellow components by using L*, a*, and b*, respectively, as geometric coordinates to characterize specific colors. It has been widely used to determine and classify floral and fruit color phenotypes, including peony, blueberry, chrysanthemum, cassava, calla, etc. [10,11,12,13,14]. Therefore, exploring the correlation between anthocyanins and flower color space values, including L*, a*, b*, C*, as well as h° by linear regression analysis is necessary.
Hibiscus syriacus L, belonging to the Malvaceae family, is a flowering shrub of high ornamental value, which is widely cultivated in East Asia, including countries such as China and South Korea. Its blooming period generally lasts for five months, from late May to early October. Its delicate and attractive flowers have a wide range of colors, shapes, and sizes. H. syriacus has high medicinal and industrial value because of its biological functions, which can help people resist disease, and it is also rich in nutrients [15,16]. In addition, the extraction method of H. syriacus anthocyanins and the physicochemical properties and biological functions of anthocyanin extraction have also been reported [17,18]. At present, many different cultivars have been obtained by different breeding methods. For example, the new varieties ‘Daewangchun’ and ‘Red Bohanjae’ were developed by interspecific hybridization [19,20], and a new variety, ‘Dasom,’ was created by radiation breeding [21]. H. syriacus can be classified by two flower phenotypes: petal morphology and colors. The molecular mechanism of H. syriacus anthocyanin accumulation was preliminarily explained by transcriptome [22]. The anthocyanin component and content in H. syriacus has been reported [23]. However, this study included only a few cultivars, which could not reflect the color of most H. syriacus cultivars. The relationship between anthocyanins and color space values, which is conducive to the directional breeding of flower color, has not been discussed. Therefore, exploring the mechanism of H. syriacus petal color formation will be helpful for cultivating new cultivars with different floral colors and higher commercial values.
In this study, cultivars of H. syriacus with different colors were used as the materials, the petal colors were described by Royal Horticultural Society Color Chart (RHSCC) and a colorimeter, and the anthocyanins in the petals were identified and quantified by UPLC-MS. The relationship between anthocyanin and color was analyzed to provide more information for revealing the mechanism of color formation and the breeding of H. syriacus.
2. Materials and Method
2.1. Plant Materials
Eight cultivars of H. syriacus, exhibiting different colors, were selected for this experiment, including ‘Pink Giant,’ ‘Albus Single,’ ‘Blueberry Smoothie,’ ‘Pink Chiffon,’ ‘Bridesmaid,’ ‘Blue Chiffon,’ ‘Magenta Chiffon,’ and ‘Raspberry Smoothie.’ Among them, ‘Pink Giant’ was bred in France; ‘Pink Chiffon,’ ‘Magenta Chiffon,’ and ‘Blue Chiffon’ were bred by Roderick Woods, a famous British hibiscus breeder; ‘Blueberry Smoothie’ and ‘Raspberry Smoothie’ are American-bred cultivars; and ‘Bridesmaid’ and ‘Albus Single’ are native cultivars in China. These cultivars of H. syriacus were planted in March 2019 in the experimental station of Wenzhou Vocational College of Science and Technology. The pH of the soil is 7.27. In 2021, The ‘Magenta Chiffon’ first bloomed on 10 June, followed by the ‘Pink Chiffon’ and ‘Raspberry Smoothie’ on 12 June, ‘Pink Giant’ on 15 June, the ‘Bridesmaid’ on 18 June, the ‘Blue Chiffon’ on 19 June, the ‘Albus Single’ on 10 July, and ‘Blueberry Smoothie,’ the latest to bloom, on 12 July. The colors of these cultivars include white, blue-purple, pink, pink-purple, light blue, magenta, and deep red (Figure 1), which basically cover all the flower colors of the H. syriacus. These materials were grown at Wenzhou Vocational College of Science and Technology, China (120°7′, 27°99′), under natural temperatures and light conditions. On 7 September 2021, at 8–10 a.m., five fully open flowers of each cultivar were sampled for color classification, and 5.0 g of petal from each cultivar were sampled into test tubes and stored in a –80 °C refrigerator for later use, after quick freezing in liquid nitrogen.

Figure 1.
Flower colors of eight cultivars of Hibiscus syriacus. (A) ‘Albus Single’ (AS); (B) ‘Pink Chiffon’ (PC); (C) ‘Blue Chiffon’ (BC); (D) ‘Blueberry Smoothie’ (BS); (E) ‘Bridesmaid’ (BM); (F) ‘Raspberry Smoothie’ (RS); (G) ‘Magenta Chiffon’ (MC); (H) ‘Pink Giant’ (PG).
2.2. Colorimetric Measurement and Classification
The color comparison of H. syriacus petals was carried out using the RHSCC. The color space values of eight cultivars were measured with a colorimeter (CR-10 plus, Konica Minolta, JPN) under the condition of a C/2 light source. Five flowers were taken from each cultivar, and one petal was taken from each flower as a sample. The petals were placed on clean white paper, and the colorimeter was used to measure each sample at the middle of the petals and not touch the spot [24]. The average values of L*, a*, and b* were obtained. The lightness and darkness of the color, ranging from white to black, are represented by L*; a* and b* represent different color directions in the color system: a* ranges from red to green, b* ranges from yellow to blue. C* and h° were derived from a* and b*, using the following equations:
C* = [(a*)2 + (b*)2]1/2
h° = tan−1 (b*/a*)
2.3. Extraction and Analysis of Anthocyanins
UPLC-MS was used to analyze the composition and content of the anthocyanins in the petals of H. syriacus with different flower colors. Each petal sample was ground to a powder with a ball mill (30 Hz, 1.5 min), and 50 mg of the powder was weighed and dissolved in 500 μL of extraction solution (50% methanol in water, containing 0.1% hydrochloric acid); the mixture was then vortexed for 5 min, sonicated for 5 min, centrifuged for 3 min (12,000 r/min, 4 °C), the supernatant was aspirated, and the operation was repeated once. The two supernatants were combined, and the samples were filtered with a microporous membrane (0.22 μm pore size) and stored in a sample vial for UPLC-MS analysis. The MWDB (Metware Database) was constructed based on the standard products, and the data detected by mass spectrometry was qualitatively analyzed. Quantification was performed using the multiple reaction monitoring (MRM) analysis of triple quadrupole mass spectrometry. The liquid phase conditions mainly included the following: the chromatographic column was ACQUITY BEH C18 1.7 µm, 2.1 mm × 100 mm; the mobile phase was ultrapure water (add 0.1% formic acid) for phase A, and methanol (add 0.1% formic acid) for phase B; the elution gradient was 0 min with a phase B ratio of 5%, increased to 50% at 6 min, increased to 95% at 12 min, maintained for 2 min, decreased to 5% at 14 min, and equilibrated for 2 min; the flow rate was 0.35 mL/min; the column temperature was 40 °C; and the injection volume was 2μL. The mass spectrometry conditions mainly included: electrospray ionization (ESI) temperature of 550 °C, mass spectrometry voltage of 5500 V in positive ion mode, and curtain gas (CUR) of 35 psi. In Q-Trap 6500+, each ion pair is scanned according to the optimized declustering potential (DP) and collision energy (CE).
2.4. Correlation between Color Space Values and Anthocyanins
Linear regression analysis was used to investigate the relationships between flower color space values, including L*, a*, b*, C*, and h°. Statistical analyses were performed with Originpro 8.0 and SPSS 26.0.
3. Result
3.1. Colorimetric Evaluation and Classification
The eight cultivars of H. syriacus in the present study (Figure 1) were roughly divided into five major color lines according to the comparison results of the RSHCC (Table 1): (1) the red-purple line, including ‘Pink Chiffon,’ ‘Pink Giant,’ ‘Magenta Chiffon,’ and ‘Raspberry Smoothie;’ (2) the white line, including ‘Albus Single;’ (3) the violet line, with ‘Blueberry Smoothie;’ (4) the purple line, with ‘Bridesmaids;’ and (5) the purplish blue line, with ‘Blue Chiffon.’ To better explore the relationship between the color space value and anthocyanin in different color lines of H. syriacus, we divided the white, violet, purple, and blue-purple lines into one group, called the purple group, and the red-purple lines into another, called the red-purple group.

Table 1.
Cultivars of Hibiscus syriacus used in this study, along with their chromatic parameters.
A colorimeter was used to determine the color space values (L*, a*, and b*) of eight cultivars of H. syriacus. The results showed that the L* values of the petals in different color lines decreased gradually, with the color becoming darker (Table 1). Among the petals of the five flower color lines, red-purple, violet, and purple were mainly concentrated in quadrant IV. The white line ‘Albus Single’ had the highest L* and the lowest a*, while the red-purple line ‘Pink Giant’ had the lowest L* and the highest a*. Furthermore, the white line ‘Albus Single’ had the highest b*, while the red-purple line ‘Raspberry Smoothie’ had the lowest (Figure 2). To elucidate the distribution of these color space values, correlation analysis was performed on L*, a*, and b*, which showed that L* was negatively correlated with a* (R2 = 0.776, p = 0.04) in all color groups; there was a weak positive correlation between L* and b* (R2 = 0.525, p = 0.04), and a strong negative correlation between L* and c* (R2 = 0.799, p < 0.01); we found that a* and c* both increased with the decrease of L*. (Figure 3). In addition, the correlations among L*, a*, b*, and c* were different among different color groups of H. syriacus. In the red-purple group, there was a strong negative correlation between L* and c* (R2 = 0.812, p < 0.01), and in the purple group, there was a strong positive correlation between L* and a* (R2 = 0.803, p < 0.01) and L* and b* (R2 = 0.976, p < 0.01) (Figure 4).
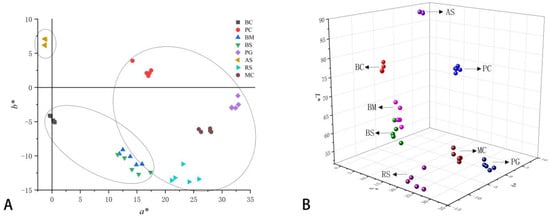
Figure 2.
Flower color distribution of Hibiscus syriacus in two-dimensional rectangular (a*, and b*) (A), and three-dimensional (L*, a* and b*) coordinates (B).
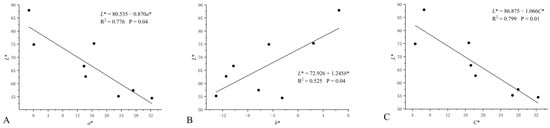
Figure 3.
The linear regression between L* and a* (A), b* (B), C* (C) of eight cultivars of Hibiscus syriacus. Each solid circle in each panel was the mean value of five replicates of each cultivar of Hibiscus syriacus.
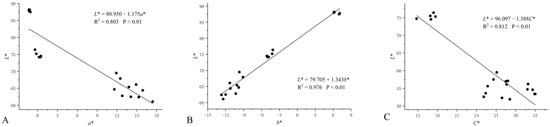
Figure 4.
The linear regression between L* and a* (A), L* and b* (B) in the purple group of Hibiscus syriacus (B), and L* and C* (C) in the red-purple group of Hibiscus syriacus. Each solid circle in each panel was the mean value of five replicates of each cultivar of Hibiscus syriacus.
3.2. Identification and Quantification of Anthocyanins
The TIC diagram of a QC sample is shown below (Figure 5A). The results of the multi-peak detection in the MRM mode revealed the metabolites of the sample (Figure 5C). Moreover, we superimposed the TIC diagram of three QC samples, and the results showed that the metabolites were highly overlapped (Figure 5B), indicating that the detection technology had good repeatability. Based on the local metabolite database, qualitative and quantitative mass spectrometry analysis of metabolites in the samples was performed. In the picture (Figure 5C), each peak of a different color represents the detected metabolite. Information about metabolite serial numbers, peak integration values, and metabolite names is listed in Table 1. In total, 52 anthocyanins were identified, among which were cyanidin, delphinidin, malvidin, petunidin, peonidin, and pelargonidin, and ‘Pink Giant’ contained the greatest number of species of the anthocyanin component (47), and ‘Albus Single’ contained the lowest number, with only 20 (Figure 6A). As for the total anthocyanin content, it was found that ‘Albus Single’ contained the smallest amount at 3.13 μg g−1, and ‘Pink Giant’ contained the highest amount at 2713.21 μg g−1 (Figure 6B). The highest content identified among the different types of anthocyanins was for cyanidin, the lowest was for pelargonidin. We found that the color of the flowers changed from white to pink to blue to purple to red-purple as the total anthocyanin content increased.
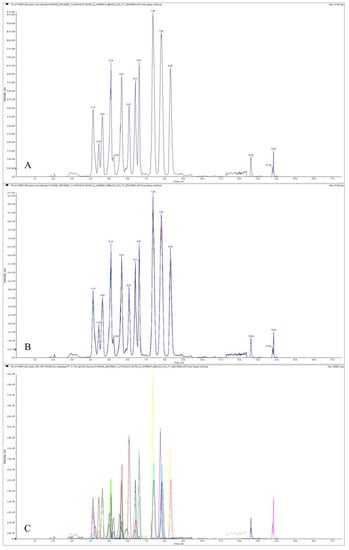
Figure 5.
Total ion current diagram for quality control sample by mass spectrometry (A), and total ion current overlaps for quality control sample by mass spectrometry (B); multi-peak detection plot of metabolites by MRM metabolite (C).
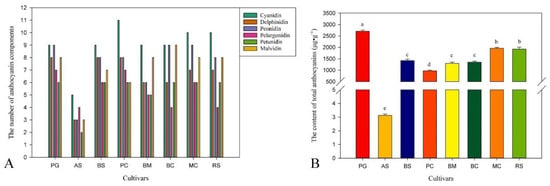
Figure 6.
The number of anthocyanin components (A), and the contents of total anthocyanins in petals of Hibiscus syriacus (B). In (B) different letters indicate significant differences at p ≤ 0.05.
3.3. Correlation between Color Space Values and Anthocyanins
After horizontal normalization of the relative anthocyanin content of each metabolite, the H. syriacus cultivars and metabolites were divided into two clear categories. Accordingly, we found that ‘Raspberry Smoothie’ and ‘Magenta Chiffon’ made up one group, and ‘Blueberry Smoothie’ and ‘Bridesmaid’ constituted another group, indicating that the relative content of anthocyanins between their groups was relatively close, so the flower color phenotype is roughly the same. We also found that petunidin and delphinidin were clustered together, and pelargonidin and malvidin were clustered together. According to the heat map clustering of anthocyanin component, ‘Albus Single’ and ‘Blue Chiffon’ were in one group, and ‘Bridesmaids’ and ‘Magenta Chiffon’ were in another group (Figure 7B), which indicates that the relative content of the anthocyanin component of the two cultivars within this group is closer. In order to explore the relationship between anthocyanin content in H. syriacus petals and their flower color, a stepwise multiple linear regression analysis was conducted by using color space values and anthocyanins. The L *, a *, and b * values, which were variables describing the flower colors, were taken as dependent variables, and the contents of total anthocyanin (X1), the main anthocyanin, as well as that of cyanidin (X2), delphinidin (X3), peonidin (X4), pelargonidin (X5), petunidin (X6), and malvidin (X7), were taken as independent variables to establish the equations and deduce the quantitative relationships. It can be seen from the R2, r, and P values that there was a significant correlation between flower color and anthocyanin content (Table 2). According to the regression equation of H. syriacus, it was determined that X6 was significantly and positively correlated with L*. In the red-purple group, L* was significantly negatively correlated with X6, while a* was significantly positively correlated with X3. In the purple group, L* was significantly negatively correlated with X3, and both X6 and X7 were significantly correlated with b*; the magnitude of the absolute value of the regression coefficient can directly determine the contribution of anthocyanins to flower color, while the regression coefficient of X6 (0.107) was greater than that of X7 (0.006), indicating that the negative correlation effect of X6 on b* was significant. Therefore, the anthocyanins that determine the space value of color were delphinidin and petunidin, and malvidin can also influence flower color, but the effect was slight.
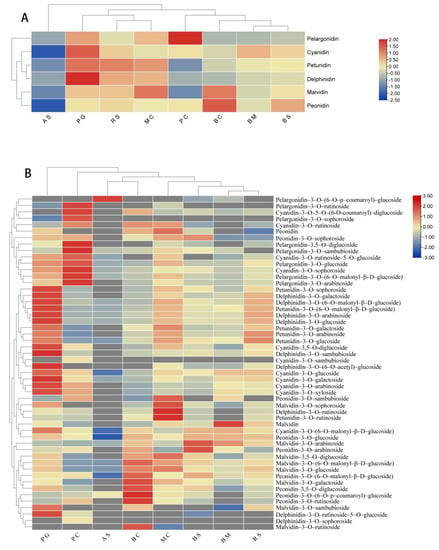
Figure 7.
Heat map of relative anthocyanin contents of different Hibiscus syriacus (A), clustering heat map of relative anthocyanin component contents of different Hibiscus syriacus (B).
Table 2.
Stepwise regression equation of color space value with anthocyanin of Hibiscus syriacus.
In order to further explore the relationship between the anthocyanin derivatives in petals and flower color, we performed a stepwise regression for the anthocyanin derivatives and L*, a*, and b* (Table 3). From the H. syriacus regression equation, L* was significantly negatively correlated with petunidin-3-O-glucoside. In the red-purple group, L* was significantly negatively correlated with petunidin-3-O-glucoside, and a* was significantly positively correlated with delphinidin-3-O-(6-O-malonyl-β-D-glucoside).
Table 3.
Stepwise multiple regression equations of anthocyanin with L*, a*, and b*.
In the purple group, the type with a significantly negative correlation on L* was delphinidin-3-O-glucoside, and b* was significantly negatively correlated with petunidin-3-O-glucoside.
4. Discussion
H. syriacus is a traditional ornamental, edible, and medicinal flowering shrub with high economic value [25,26]. We found that the color lines of the H. syriacus flowers include white, purple, blue-purple, violet, and red-purple line, but lack a red, blue, yellow, and green line. Generally, the yellow line contains a large amount of carotenoids in the petals [27], while the green line has higher chlorophyll levels [28]. Therefore, the flower colors reveal the fact that the petals of these H. syriacus cultivars contain low amounts of carotenoids and chlorophyll, but this fact still needs to be proved by further experiments. Therefore, we determined to color-relate anthocyanins to the petal color to provide a reference and a direction for H. syriacus flower color breeding.
Based on the results, the flower color was not only related to the type of anthocyanins, but also to the content of anthocyanins. The anthocyanin contained in the aglycone structure petals of H. syriacus included: cyanidin, delphinidin, malvidin, petunidin, peonidin, and pelargonidin. Generally, pelargonidin and cyanidin cause petals to produce red and purple colors, respectively, while delphinidin produce blue and purple colors [29]. In the present study, except for ‘Albus Single’ and ‘Pink Chiffon,’ the remaining six cultivars contained high amounts of delphinidin, causing the petals to emerge as purple. Although a* was positively correlated with delphinidin in H. syriacus, the red-purple group was compared with the purple group, and the former showed red color precisely because of the large amount of pelargonidin in the petals. ‘Pink Giant’ has a deep purple-red color because it contains the highest levels of cyanidin and higher levels of pelargonidin. If the type and content of anthocyanins are similar, the color of the flowers will be closer, for example, ‘Magenta Chiffon’ and ‘Raspberry Smoothie’ have a similar number of component and a similar total content of anthocyanins, so that they have similar colors, and they were grouped together in the anthocyanin component clustering heat map (Figure 7A). Meanwhile, in the cluster heat map, petunidin and delphinidin were grouped together, and pelargonidin and malvidin were grouped together. This shows that the functions of the substances in these two groups were identical and will have the same effect on a certain direction of the color space (Figure 7A). The petals of white flowers also contain few anthocyanins, including examples such as white azalea, honeysuckle, and rose [30,31,32], and nearly all white flowers contain some flavanols and flavonoids, such as kaempferol, quercetin, apigenin, and luteolin, so almost all white flowers are “light yellow” [33]. We detected a small amount of anthocyanins in ‘Albus Single,’ which has the highest b* level among all the cultivars. We also discovered that the higher content of anthocyanins in the petals influenced the darker flower color, which is consistent with the previous studies on azaleas and roses [30,32].
There were significant differences in the effects of the type and quantity of the anthocyanin component on the flower color of different color cultivars. Previously, a total of 40 anthocyanins were reported in four H. syriacus cultivars. In this study, total of 52 anthocyanin components were qualitatively and quantitatively determined. According to the stepwise multiple regression equation, it can be determined that there was a linear relationship between anthocyanins in H. syriacus petals and their flower color, and petunidin-3-O-glucoside had a significant negative correlation with the brightness of the petals. In the purple group, however, it is delphinidin-3-O-glucoside which has a significantly negative correlation with petal brightness. Delphinidin-3-O-(6-O-malonyl-β-D-glucoside) positively regulated the redness of H. syriacus petals significantly in the red-purple group. At the same time, petunidin-3-O-glucoside had a significantly negative regulatory effect on the blueness of petals in the purple group. We also found that delphinidin-3-O-glucoside and delphinidin-3-O-arabinoside made up a group, and petunidin-3-O-glucoside and petunidin-3-O-arabinoside constituted a group, according to the clustering heat map of the anthocyanin components. The content between the two groups has a similar multiple relationship in each cultivar, and perhaps they can jointly control the brightness or blueness through some relationship. We also used linear regression to analyze the correlation between flower color and anthocyanins, and calculated the anthocyanin content of H. syriacus rapidly and accurately through the spatial value of flower color.
A comparative transcriptome analysis of the two cultivars of H. syriacus [34] indicated that the differential genes between ‘Albus Single’ and ‘Pink Giant’ may be C4H, CHS, CHI and DFR. The upregulation of the expression of the above structural genes leads to the accumulation of anthocyanins. In the study of cyclamen, the F3′5′H gene was suppressed, resulting in a decrease in delphinidin and an increase in cyanidin, leading to a change in petal color from purple to red and finally, to pink [35]. Therefore, we can increase or decrease other flower anthocyanins in the petals by molecular breeding to make it express different colors. The path for the flower color directed breeding was indicated, thus expanding the germplasm resources of H. syriacus.
5. Conclusions
In this study, we divided H. syriacus into five color lines, and then into two groups. L* had a comparatively significant negative correlation with a* and C*. The aglycon skeletons of anthocyanins included cyanidin, delphinidin, malvidin, petunidin, peonidin, and pelargonidin. A total of 52 anthocyanin components were detected from H. syriacus. The types and contents of most anthocyanin components were greater in the red-purple group than in the purple group. Petunidin-3-O-glucoside, delphinidin-3-O-(6-O-malonyl-β-D-glucoside), and delphinidin-3-O-glucoside were correlated to petal brightness, redness, and blueness, respectively. Flower color was related to the type and content of anthocyanin components. Finally, we can quickly and accurately estimate anthocyanin content using linear equations based on color space value through the relationship between flower color and anthocyanin.
At present, there are only white, purple, and red-purple H. syriacus on the market, and other color cultivars have not yet been cultivated. We can increase or reduce the anthocyanin in H. syriacus petals through molecular breeding, or change the flower color other ways, even creating a new flower color, so as to cultivate new cultivars to satisfy the various demands of the market. Therefore, it is particularly important to explore the relationship between anthocyanins and flower color, thereby providing a reference for the directional breeding of H. syriacus flower color.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae9010048/s1, Table S1: The composition and content of anthocyanin in Hibiscus syriacus flowers.
Author Contributions
Resources, J.C.; methodology, J.C. and L.Z.; analysis, J.C. and H.Y.; writing, J.C., H.Y., J.W. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Wenzhou Featured and Prevalent Tree and Flower New Variety Breeding Cooperation Group Project: Germplasm Resources Collection and New Variety Breeding of Hibiscus syriacus (2019ZX004-4) and the Personnel Startup Project of the Scientific Research and Development Foundation of Zhejiang A&F University (2021FR041).
Data Availability Statement
All datasets obtained for this study are included in the manuscript/Supplementary Materials.
Acknowledgments
We thank Yiguang Wang for the revision of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Ohmiya, A. Qualitative and quantitative control of carotenoid accumulation in flower petals. Sci. Hortic. Amsterdam. 2013, 163, 10–19. [Google Scholar] [CrossRef]
- Clevidence, B.; Paetau, I.; Smith, J.C. Bioavailability of carotenoids from vegetables. HortScience 2000, 35, 585–588. [Google Scholar] [CrossRef]
- Kong, K.W.; Khoo, H.E.; Prasad, K.N.; Ismail, A.; Tan, C.P.; Rajab, N.F. Revealing the power of the natural red pigment lycopene. Molecules 2010, 15, 959–987. [Google Scholar] [CrossRef] [PubMed]
- Strack, D.; Vogt, T.; Schliemann, W. Recent advances in betalain research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef] [PubMed]
- Strack, D.; Wray, V. The anthocyanins. In The Flavonoids Advances in Research Since 1986, 1st ed.; Harborne, J.B., Ed.; Chapman & Hall: London, UK, 1994; Volume 1, pp. 1–22. [Google Scholar]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Conner, P.J.; MacLean, D. Fruit anthocyanin profile and berry color of muscadine grape cultivars and Muscadinia germplasm. HortScience. 2013, 48, 1235–1240. [Google Scholar] [CrossRef]
- Xue, L.; Wang, Z.; Zhang, W.; Li, Y.; Wang, J.; Lei, J. Flower pigment inheritance and anthocyanin characterization of hybrids from pink-flowered and white-flowered strawberry. Sci. Hortic. 2016, 200, 143–150. [Google Scholar] [CrossRef]
- Lu, C.; Li, Y.; Wang, J.; Qu, J.; Chen, Y.; Chen, X.; Huang, H.; Dai, S. Flower color classification and correlation between color space values with pigments in potted multiflora chrysanthemum. Sci. Hortic. 2021, 283, 110082. [Google Scholar] [CrossRef]
- Jia, N.; Shu, Q.Y.; Wang, L.S.; Du, H.; Xu, Y.J.; Liu, Z.A. Analysis of petal anthocyanins to investigate coloration mechanism in herbaceous peony cultivars. Sci. Hortic. 2008, 117, 167–173. [Google Scholar] [CrossRef]
- Cesa, S.; Carradori, S.; Bellagamba, G.; Locatelli, M.; Casadei, M.A.; Masci, A.; Paolicelli, P. Evaluation of processing effects on anthocyanin content and colour modifications of blueberry (Vaccinium spp.) extracts: Comparison between HPLC-DAD and CIELAB analyses. Food Chem. 2017, 232, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Song, Y.; Jin, X.; Su, T.; Pu, Y. Effects of pigment constituents and their distribution on spathe coloration of Zantedeschia hybrida. HortScience 2017, 52, 1840–1848. [Google Scholar] [CrossRef]
- de Matos, M.F.R.; Bezerra, P.Q.M.; Correia, L.C.A.; Viola, D.N.; de Oliveira Rios, A.; Druzian, J.I.; Nunes, I.L. Innovative methodological approach using CIELab and dye screening for chemometric classification and HPLC for the confirmation of dyes in cassava flour: A contribution to product quality control. Food Chem. 2021, 365, 130446. [Google Scholar] [CrossRef] [PubMed]
- Punasiya, R.; Devre, K.; Pillai, S. Pharmacognostic and Pharmacological overview on Hibiscus syriacus L. Int. J. Pharm. Life Sci. 2014, 5, 3617–3621. [Google Scholar]
- Kim, Y.R.; Lee, S.Y.; Lee, S.M.; Shim, I.; Lee, M.Y. Effect of Hibiscus syriacus Linnaeus extract and its active constituent, saponarin, in animal models of stress-induced sleep disturbances and pentobarbital-induced sleep. Biomed. Pharmacother. 2022, 146, 112301. [Google Scholar] [CrossRef]
- Karunarathne, W.; Molagoda, I.; Park, S.; Kim, J.; Lee, O.-K.; Kwon, H.; Oren, M.; Choi, Y.; Ryu, H.; Oh, S.-R.; et al. Anthocyanins from Hibiscus syriacus L. inhibit melanogenesis by activating the ERK signaling pathway. Biomolecules 2019, 9, 645. [Google Scholar] [CrossRef]
- Kurtulbaş, E.; Pekel, A.G.; Bilgin, M.; Makris, D.P.; Şahin, S. Citric acid-based deep eutectic solvent for the anthocyanin recovery from Hibiscus sabdariffa through microwave-assisted extraction. Biomass Convers. Biorefin. 2020, 12, 351–360. [Google Scholar] [CrossRef]
- Ha, Y.M.; Kim, D.Y.; Shim, K.K. A new cultivar Hibiscus syriacus ‘Red Bohanjae’ with small violet-pink flowers for a pot plant. Hortic. Sci. Technol. 2015, 33, 292–297. [Google Scholar]
- Ha, Y.M.; Lim, K.B.; Shim, K.K. Development of a new hibiscus cultivar ’Daewangchun’ with vigorous growth and unique red eye through interspecific hybridization. Hortic. Sci. Technol. 2015, 33, 453–458. [Google Scholar] [CrossRef][Green Version]
- Kim, S.H.; Kim, D.S.; Kim, J.B.; Ha, B.K.; Lee, D.M.; Song, H.S.; Kang, S.Y. Hibiscus syriacus ‘Dasom’, a new flower-color mutant variety developed by radiation breeding. Hortic. Sci. Technol. 2015, 30, 298–301. [Google Scholar]
- Wang, X.; Li, L.; Liu, C.; Zhang, M.; Wen, Y. An integrated metabolome and transcriptome analysis of the Hibiscus syriacus L. petals reveal the molecular mechanisms of anthocyanin accumulation. Front. Genet. 2022, 13, 995748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, Y.; Chong, S.; Yan, S.; Yu, R.; Chen, R.; Si, J.; Zhang, X. Identification and quantitative analysis of anthocyanins composition and their stability from different strains of Hibiscus syriacus L. flowers. Ind. Crops Prod. 2022, 177, 114457. [Google Scholar] [CrossRef]
- Hong, Y.; Tang, X.; Huang, H.; Zhang, Y.; Dai, S. Transcriptomic analyses reveal species-specific light-induced anthocyanin biosynthesis in chrysanthemum. BMC Genom. 2015, 16, 202. [Google Scholar] [CrossRef] [PubMed]
- Molagoda, I.M.N.; Lee, K.T.; Choi, Y.H.; Kim, G.Y. Anthocyanins from Hibiscus syriacus L. inhibit oxidative stress-mediated apoptosis by activating the Nrf2/HO-1 signaling pathway. Antioxidants 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Lee, K.T.; Choi, Y.H.; Jin, C.Y.; Kim, G.Y. Anthocyanin-enriched polyphenols from Hibiscus syriacus L. (Malvaceae) exert anti-osteoporosis effects by inhibiting GSK-3β and subsequently activating β-catenin. Phytomedicine 2021, 91, 153721. [Google Scholar] [CrossRef]
- Giuliano, G.; Tavazza, R.; Diretto, G.; Beyer, P.; Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotechnol. 2008, 26, 139–145. [Google Scholar] [CrossRef]
- Fu, H.; Zeng, T.; Zhao, Y.; Luo, T.; Deng, H.; Meng, C.; Luo, J.; Wang, C. Identification of chlorophyll metabolism-and photosynthesis-related genes regulating green flower color in chrysanthemum by integrative transcriptome and weighted correlation network analyses. Genes 2021, 12, 449. [Google Scholar] [CrossRef]
- Bueno, J.M.; Sáez-Plaza, P.; Ramos-Escudero, F.; Jiménez, A.M.; Fett, R.; Asuero, A.G. Analysis and antioxidant capacity of anthocyanin pigments. Part II: Chemical structure, color, and intake of anthocyanins. Crit. Rev. Anal. Chem. 2012, 42, 126–151. [Google Scholar] [CrossRef]
- Meanchaipiboon, S.; Kobayashi, N.; Nakatsuka, A. Analyses of pigment compositions and anthocyanin biosynthesis gene expression in Hirado azalea cultivars. Hortic. J. 2020, 89, 284–291. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, W.; Xiang, W.; Wang, D.; Xue, B.; Liu, X.; Xing, L.; Wu, D.; Wang, S.; Guo, Q.; et al. Integrated metabolic profiling and transcriptome analysis of pigment accumulation in Lonicera japonica flower petals during color-transition. BMC Plant Bio. 2021, 21, 98. [Google Scholar]
- Yeon, J.Y.; Kim, W.S. Floral pigment-scent associations in eight cut rose cultivars with various petal colors. Hortic. Environ. Biotechnol. 2020, 61, 633–641. [Google Scholar] [CrossRef]
- Iwashina, T. Contribution to flower colors of flavonoids including anthocyanins: A review. Nat. Prod. Commun. 2015, 10, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hou, R.; Zhu, J.; Zhang, L. Transcriptome sequencing analysis of petals of two varieties of Hibiscus syriacus. MPB 2022, 20, 2507–2516. [Google Scholar]
- Boase, M.R.; Lewis, D.H.; Davies, K.M.; Marshall, G.B.; Patel, D.; Schwinn, K.E.; Deroles, S.C. Isolation and antisense suppression of flavonoid 3′,5′-hydroxylasemodifies flower pigments and colour in cyclamen. BMC Plant Bio. 2010, 10, 107. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).