
Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Treatments
2.2. Physiological and Biochemical Indexes and Photosynthetic Parameter Measurements
2.2.1. Physiological and Biochemical Measurements
2.2.2. Photosynthetic Parameter Measurements
2.3. Statistical Analysis
3. Results
3.1. Effect of MT on Photosynthetic Pigments and Water Status of Chrysanthemum Seedlings under Drought Stress
3.2. Effect of MT on Reactive Oxygen Species and the Antioxidant System of Chrysanthemum Seedlings under Drought Stress
3.3. Effect of MT on Organic Osmoregulatory Substances in Seedlings under Drought Stress
3.4. Effect of MT on Relative Plasma Membrane Permeability and Membrane Peroxidation in Chrysanthemum Seedlings under Drought Stress
3.5. Effect of Exogenous MT on Chrysanthemum Seedling Leaf Photosynthesis under Drought Stress
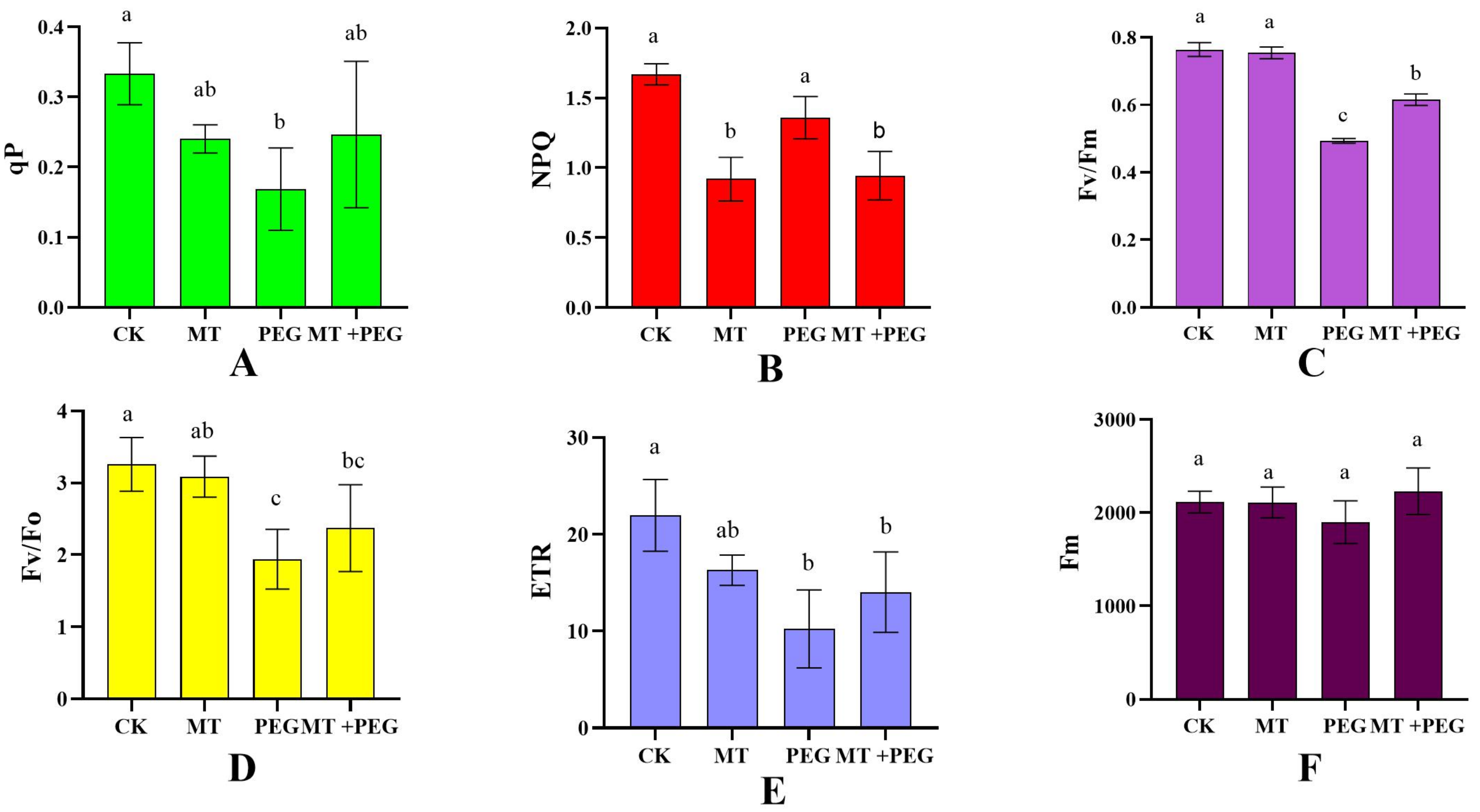
3.6. Effect of Exogenous MT on Chrysanthemum Seedling Chlorophyll Fluorescence Parameters under Drought Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Presowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 2008, 45, 24–31. [Google Scholar] [CrossRef]
- Xu, Y.J.; Zhao, X.; Aiwaili, P.; Mu, X.Y.; Zhao, M.; Zhao, J.; Cheng, L.N.; Ma, C.; Gao, J.P.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. Cell Mol. Biol. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Wang, Z.L.; Xi, L.Z.; Li, Y.H.; Dong, A.X. Characteristics of droughts in eastern Northwest China over the past 546 years. Acta Meteorol. Sin. 2020, 78, 72–85. [Google Scholar]
- Krisnayanti, D.S.; Bunganaen, W.; Frans, J.H.; Seran, Y.A.; Legono, D. Curve number estimation for ungauged watershed in semi-arid region. Civ. Eng. J. 2021, 7, 1070–1083. [Google Scholar] [CrossRef]
- Liu, X.; Fu, L.J.; Yu, J.; Zheng, N.Y.; Liu, B.D.; Ding, G.H. Mitigating effects of five exogenous substances on the growth of penstemon seedlings under drought stress. Northwest J. Bot. 2022, 42, 1169–1179. [Google Scholar]
- Xiao, S.; Liu, L.T.; Wang, H.; Li, D.X.; Bai, Z.Y.; Zhang, Y.J.; Sun, H.C.; Zhang, K.; Li, C.D. Exogenous melatonin accelerates seed germination in cotton (Gossypium hirsutum L.). PLoS ONE 2019, 4, e0216575. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Roles of exogenous glutathione in antioxidant defense system and methylglyoxal detoxification during salt stress in mung bean. Biologia Plantarum. 2015, 59, 745–756. [Google Scholar] [CrossRef]
- Ye, J.; Deng, X.P.; Wang, S.W.; Yin, L.N.; Chen, D.J.; Xiong, B.L.; Wang, X.Y. Effects of melatonin on growth, photosynthetic and antioxidant properties of wheat seedlings under drought stress. J. Wheat Crops 2015, 35, 1275–1283. [Google Scholar]
- Zhang, M.C.; He, S.Y.; Qin, B.; Wang, M.X.; Jin, X.J.; Ren, C.Y.; Wu, Y.K.; Zhang, Y.S. Effects of exogenous melatonin on morphology, photosynthetic physiology and yield of spring soybean variety Suinong 26 under drought stress. J. Crop Sci. 2021, 47, 1791–1805. [Google Scholar]
- Xing, X.J.; Ding, Y.R.; Jin, J.Y.; Song, A.P.; Chen, S.M.; Chen, F.D.; Fang, W.M.; Jiang, J.F. Physiological and transcripts analyses reveal the mechanism by which melatonin alleviates heat stress in chrysanthemum seedlings. Front. Plant Sci. 2021, 12, 673236. [Google Scholar] [CrossRef]
- Sharma, A.; Zheng, B. Melatonin Mediated Regulation of Drought Stress: Physiological and Molecular Aspects. Plants 2019, 8, 190. [Google Scholar] [CrossRef]
- Cui, G.B.; Zhao, X.X.; Liu, S.D.; Sun, F.L.; Zhang, C.; Xi, Y.J. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Zeng, L.; Cheng, Y.; Lu, G.Y.; Fu, G.P.; Ma, H.Q.; Liu, Q.Y.; Zhang, X.K.; Zou, X.L.; Li, C.H. Exogenous melatonin alleviates damage from drought stress in Brassica napus L. (rapeseed) seedlings. Acta Physiol. Plant. 2018, 40, 3. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Wu, M.J.; Wang, Y.Q.; Yan, Y.R.; Mao, Q.; Ren, J.J.; Ma, R.H.; Liu, A.R.; Chen, S.C. Melatonin alleviates iron stress by improving iron homeostasis, antioxidant defense and secondary metabolism in cucumber. Sci. Hortic. 2020, 265, 109205. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Zhou, Z.; Cruz, M.H.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting plants to survive and thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef]
- Shi, H.T.; Chan, Z.L. The cysteine2/histidine2-type transcription factor ZINC FINGER OF ARABIDOPSIS THALIANA 6-activated C-REPEAT-BINDING FACTOR pathway is essential for melatonin-mediated freezing stress resistance in Arabidopsis. J. Pineal Res. 2014, 57, 185–191. [Google Scholar] [CrossRef]
- Xu, X.D. Effect of Exogenous Melatonin on the Physiological Response of Cucumber Seedlings under High Temperature Stress. Master’s Thesis, Northwest Agriculture and Forestry University, Xianyang, China, 2010. [Google Scholar]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.X.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.Q.; Zhang, H.J.; Cao, Y.Y.; Weeda, S.; Ren, S.X.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Hernández, R.J.; Cano, A.; Arnao, M.B. Melatonin acts as a growth-stimulating compound in some monocot species: Melatonin as growth promoter in plants. J. Pineal Res. 2005, 39, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.W.; Liang, D.; Ma, F.W. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhao, B.; Zhang, H.J.; Weeda, S.; Yang, C.; Yang, Z.C.; Ren, S.X.; Guo, Y.D. Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J. Pineal Res. 2013, 54, 15–23. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.F.; Ma, F.W. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wang, G.; Wang, M.L.; Zhang, S.X. Exogenous Melatonin Improves Tolerance to Water Deficit by Promoting Cuticle Formation in Tomato Plants. Molecules 2018, 23, 1605. [Google Scholar] [CrossRef]
- Ye, J.; Wang, S.W.; Deng, X.P.; Yin, L.N.; Xiong, B.L.; Wang, X.Y. Melatonin increased maize (Zea mays L.) seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage. Acta Physiol. Plant. 2016, 38, 2. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S. Melatonin alleviates cold-induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Gao, J.F. Plant Physiology Laboratory Guide; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Zhao, S.J.; Xu, C.C.; Zou, Q.; Meng, Q.W. Improvement of the method for determination of malondialdehyde in plant tissues. Plant Physiol. Lett. 1994, 3, 207–210. [Google Scholar] [CrossRef]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Leng, F.; Sun, S.; Jing, Y.; Wang, F.; Wei, Q.; Wang, X. A rapid and sensitive method for determination of trace amounts of glucose by anthrone-sulfuric acid method. Bulg. Chem. Commun. 2016, 48, 109–113. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Wang, N.; Li, X.; Zhu, W.; Liu, Y.; Luo, L.; Liu, Y. Establishment of a method for the detection of endogenous hydrogen peroxide in Escherichia coli. Agric. Prod. Process. 2013, 23, 67–70. [Google Scholar]
- Feng, Y.H. Cloning and Expression Analysis of LmP5CS Gene from Lycium barbarum. Master’s Thesis, Tianjin University, Tianjin, China, 2013. [Google Scholar]
- Zhao, C.F.; Wang, C.G.; Li, H.J.; Zheng, X.H.; Yang, M.; Zhang, R.H. Effect of exogenous melatonin on photosynthesis of maize leaves under drought and rehydration conditions. J. Ecol. 2021, 41, 1431–1439. [Google Scholar]
- Li, D.; Shen, H.T.; Wang, Y.F.; Wang, Y.H.; Wang, L.J.; Zhao, S.M.; Liu, L. Effect of exogenous melatonin on photosynthetic carbon assimilation and endogenous hormones in tobacco seedlings under drought stress. J. Grass Ind. 2021, 30, 130–139. [Google Scholar]
- Hai, X.; Mi, J.Z.; Zhao, B.P.; Yan, W.K.; Liu, J.H.; Zhang, B.J. Effect of exogenous spermidine on growth and physiological characteristics of oat seedlings under salt stress. Northwest J. Bot. 2021, 41, 1003–1011. [Google Scholar]
- Du, Z.; Hou, W.; Wang, L.; Li, L.; Zhang, K.; Lu, Y.C. Effects of exogenous melatonin on maize seedlings under drought stress. Chin. Agron. Bull. 2020, 36, 14–19. [Google Scholar]
- Qin, Y.; Druzhinina, I.S.; Pan, X.Y.; Yuan, Z.L. Microbially Mediated Plant Salt Tolerance and Microbiome-based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.J.; Zhao, L.F.; Xing, H.F.; Luo, Y.X.; Wei, Z.X. Effect of endophytic bacteria on proline and malondialdehyde in wheat seedlings under salt stress. J. Ecol. 2020, 40, 3726–3737. [Google Scholar]
- Huang, B.; Chen, Y.E.; Zhao, Y.Q.; Ding, C.B.; Liao, J.Q.; Hu, C.; Zhou, L.J.; Zhang, Z.W.; Yuan, S.; Yuan, M. Exogenous melatonin alleviates oxidative damages and protects photosystem II in maize seedlings under drought stress. Front. Plant Sci. 2019, 10, 677. [Google Scholar] [CrossRef]
- Wang, Y.P.; Reiter, R.J.; Chan, Z.L. Phytomelatonin: A universal abiotic stress regulator. J. Exp. Bot. 2018, 69, 963–974. [Google Scholar] [CrossRef]
- Li, A.; Sun, W.L.; Li, L.; Sun, F. Effects of exogenous melatonin on drought stress in wheat. Effect of exogenous melatonin on the growth of Perilla seedlings under drought stress. Jiangsu Agric. Sci. 2018, 46, 70–73. [Google Scholar]
- Wang, Y.; Chen, L.K.; Zhu, X.L.; Mo, M.Y.; Xu, Y.; Chen, F. Effects of different heat stress treatments on antioxidant-related enzyme activities in Chrysanthemum. North Hortic. 2010, 9, 98–101. [Google Scholar]
- Qin, B.; Zhang, M.C.; He, S.Y.; Zhang, C.Y.; Wang, M.Y.; Jin, X.J.; Wang, M.X.; Zhang, Y.X.; Hu, G.H. Mitigating effect of melatonin immersion on drought stress during seed germination of soybean. Agric. Res. Arid. Reg. 2020, 38, 192–198. [Google Scholar]
- Song, W.J.; Zhao, Y.C.; Fang, S.H.; Wang, C.D.; Zhuo, Q.C.; Xie, Y.; Qiu, C.L.; Dong, J.X. Effect of potassium xanthate on water retention and osmoregulation capacity of roasted tobacco seedlings under drought stress. China Tob. Sci. 2017, 38, 61–66. [Google Scholar]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.P.; Wu, F.B. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. Int. J. Nat. Synth. Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Zhang, J.; Mason, A.S.; Wu, J.; Liu, S.; Zhang, X.C.; Luo, T.; Redden, B.; Batley, J.; Hu, L.Y.; Yan, G.J. Identification of putative candidate genes for water stress tolerance in canola (Brassica napus). Front. Plant Sci. 2015, 6, 01058. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.K.; Min, W.F.; Akhtar, M.; Lu, X.P.; Bai, X.R.; Zhang, Y.X.; Tian, L.; Li, P.F. Melatonin Enhances Drought Tolerance in Rice Seedlings by Modulating Antioxidant Systems, Osmoregulation, and Corresponding Gene Expression. Int. J. Mol. Sci. 2022, 23, 12075. [Google Scholar] [CrossRef]
- Wu, S.Z.; Liang, W.B.; Nie, D.L.; Bai, W.F.; Shen, S.Z.; Li, J.H.; Yu, L. Effect of high temperature stress on photosynthesis of different blueberry varieties. J. Cent. South Univ. For. Sci. Technol. 2017, 37, 18. [Google Scholar]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, J.J.; Sun, L.Q.; Huang, B.; Ding, C.B.; Gu, Y.; Liao, J.Q.; Hu, C.; Zhang, Z.W.; Yuan, S.; et al. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol. Plant. 2018, 164, 349–363. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Sci. 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Zhou, J.; Xiao, W.; Chen, X.D.; Gao, D.S.; Li, L. Effect of calcium amino acid spraying on photosynthetic performance, fruit calcium content and quality of golden pear. J. Shandong Agric. Univ. Nat. Sci. Ed. 2018, 49, 551–555. [Google Scholar]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Lukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Van Kooten, K.O.; Snel, J.F.H. The Use of Chlorophyll Nomenclature in Plant Stress Physiology. Photosynth 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Michal, K.; David, K.; Ladislav, N. On the Relation between the Non Photochemical Quenching of Chlorophyll Fluorescence and Photosystem I Light Harvesting Efficiency—A Repetitive Flash Fluorescence Induction Study. Photosynth 2001, 68, 571–576. [Google Scholar]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.; Mattoo, A. Photoprotection, Photoinhibition, Gene Regulation, and Environment; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Murchie, E.H.; Niyogi, K.K. Manipulation of photoprotection to improve plant photosynthesis. Plant Physiol. 2011, 155, 86–92. [Google Scholar] [CrossRef]
- Irshad, A.; Ur Rehman, R.N.; Dubey, S.; Khan, M.A.; Yang, P.Z.; Hu, T.M. Rhizobium inoculation and exogenous melatonin synergistically increased thermotolerance by improving antioxidant defense, photosynthetic efficiency, and nitro-oxidative homeostasis in Medicago truncatula. Front. Ecol. Evol. 2022, 2022, 945695. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Hu, T.; Huo, Y.; Wang, L.; Zhang, L.; Yan, R. Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress. Horticulturae 2023, 9, 106. https://doi.org/10.3390/horticulturae9010106
Luo Y, Hu T, Huo Y, Wang L, Zhang L, Yan R. Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress. Horticulturae. 2023; 9(1):106. https://doi.org/10.3390/horticulturae9010106
Chicago/Turabian StyleLuo, Yan, Taotao Hu, Yunyun Huo, Lingling Wang, Li Zhang, and Rui Yan. 2023. "Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress" Horticulturae 9, no. 1: 106. https://doi.org/10.3390/horticulturae9010106
APA StyleLuo, Y., Hu, T., Huo, Y., Wang, L., Zhang, L., & Yan, R. (2023). Effects of Exogenous Melatonin on Chrysanthemum Physiological Characteristics and Photosynthesis under Drought Stress. Horticulturae, 9(1), 106. https://doi.org/10.3390/horticulturae9010106