
Falanghina Grapevine (Vitis vinifera L.) Yield and Berry Quality under Different Pedoclimatic Conditions in Southern Italy


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
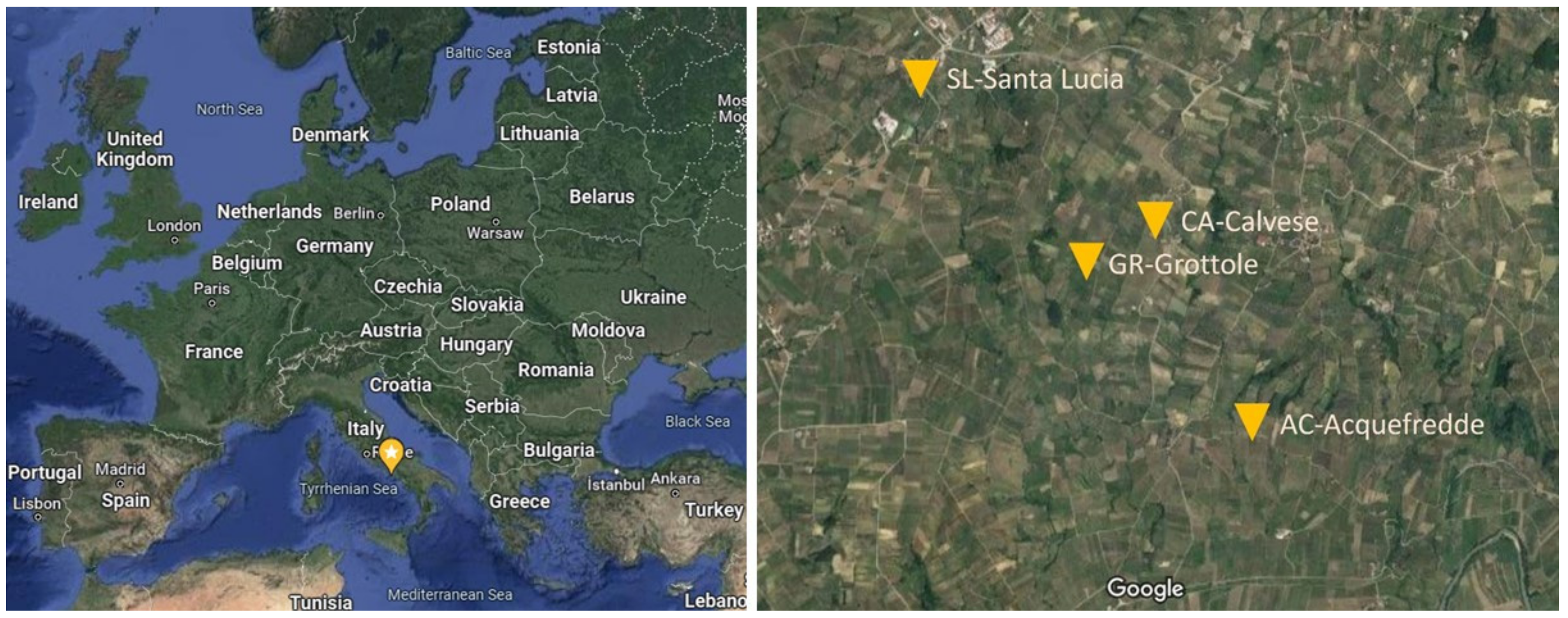
2.1. Study Site
2.2. Vegetative Growth and Yield Components, at Harvest
2.3. Berry and Must Quality Traits
2.4. Berry Mineral Composition
2.5. Data Elaboration
3. Results
3.1. Growth and Berry Quality Traits
3.2. Must Mineral and Organic Acids Composition
3.3. Must Chemical Analysis
3.4. Must Organic Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Rosas, I.; Deis, L.; Baldo, Y.; Cavagnaro, J.B.; Cavagnaro, P.F. High Temperature Alters Anthocyanin Concentration and Composition in Grape Berries of Malbec, Merlot, and Pinot Noir in a Cultivar-Dependent Manner. Plants 2022, 11, 926. [Google Scholar] [CrossRef]
- Pörtner, H.-; Roberts, D.; Tignor, M.; Poloczanska, E.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S.; Möller, V.; et al. Sixth Assessment Report Citations to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2022. [Google Scholar]
- Chmielewski, F.-M.; Rötzer, T. Response of Tree Phenology to Climate Change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Bonfante, A.; Monaco, E.; Langella, G.; Mercogliano, P.; Bucchignani, E.; Manna, P.; Terribile, F. A Dynamic Viticultural Zoning to Explore the Resilience of Terroir Concept under Climate Change. Sci. Total Environ. 2018, 624, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of Water Use Efficiency in Grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Deluc, L.G.; Quilici, D.R.; Decendit, A.; Grimplet, J.; Wheatley, M.D.; Schlauch, K.A.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water Deficit Alters Differentially Metabolic Pathways Affecting Important Flavor and Quality Traits in Grape Berries of Cabernet Sauvignon and Chardonnay. BMC Genom. 2009, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical Changes throughout Grape Berry Development and Fruit and Wine Quality. Food Glob. Sci. Books 2007, 1, 1–22. [Google Scholar]
- Oliveira, C.; Silva Ferreira, A.C.; Mendes Pinto, M.; Hogg, T.; Alves, F.; Guedes De Pinho, P. Carotenoid Compounds in Grapes and Their Relationship to Plant Water Status. J. Agric. Food Chem. 2003, 51, 5967–5971. [Google Scholar] [CrossRef]
- Cabral, I.L.; Teixeira, A.; Lanoue, A.; Unlubayir, M.; Munsch, T.; Valente, J.; Alves, F.; da Costa, P.L.; Rogerson, F.S.; Carvalho, S.M.P.; et al. Impact of Deficit Irrigation on Grapevine Cv. ‘Touriga Nacional’ during Three Seasons in Douro Region: An Agronomical and Metabolomics Approach. Plants 2022, 11, 732. [Google Scholar] [CrossRef]
- Tombesi, S.; Sabbatini, P.; Frioni, T.; Grisafi, F.; Barone, F.; Zani, P.; Palliotti, A.; Poni, S. Grapevine Response to Stress Generated by Excessive Temperatures during the Budburst. Horticulturae 2022, 8, 187. [Google Scholar] [CrossRef]
- Greer, D.H.; Weedon, M.M. Modelling Photosynthetic Responses to Temperature of Grapevine (Vitis vinifera Cv. Semillon) Leaves on Vines Grown in a Hot Climate. Plant Cell Environ. 2012, 35, 1050–1064. [Google Scholar] [CrossRef]
- Zarrouk, O.; Brunetti, C.; Egipto, R.; Pinheiro, C.; Genebra, T.; Gori, A.; Lopes, C.M.; Tattini, M.; Chaves, M.M. Grape Ripening Is Regulated by Deficit Irrigation/Elevated Temperatures According to Cluster Position in the Canopy. Front. Plant Sci. 2016, 7, 1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, A.K.; de Cortázar-Atauri, I.G.; van Leeuwen, C.; Chuine, I. General Phenological Model to Characterise the Timing of Flowering and Veraison of Vitis vinifera L. Aust. J. Grape Wine Res. 2011, 17, 206–216. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Destrac-Irvine, A. Modified Grape Composition under Climate Change Conditions Requires Adaptations in the Vineyard. Oeno One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of Temperature on Anthocyanin Biosynthesis in Grape Berry Skins.Aust. J. Grape Wine Res. 2006, 57, 54. [Google Scholar]
- Sweetman, C.; Deluc, L.G.; Cramer, G.R.; Ford, C.M.; Soole, K.L. Regulation of Malate Metabolism in Grape Berry and Other Developing Fruits. Phytochemistry 2009, 70, 1329–1344. [Google Scholar] [CrossRef] [PubMed]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of Sunlight and Temperature Effects on the Composition of Vitis vinifera Cv. Merlot Berries. Aust. J. Grape Wine Res. 2002, 53, 171–182. [Google Scholar]
- Sadras, V.O.; Petrie, P.R.; Moran, M.A. Effects of Elevated Temperature in Grapevine. II Juice PH, Titratable Acidity and Wine Sensory Attributes. Aust. J. Grape Wine Res. 2013, 19, 107–115. [Google Scholar] [CrossRef]
- Sugiura, T.; Shiraishi, M.; Konno, S.; Sato, A. Prediction of Skin Coloration of Grape Berries from Air Temperature. J. Hortic. 2018, 87, 18–25. [Google Scholar] [CrossRef]
- Gambetta, J.M.; Holzapfel, B.P.; Stoll, M.; Friedel, M. Sunburn in Grapes: A Review. Front. Plant Sci. 2021, in press. [CrossRef]
- Hulands, S.; Greer, D.H.; Harper, J.D.I. The Interactive Effects of Temperature and Light Intensity on Vitis vinifera Cv. “Semillon” Grapevines. II. Berry Ripening and Susceptibility to Sunburn at Harvest. Europ. J. Hortic. Sci. 2014, 79, 1–7. [Google Scholar]
- Pérez-Álvarez, E.P.; Intrigliolo Molina, D.S.; Vivaldi, G.A.; García-Esparza, M.J.; Lizama, V.; Álvarez, I. Effects of the Irrigation Regimes on Grapevine Cv. Bobal in a Mediterranean Climate: I. Water Relations, Vine Performance and Grape Composition. Agric. Water Manag. 2021, 248, 106772. [Google Scholar] [CrossRef]
- Boselli, M.; Corso, C.; Monaco, A. Ampelographic Characterization of White Grape Varieties in Campania (Southern Italy) by Multivariate Analysis. Acta Hortic. 2000, 528, 75–81. [Google Scholar] [CrossRef]
- Di Vaio, C.; Villano, C.; Lisanti, M.T.; Marallo, N.; Cirillo, A.; di Lorenzo, R.; Pisciotta, A. Application of Anti-Transpirant to Control Sugar Accumulation in Grape Berries and Alcohol Degree in Wines Obtained from Thinned and Unthinned Vines of Cv. Falanghina (Vitis vinifera L.). Agronomy 2020, 10, 345. [Google Scholar] [CrossRef]
- de Filippis, F.; Aponte, M.; Piombino, P.; Lisanti, M.T.; Moio, L.; Ercolini, D.; Blaiotta, G. Influence of Microbial Communities on the Chemical and Sensory Features of Falanghina Sweet Passito Wines. Food Res. Int. 2019, 120, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Damiano, N.; Altieri, S.; Battipaglia, G.; de Micco, V. Comparing Methods for the Analysis of Δ13C in Falanghina Grape Must from Different Pedoclimatic Conditions. Horticulturae 2022, 8, 226. [Google Scholar] [CrossRef]
- Damiano, N.; Arena, C.; Bonfante, A.; Caputo, R.; Erbaggio, A.; Cirillo, C.; de Micco, V. How Leaf Vein and Stomata Traits Are Related with Photosynthetic Efficiency in Falanghina Grapevine in Different Pedoclimatic Conditions. Plants 2022, 11, 1507. [Google Scholar] [CrossRef]
- Terribile, F.; Gennaro, A.D.; Mascellis, R.D. Carta dei suoli della Valle Telesina (1:50,000). In Progetto UOT Relazione Finale Convenzione CNR-ISPAIM-Regione Campania Assessorato Alla Agricoltura; Raccolta di 10 Carte Pedologiche della Regione Campania; Stampa System Cart: Rome, Italy, 1996. [Google Scholar]
- Cirillo, C.; Arena, C.; Rouphael, Y.; Caputo, R.; Amitrano, C.; Petracca, F.; de Francesco, S.; Vitale, E.; Erbaggio, A.; Bonfante, A.; et al. Counteracting the Negative Effects of Copper Limitations Through the Biostimulatory Action of a Tropical Plant Extract in Grapevine Under Pedo-Climatic Constraints. Front. Environ. Sci. 2021, 9, 587550. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Forlani, M.; Basile, B. Influence of Intensity of Post-Veraison Defoliation or Shoot Trimming on Vine Physiology, Yield Components, Berry and Wine Composition in Aglianico Grapevines. Aust. J. Grape Wine Res. 2017, 23, 226–239. [Google Scholar] [CrossRef]
- European Union [EU] Commission Regulation (EEC) No 2676/90(REGULATION, H.A.T. 1990. Commission Regulation (EEC) No. 2676/90 Community Methods for the Analysis of Wines Official Journal L, 3 October 1990; European Union: Brussels, Belgium, 1990; pp. 1–192. [Google Scholar]
- Zhifeng, G.; Chengguang, F. Determination of organic acids and inorganic anions by single-column ion chromatography. J. Instrum. Anal. 1994, 5, 32–36. [Google Scholar]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the Relationship Between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Funct. Plant Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Rodrigues, P.; Pedroso, V.; Gonçalves, F.; Reis, S.; Santos, J.A. Temperature-Based Grapevine Ripeness Modeling for Cv. Touriga Nacional and Encruzado in the Dão Wine Region, Portugal. Agronomy 2021, 11, 1777. [Google Scholar] [CrossRef]
- Cirilli, M.; Bellincontro, A.; de Santis, D.; Botondi, R.; Colao, M.C.; Muleo, R.; Mencarelli, F. Temperature and Water Loss Affect ADH Activity and Gene Expression in Grape Berry during Postharvest Dehydration. Food Chem. 2012, 132, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.C.; Šuklje, K.; Antalick, G.; Schmidtke, L.M.; Blackman, J.W. Late-Season Shiraz Berry Dehydration That Alters Composition and Sensory Traits of Wine. J. Agric. Food Chem. 2018, 66, 7750–7757. [Google Scholar] [CrossRef] [PubMed]
- Ollat, N.; Cookson, S.J.; Destrac-Irvine, A.; Lauvergeat, V.; Ouaked-Lecourieux, F.; Marguerit, E.; Barrieu, F.; Dai, Z.; Duchêne, E.; Gambetta, G.A.; et al. Grapevine Adaptation to Abiotic Stress: An Overview. Acta Hortic. 2019, 1248, 497–512. [Google Scholar] [CrossRef]
- Rienth, M.; Torregrosa, L.; Sarah, G.; Ardisson, M.; Brillouet, J.M.; Romieu, C. Temperature Desynchronizes Sugar and Organic Acid Metabolism in Ripening Grapevine Fruits and Remodels Their Transcriptome. BMC Plant Biol. 2016, 16, 164. [Google Scholar] [CrossRef]
- Zhu, J.; Gou, F.; Rossouw, G.; Begum, F.; Henke, M.; Johnson, E.; Holzapfel, B.; Field, S.; Seleznyova, A. Simulating Organ Biomass Variability and Carbohydrate Distribution in Perennial Fruit Crops: A Comparison between the Common Assimilate Pool and Phloem Carbohydrate Transport Models. Silico Plants 2021, 3, diab024. [Google Scholar] [CrossRef]
- Li, Y.M.; Forney, C.; Bondada, B.; Leng, F.; Xie, Z. sen The Molecular Regulation of Carbon Sink Strength in Grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 11, 606918. [Google Scholar] [CrossRef]
- Pandey, R. Mineral Nutrition of Plants. In Plant Biology and Biotechnology: Plant Diversity, Organization, Function and Improvement; Springer: New Delhi, India, 2015; Volume 1, pp. 499–538. ISBN 9788132222866. [Google Scholar]
- Brataševec, K.; Sivilotti, P.; Vodopivec, B.M. Soil and Foliar Fertilization Affects Mineral Contents in Vitis vinifera L. Cv. “rebula” Leaves. J. Soil Sci. Plant Nutr. 2013, 13, 650–663. [Google Scholar]
- He, Z.L.; Yang, X.E.; Stoffella, P.J. Trace Elements in Agroecosystems and Impacts on the Environment. J. Trace Elem. Med. Biol. 2005, 19, 125–140. [Google Scholar] [CrossRef]
- Baggett, J.P.; Habibsadeh, S.; Toups, H.S.; Cochetel, N.; Ghan, R.; Robinson, M.L.; Barrios-Masias, F.H.; Cramer, G.R. Is Foliar Cl- Concentration the Cause of Photosynthetic Decline in Grapevine during Mild Salinity? Oeno One 2021, 55, 33–48. [Google Scholar] [CrossRef]
- Munns, R. Comparative Physiology of Salt and Water Stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Sadras, V.O.; Moran, M.A. Elevated Temperature Decouples Anthocyanins and Sugars in Berries of Shiraz and Cabernet Franc. Aust. J. Grape Wine Res. 2012, 18, 115–122. [Google Scholar] [CrossRef]
- Pinasseau, L.; Vallverdú-Queralt, A.; Verbaere, A.; Roques, M.; Meudec, E.; le Cunff, L.; Péros, J.P.; Ageorges, A.; Sommerer, N.; Boulet, J.C.; et al. Cultivar Diversity of Grape Skin Polyphenol Composition and Changes in Response to Drought Investigated by LC-MS Based Metabolomics. Front. Plant Sci. 2017, 8, 1826. [Google Scholar] [CrossRef] [PubMed]
- Palliotti, A.; Tombesi, S.; Silvestroni, O.; Lanari, V.; Gatti, M.; Poni, S. Changes in Vineyard Establishment and Canopy Management Urged by Earlier Climate-Related Grape Ripening: A Review. Sci. Hortic. 2014, 178, 43–54. [Google Scholar] [CrossRef]
- Coombe, B.G.; Iland, P.G. Grape Berry Development and Winegrape Quality. Viticulture 2004, 1, 210–248. [Google Scholar]
- Lund, S.; Bohlmann, J. The Molecular Basis for Wine Grape Quality-A Volatile Subject. Science 2006, 311, 804–805. [Google Scholar] [CrossRef] [Green Version]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight Exposure and Temperature Effects on Berry Growth and Composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. Aust. J. Grape Wine Res. 2001, 52, 1–7. [Google Scholar]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of Anthocyanins in Red-Wine Grape under High Temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef]
- Teixeira, A.; Eiras-Dias, J.; Castellarin, S.D.; Gerós, H. Berry Phenolics of Grapevine under Challenging Environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Total Leaf Area | Yield | Bunch Number | Actual Fertility | Y/LA | SSC | pH | TA | |
|---|---|---|---|---|---|---|---|---|
| m2 vine−1 | kg vine−1 | n° vine−1 | Kg m−2 | °Brix | g l−1 tartaric acid equivalent | |||
| Field (F) | ||||||||
| SL | 7.87 ± 0.60 a | 6.92 ± 0.28 a | 23.2 ± 1.20 a | 1.06 ± 0.030 b | 1.12 ± 0.12 b | 16.3 ± 0.32 c | 3.16 ± 0.076 a | 6.26 ± 0.615 a |
| CA | 5.04 ± 0.30 c | 2.25 ± 0.15 d | 13.8 ± 0.65 d | 0.92 ± 0.037 c | 0.53 ± 0.03 c | 19.3 ± 0.45 a | 3.30 ± 0.082 a | 5.47 ± 0.600 a |
| GR | 6.30 ± 0.46 b | 3.65 ± 0.23 c | 20.4 ± 0.99 b | 1.06 ± 0.034 b | 0.67 ± 0.09 c | 17.8 ± 0.47 b | 3.39 ± 0.079 a | 5.61 ± 0.572 a |
| AC | 3.79 ± 0.22 d | 4.37 ± 0.20 b | 17.1 ± 0.81 c | 1.23 ± 0.053 a | 1.40 ± 0.10 a | 18.4 ± 0.57 ab | 3.22 ± 0.042 a | 5.76 ± 0.443 a |
| Year (Y) | ||||||||
| 2019 | 7.14 ± 0.48 a | 4.85 ± 0.27 a | 17.7 ± 0.60 b | 1.22 ± 0.034 a | 0.89 ± 0.07 b | 20.0 ± 0.40 a | 3.26 ± 0.070 ab | 7.62 ± 0.603 a |
| 2020 | 5.49 ± 0.22 b | 3.56 ± 0.23 b | 14.2 ± 0.59 c | 0.80 ± 0.031 b | 0.66 ± 0.03 b | 17.8 ± 0.44 b | 3.16 ± 0.033 b | 5.63 ± 0.195 b |
| 2021 | 3.50 ± 0.24 c | 4.48 ± 0.29 a | 23.9 ± 1.04 a | 1.18 ± 0.029 a | 1.56 ± 0.16 a | 16.0 ± 0.32 c | 3.38 ± 0.073 a | 4.07 ± 0.186 c |
| Significance 1 | ||||||||
| F | ** | *** | *** | *** | *** | *** | NS | NS |
| Y | *** | *** | *** | *** | *** | *** | * | * |
| F × Y | NS | *** | *** | *** | *** | *** | NS | NS |
| Na⁺ | K⁺ | Mg2⁺ | Ca2⁺ | SO₄2− | PO₄3− | |
|---|---|---|---|---|---|---|
| (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | |
| Field (F) | ||||||
| SL | 0.150 ± 0.024 a | 0.97 ± 0.04 c | 0.066 ± 0.004 b | 0.081 ± 0.003 bc | 0.034 ± 0.002 b | 0.101 ± 0.007 c |
| CA | 0.091 ± 0.030 a | 1.60 ± 0.10 a | 0.089 ± 0.005 a | 0.128 ± 0.016 a | 0.045 ± 0.004 a | 0.174 ± 0.012 a |
| GR | 0.106 ± 0.021 a | 1.34 ± 0.07 b | 0.065 ± 0.006 b | 0.086 ± 0.007 b | 0.045 ± 0.003 a | 0.128 ± 0.009 bc |
| AC | 0.097 ± 0.016 a | 1.38 ± 0.04 b | 0.068 ± 0.008 b | 0.068 ± 0.006 c | 0.035 ± 0.003 b | 0.156 ± 0.014 ab |
| Year (Y) | ||||||
| 2019 | 0.091 ± 0.023 a | 1.31 ± 0.09 a | 0.064 ± 0.002 b | 0.069 ± 0.004 b | 0.033 ± 0.002 b | 0.140 ± 0.012 ab |
| 2020 | 0.129 ± 0.019 a | 1.31 ± 0.08 a | 0.093 ± 0.003 a | 0.101 ± 0.005 a | 0.042 ± 0.002 a | 0.155 ± 0.012 a |
| 2021 | 0.113 ± 0.020 a | 1.35 ± 0.09 a | 0.059 ± 0.006 b | 0.103 ± 0.015 a | 0.044 ± 0.004 a | 0.123 ± 0.011 b |
| Significance 1 | ||||||
| F | NS | *** | *** | *** | ** | *** |
| Y | NS | NS | *** | *** | ** | * |
| F × Y | NS | ** | ** | *** | NS | NS |
| Malate | Tartrate | Citrate | Isocitrate | |
|---|---|---|---|---|
| (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | |
| Field (F) | ||||
| SL | 2.46 ± 0.37 a | 6.64 ± 0.27 c | 0.358 ± 0.011 b | 0.079 ± 0.011 b |
| CA | 1.59 ± 0.33 b | 10.0 ± 1.18 a | 0.371 ± 0.028 b | 0.098 ± 0.011 a |
| GR | 1.86 ± 0.25 b | 7.63 ± 0.43 b | 0.458 ± 0.028 a | 0.100 ± 0.013 a |
| AC | 2.76 ± 0.22 a | 6.48 ± 0.26 c | 0.373 ± 0.018 b | 0.083 ± 0.012 b |
| Year (Y) | ||||
| 2019 | 3.04 ± 0.24 a | 7.18 ± 0.42 b | 0.430 ± 0.024 a | 0.101 ± 0.007 b |
| 2020 | 1.83 ± 0.12 b | 6.60 ± 0.25 b | 0.403 ± 0.022 a | 0.119 ± 0.006 a |
| 2021 | 1.63 ± 0.28 b | 9.29 ± 0.94 a | 0.338 ± 0.009 b | 0.051 ± 0.003 c |
| Significance 1 | ||||
| F | ** | *** | *** | * |
| Y | *** | *** | *** | *** |
| F × Y | NS | *** | ** | * |
| Reducing Sugars | pH | YAN | Calcium | Catechins | Total Polyphenols | |
|---|---|---|---|---|---|---|
| g/L | mg/L | mg/L | mg/L | ppm | ||
| Field (F) | ||||||
| SL | 201 ± 4.90 b | 3.11 ± 0.01 c | 88.4 ± 15.7 b | 58.8 ± 7.49 bc | 5.49 ± 1.24 a | 628.5 ± 254.2 b |
| CA | 230 ± 12.07 a | 3.14 ± 0.05 bc | 81.0 ± 10.2 b | 103 ± 16.4 a | 8.78 ± 1.61 a | 1011 ± 233.5 a |
| GR | 213 ± 6.88 ab | 3.21 ± 0.04 b | 144 ± 22.8 a | 66.0 ± 7.02 b | 6.98 ± 1.35 a | 708.2 ± 218.8 b |
| AC | 208 ± 25.47 b | 3.32 ± 0.05 a | 82.0 ± 13.0 b | 46.6 ± 9.01 c | 7.47 ± 1.48 a | 854.4 ± 120.4 ab |
| Year (Y) | ||||||
| 2019 | 196 ± 16.08 b | 3.14 ± 0.03 b | 117 ± 17.5 a | 81.5 ± 15.4 a | 7.40 ± 1.23 a | 1467 ± 123.6 a |
| 2020 | 236 ± 8.60 a | 3.32 ± 0.04 c | 125 ± 11.3 a | 42.4 ± 6.13 b | 6.49 ± 1.19 a | 543.4 ± 130.0 b |
| 2021 | 207 ± 9.72 b | 3.13 ± 0.03 a | 53.7 ± 6.39 b | 82.3 ± 2.01 a | 7.65 ± 1.35 a | 391.3 ± 105.1 b |
| Significance 1 | ||||||
| F | * | *** | *** | *** | NS | * |
| Y | *** | *** | *** | *** | NS | *** |
| F × Y | *** | NS | * | *** | NS | *** |
| Titratable Acidity | Volatile Acidity | Malic Acid | Gluconic Acid | Citric Acid | Tartaric Acid | |
|---|---|---|---|---|---|---|
| g/L | g/L | g/L | g/L | g/L | g/L | |
| Field (F) | ||||||
| SL | 6.80 ± 0.44 a | 0.067 ± 0.012 b | 2.81 ± 0.36 a | 0.350 ± 0.033 a | 0.290 ± 0.023 b | 5.09 ± 0.24 b |
| CA | 6.56 ± 0.49 a | 0.091 ± 0.018 a | 1.93 ± 0.13 c | 0.462 ± 0.088 a | 0.396 ± 0.017 a | 6.06 ± 0.56 a |
| GR | 6.34 ± 0.37 a | 0.064 ± 0.111 b | 2.53 ± 0.29 ab | 0.281 ± 0.058 a | 0.285 ± 0.042 b | 5.38 ± 0.26 b |
| AC | 4.64 ± 0.44 b | 0.082 ± 0.022 ab | 2.33 ± 0.23 b | 0.408 ± 0.111 a | 0.161 ± 0.054 b | 3.22 ± 0.17 c |
| Year (Y) | ||||||
| 2019 | 6.88 ± 0.58 a | 0.233 ± 0.004 b | 2.98 ± 0.30 a | 0.436 ± 0.079 a | 0.340 ± 0.037 a | 5.47 ± 0.52 a |
| 2020 | 5.12 ± 0.27 b | 0.127 ± 0.009 a | 1.97 ± 0.11 b | 0.348 ± 0.067 a | 0.360 ± 0.026 a | 4.07 ± 0.20 b |
| 2021 | 6.26 ± 0.24 a | 0.078 ± 0.007 c | 2.23 ± 0.18 b | 0.343 ± 0.059 a | 0.276 ± 0.036 b | 5.28 ± 0.38 a |
| Significance 1 | ||||||
| F | *** | * | *** | NS | *** | *** |
| Y | *** | *** | *** | NS | ** | *** |
| F × Y | *** | * | *** | NS | *** | ** |
| Year 2019 | Year 2020 | Year 2021 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| r | s 1 | r | s 1 | r | s 1 | ||||||
| BW | TLA | −0.111 | NS | BW | TLA | 0.585 | **** | BW | TLA | 0.054 | NS |
| BW | SSC | −0.749 | **** | BW | SSC | −0.198 | NS | BW | SSC | −0.615 | **** |
| BW | pH | −0.551 | **** | BW | pH | −0.475 | ** | BW | pH | 0.226 | NS |
| BW | GLU | −0.421 | NS | BW | GLU | −0.112 | NS | BW | GLU | −0.051 | NS |
| BW | CIT | −0.359 | NS | BW | CIT | 0.001 | NS | BW | CIT | −0.650 | *** |
| BW | ANT | −0.555 | ** | BW | ANT | −0.427 | NS | BW | ANT | −0.216 | NS |
| SSC | pH | 0.546 | **** | SSC | pH | 0.620 | **** | SSC | pH | −0.252 | NS |
| SSC | TA | 0.180 | NS | SSC | TA | −0.368 | * | SSC | TA | 0.327 | NS |
| SSC | MAL | −0.386 | NS | SSC | MAL | −0.497 | * | SSC | MAL | 0.642 | *** |
| SSC | GLU | 0.390 | NS | SSC | GLU | 0.206 | NS | SSC | GLU | 0.688 | **** |
| SSC | CIT | 0.373 | NS | SSC | CIT | 0.718 | **** | SSC | CIT | 0.860 | **** |
| SSC | TAR | 0.394 | NS | SSC | TAR | −0.683 | **** | SSC | TAR | −0.374 | NS |
| SSC | TPO | 0.320 | NS | SSC | TPO | 0.864 | **** | SSC | TPO | 0.791 | **** |
| pH | MAL | −0.017 | NS | pH | MAL | 0.197 | ** | pH | MAL | −0.064 | NS |
| pH | GLU | −0.010 | NS | pH | GLU | −0.188 | ** | pH | GLU | −0.209 | NS |
| pH | CIT | 0.464 | * | pH | CIT | 0.600 | ** | pH | CIT | 0.090 | NS |
| pH | TAR | 0.607 | ** | pH | TAR | −0.644 | *** | pH | TAR | −0.239 | NS |
| pH | TPO | 0.450 | * | pH | TPO | 0.628 | *** | pH | TPO | 0.024 | NS |
| TA | CIT | 0.672 | **** | TA | CIT | −0.102 | NS | TA | CIT | 0.150 | NS |
| TA | TAR | 0.686 | **** | TA | TAR | 0.255 | NS | TA | TAR | 0.718 | **** |
| TA | TPO | 0.760 | **** | TA | TPO | −0.329 | NS | TA | TPO | −0.253 | NS |
| MAL | GLU | −0.021 | NS | MAL | GLU | −0.267 | NS | MAL | GLU | 0.888 | **** |
| MAL | CIT | 0.551 | ** | MAL | CIT | −0.001 | NS | MAL | CIT | 0.494 | * |
| MAL | TAR | 0.386 | NS | MAL | TAR | −0.030 | NS | MAL | TAR | −0.718 | **** |
| MAL | TPO | 0.429 | NS | MAL | TPO | −0.301 | NS | MAL | TPO | 0.572 | ** |
| GLU | CIT | 0.257 | NS | GLU | CIT | 0.432 | NS | GLU | CIT | 0.596 | ** |
| GLU | TAR | 0.374 | NS | GLU | TAR | 0.028 | NS | GLU | TAR | −0.579 | ** |
| GLU | TPO | 0.561 | ** | GLU | TPO | 0.391 | NS | GLU | TPO | 0.534 | ** |
| CIT | TAR | 0.943 | **** | CIT | TAR | −0.658 | *** | CIT | TAR | −0.118 | NS |
| CIT | TPO | 0.946 | **** | CIT | TPO | 0.851 | **** | CIT | TPO | 0.741 | **** |
| TAR | TPO | 0.942 | **** | TAR | TPO | −0.582 | ** | TAR | TPO | −0.338 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damiano, N.; Cirillo, C.; Petracca, F.; Caputo, R.; Erbaggio, A.; Giulioli, M.; De Micco, V. Falanghina Grapevine (Vitis vinifera L.) Yield and Berry Quality under Different Pedoclimatic Conditions in Southern Italy. Horticulturae 2022, 8, 829. https://doi.org/10.3390/horticulturae8090829
Damiano N, Cirillo C, Petracca F, Caputo R, Erbaggio A, Giulioli M, De Micco V. Falanghina Grapevine (Vitis vinifera L.) Yield and Berry Quality under Different Pedoclimatic Conditions in Southern Italy. Horticulturae. 2022; 8(9):829. https://doi.org/10.3390/horticulturae8090829
Chicago/Turabian StyleDamiano, Nicola, Chiara Cirillo, Francesca Petracca, Rosanna Caputo, Arturo Erbaggio, Marco Giulioli, and Veronica De Micco. 2022. "Falanghina Grapevine (Vitis vinifera L.) Yield and Berry Quality under Different Pedoclimatic Conditions in Southern Italy" Horticulturae 8, no. 9: 829. https://doi.org/10.3390/horticulturae8090829
APA StyleDamiano, N., Cirillo, C., Petracca, F., Caputo, R., Erbaggio, A., Giulioli, M., & De Micco, V. (2022). Falanghina Grapevine (Vitis vinifera L.) Yield and Berry Quality under Different Pedoclimatic Conditions in Southern Italy. Horticulturae, 8(9), 829. https://doi.org/10.3390/horticulturae8090829