
Air-Pruning Containers Modify Root and Scion Growth and Alter Resource Allocation of Bench-Grafted Apple Plants


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Irrigation and Fertilization
2.3. Sampling
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bravin, E.; Kilchenmann, A.; Leumann, M. Six hypotheses for profitable apple production based on the economic work-package within the ISAFRUIT Project. J. Hort. Sci. Biotechnol. 2009, 84, 164–167. [Google Scholar] [CrossRef]
- DeMarree, A.; Robinson, T.; Hoying, S. Economics and the orchard system decision. Compact. Fruit Tree 2003, 36, 42–49. [Google Scholar]
- Goedegebure, J. Economic aspects of super-intensive apple orchards. Acta Hortic. 1993, 349, 285–294. [Google Scholar] [CrossRef]
- Heijerman, G.; Roelofs, P.; Groot, M. Profitability of the Dutch growing system of ‘Conference’. Acta Hortic. 2015, 1094, 233–238. [Google Scholar] [CrossRef]
- Robinson, T.L. Effects of tree density and tree shape on apple orchard performance. Acta Hortic. 2007, 732, 405–414. [Google Scholar] [CrossRef]
- Robinson, T.L.; Hoying, S.A.; Reginato, G.H. The tall spindle planting system: Principles and performance. Acta Hortic. 2011, 903, 571–579. [Google Scholar] [CrossRef]
- Gallardo, K.; Galinato, S. 2019 Cost Estimates of Establishing, Producing, and Packing ‘Honeycrisp’ Apples in Washington; Washington State University: Pullman, WA, USA, 2020; Available online: http://ses.wsu.edu/wp-content/uploads/2020/11/TB70E.pdf (accessed on 15 July 2022).
- Robinson, T.; Hoying, S.; Sazo, M.M.; Marree, A.D.; Dominguez, L. A vision for apple orchard systems of the future. NY Fruit Q. 2013, 21, 11–16. [Google Scholar]
- Rune, G. Slits in container wall improve root structure and stem straightness of out planted Scots pine seedlings. Silva Fenn. 2003, 37, 333–342. [Google Scholar] [CrossRef][Green Version]
- Ortega, U.; Majada, J.; Mena-Petite, A.; Sanchez-Zabala, J.; Rodriguez Iturrizar, N.; Txarterina, K.; Azpitarte, J.; Duñabeitia, M. Field performance of Pinus radiata D. Don produced in nursery with different types of containers. New Forest 2006, 31, 97–112. [Google Scholar] [CrossRef]
- Van Sambeek, J.W.; Godsey, L.D.; Walter, W.D.; Garrett, H.E.; Dwyer, J.P. Field performance of Quercus bicolor established as repeatedly air-root-pruned container and bareroot planting stock. Open J. For. 2016, 6, 163–176. [Google Scholar]
- Pregitzer, K.S.; Laskowski, M.J.; Butron, A.J.; Lessard, V.C.; Zak, D.R. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Rytter, R.-M. The effect of limited availability of N or water on C allocation to fine roots and annual fine root turnover in Alnus incana and Salix viminalis. Tree Physiol. 2013, 33, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Makkonen, K.; Helmisaari, H.S. Assessing fine-root biomass and production in a Scots pine stand—Comparison of soil core and root in-growth core methods. Plant Soil 1999, 210, 43–50. [Google Scholar] [CrossRef]
- Noguchi, K.; Sakata, T.; Mizoguchi, T.; Takahashi, M. Estimating the production and mortality of fine roots in a Japanese cedar (Cryptomeria japonica D. Don) plantation using a minirhizotron technique. J. For. Res. 2005, 10, 435–441. [Google Scholar] [CrossRef]
- An, H.; Luo, F.; Wu, T.; Wang, Y.; Xu, X.; Zhang, X.; Han, Z. Dwarfing effect of apple rootstocks is intimately associated with low number of fine roots. HortScience 2017, 52, 503–512. [Google Scholar] [CrossRef]
- Hooijdonk, B.v.; Woolley, D.; Warrington, I.; Tustin, S. Rootstocks Modify Scion Architecture, Endogenous Hormones, and Root Growth of Newly Grafted ‘Royal Gala’ Apple Trees. J. Am. Soc. Hort. Sci. 2011, 136, 93–102. [Google Scholar] [CrossRef]
- Liu, A.; Abdelfattah, A.; Wasserman, B.; Wisniewski, M.; Droby, M.; Fazio, G.; Mazzola, M.; Wu, X. Contrasting effects of genotype and root size on the fungal and bacterial communities associated with apple rootstocks. Hortic. Res. 2022, 9, uhab013. [Google Scholar] [CrossRef] [PubMed]
- Lavely, E.K.; Chen, W.; Peterson, K.A.; Klodd, A.E.; Volder, A.; Marini, R.P.; Eissenstat, D.M. On characterizing root function in perennial horticultural crops. Am. J. Bot. 1987, 107, 1214–1224. [Google Scholar] [CrossRef]
- Mccormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Erik, A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves under-standing of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.E.; Eissenstat, D.M. Beyond the roots of young seedlings: The influence of age and order on fine root physiology. J. Plant Growth Regul. 2002, 21, 324–334. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef]
- Bouma, T.J.; Yanai, R.D.; Elkin, A.D.; Hartmond, U.; Floresalva, D.E.; Eissenstat, D.M. Estimating age-dependent costs and benefits of roots with contrasting life span: Comparing apples and oranges. New Phytol. 2001, 150, 685–695. [Google Scholar] [CrossRef]
- Feng, Z.; Yang, X.; Liang, H.; Kong, Y.; Hui, D.; Zhao, J.; Guo, E.; Fan, B. Improvements in the root morphology, physiology, and anatomy of Platycladus orientalis seedlings from air-root pruning. HortScience 2018, 53, 1750–1756. [Google Scholar] [CrossRef]
- Atucha, A.; Emmett, B.; Bauerle, T.L. Growth rate of fine root systems influences rootstock tolerance to replant disease. Plant Soil 2014, 376, 337–346. [Google Scholar] [CrossRef]
- Volder, A.; Smart, D.R.; Bloom, A.J.; Eissenstat, D.M. Rapid decline in nitrate uptake and respiration with age in fine lateral roots of grape: Implications for root efficiency and competitive effectiveness. New Phytol. 2005, 165, 493–502. [Google Scholar] [CrossRef] [PubMed]
- DeJong, T.M.; Tombesi, S.; Basile, B.; Da Silva, D. Beakbane and Thompson (1939, East Malling) had it right: Scion vigour is physiologically linked to the xylem anatomy of the rootstock. Asp. Appl. Biol. 2013, 119, 51–58. [Google Scholar]
- Tombesi, S.; Johnson, R.S.; Day, K.R.; DeJong, T.M. Interactions between rootstock, inter-stem and scion xylem vessel characteristics of peach trees growing on rootstocks with contrasting size-controlling characteristics. AoB Plants 2010, 2010, plq013. [Google Scholar] [CrossRef]
- Parry, M.S. The effects of budding height on the field performance of two apple cultivars on three rootstocks. J. Hortic. Sci. 1986, 61, 1–7. [Google Scholar] [CrossRef]
- Webster, A.D. Vigour mechanisms in dwarfing rootstocks for temperate fruit trees. Acta Hortic. 2004, 658, 29–41. [Google Scholar] [CrossRef]
- Lordan, J.; Fazio, G.; Francescatto, P.; Robinson, T.L. Effects of apple (Malus x domestica) rootstocks on scion performance and hormone concentration. Sci. Hortic. 2017, 225, 96–105. [Google Scholar] [CrossRef]
- Fazio, G.; Kviklys, A.; Grusak, M.A.; Robinson, T.L. Phenotypic diversity and QTL mapping of absorption and translocation of nutrients by apple rootstocks. Asp. Appl. Biol. 2013, 119, 37–50. [Google Scholar]
- Reighard, G.L.; Bridges, W.; Rauh, B.; Mayer, N.A. Prunus rootstocks influence peach leaf and fruit nutrient content. Acta Hortic. 2013, 984, 117–124. [Google Scholar] [CrossRef]
- Elsysy, M.A.; Mickelbart, M.V.; Hirst, P.M. Effect of fruiting and biennial bearing potential on spur quality and leaf gas exchange in apple. J. Amer. Soc. Hort. Sci. 2019, 144, 31–37. [Google Scholar] [CrossRef]
- Atkinson, D. The distribution and effectiveness of the roots of tree crops. Hort. Rev. 1980, 2, 424–490. [Google Scholar]
- Johnson, I.R. A model of the partitioning of growth between the shoots and roots of vegetative plants. Ann. Bot. 1985, 55, 421–431. [Google Scholar] [CrossRef]
- Robinson, D. Compensatory changes in the partitioning of dry matter in relation to nitrogen uptake and optimal variations in growth. Ann. Bot. 1986, 58, 841–848. [Google Scholar] [CrossRef]
- Johnson, I.R.; Thornley, J.H.M. A model of root: Shoot partitioning with optimal growth. Ann. Bot. 1987, 60, 133–142. [Google Scholar] [CrossRef]
- Van der Werf, A.; Visser, J.; Schieving, F.; Lambers, H. Evidence for Optimal Partitioning of Biomass and Nitrogen at a Range of Nitrogen Availabilities for a Fast- and Slow-Growing Species. Funct. Ecol. 1993, 7, 63–74. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| 15-August-17 | 15-November-17 | |||||
|---|---|---|---|---|---|---|
| Air-Pruning | Field-Grown | Sig. | Air-Pruning | Field-Grown | Sig. | |
| Container | Liners | Container | Liners | |||
| Rootstock shank DMC per tree (g) | ||||||
| Gala’ | 22.2 ± 1.3 | 16.4 ± 2.5 | * | 34.6 ± 1.8 | 31.5 ± 3.4 | 0.34 |
| Fuji’ | 15.7 ± 1.8 | 14.4 ± 1.5 | 0.47 | 27.5 ± 1.6 | 25.2 ± 4.5 | 0.34 |
| Honeycrisp’ | 20.9 ± 1.8 | 20.4 ± 1.9 | 0.75 | 32.0 ± 1.8 | 27.7 ± 3.2 | 0.11 |
| Cultivar | *** | ** | ||||
| System | * | * | ||||
| Cultivar*system | 0.1 | 0.91 | ||||
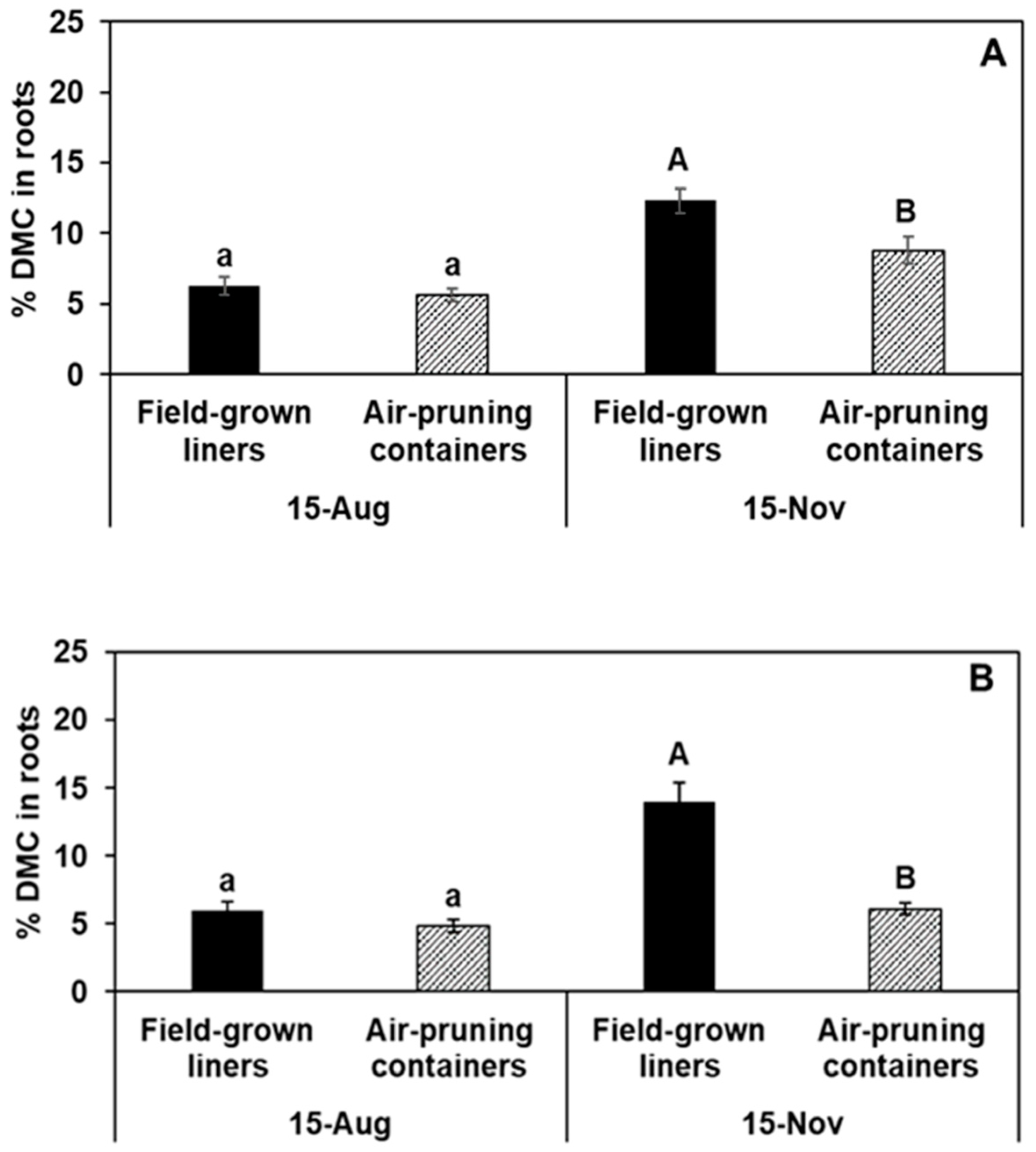
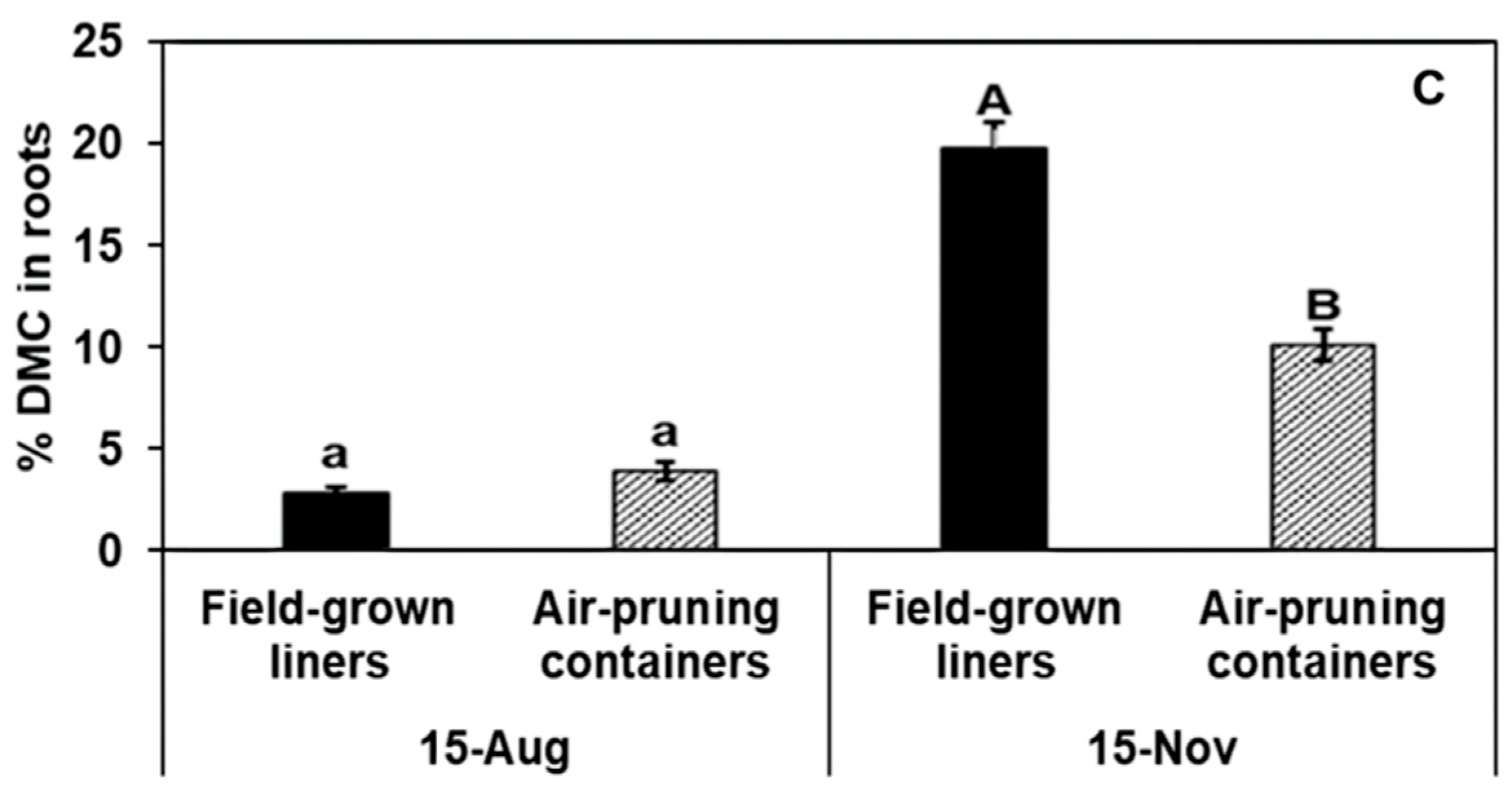
| Total root DMC per tree (g) | ||||||
| Gala’ | 0.92 ± 0.11 | 0.78 ± 0.09 | 0.19 | 3.2 ± 0.34 | 4.2 ± 0.7 | 0.07 |
| Fuji’ | 0.62 ± 0.12 | 0.52 ± 0.12 | 0.44 | 2.0 ± 0.17 | 3.2 ± 0.19 | ** |
| Honeycrisp’ | 0.62 ± 0.08 | 0.57 ± 0.06 | 0.55 | 3.0 ± 0.37 | 3.9 ± 0.58 | 0.07 |
| Cultivar | *** | * | ||||
| System | 0.11 | *** | ||||
| Cultivar*system | 0.84 | 0.87 | ||||
| Total root volume per tree (cm3) | ||||||
| Gala’ | 7.0 ± 0.91 | 2.8 ± 0.21 | *** | 8.9 ± 0.71 | 5.9 ± 0.63 | ** |
| Fuji’ | 4.1 ± 0.82 | 2.1 ± 0.41 | ** | 7.3 ± 0.38 | 6.2 ± 0.18 | 0.16 |
| Honeycrisp’ | 3.7 ± 0.26 | 2.4 ± 0.29 | ** | 8.8 ± 0.67 | 5.6 ± 0.78 | ** |
| Cultivar | *** | 0.71 | ||||
| System | *** | *** | ||||
| Cultivar*system | ** | 0.25 | ||||
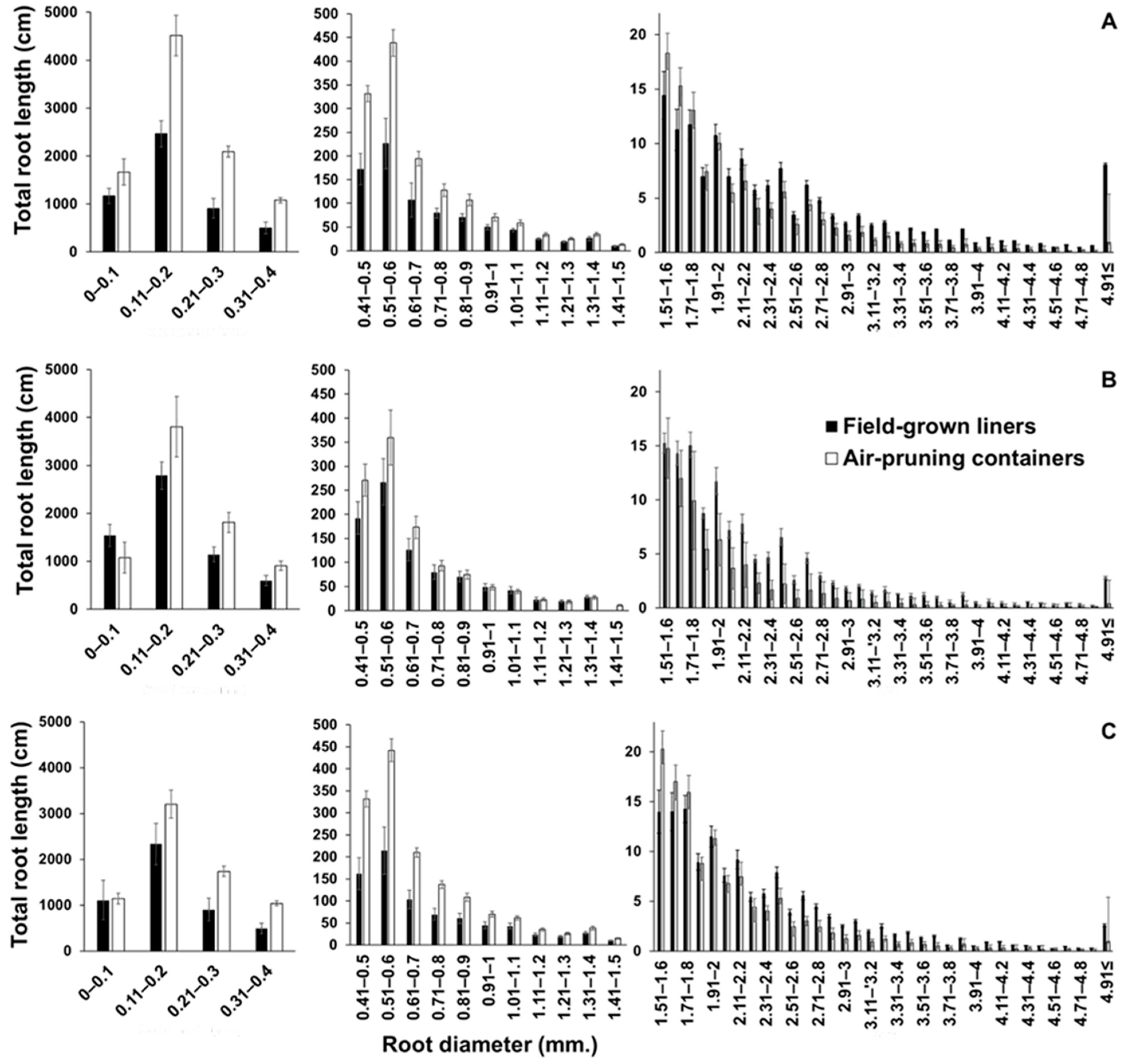
| Total root length per tree (cm) | ||||||
| Gala’ | 7475.9 ± 926 | 2391.7 ± 659 | *** | 19888.9 ± 1285 | 5617.9 ± 1748 | *** |
| Fuji’ | 3522.1 ± 699 | 1729.1 ± 354 | ** | 16932.1 ± 788 | 7335.9 ± 452 | *** |
| Honeycrisp’ | 4321.7 ± 485 | 2737.9 ± 231 | ** | 15836.7 ± 777 | 5748.2 ± 598 | *** |
| Cultivar | *** | 0.14 | ||||
| System | *** | *** | ||||
| Cultivar*system | *** | 0.08 | ||||
| Total length of roots ≤ 0.4 mm per tree (cm) | ||||||
| Gala’ | 6147.4 ± 924 | 1942.7 ± 662 | *** | 9181.7 ± 1287 | 4682.0 ± 1738 | *** |
| Fuji’ | 2751.0 ± 696 | 1392.5 ± 348 | ** | 7829.7 ± 732 | 6271.6 ± 1257 | 0.09 |
| Honeycrisp’ | 3547.5 ± 486 | 2307.2 ± 430 | 0.10 | 7120.4 ± 779 | 4830.7 ± 586 | ** |
| Cultivar | *** | 0.15 | ||||
| System | *** | *** | ||||
| Cultivar*system | *** | 0.06 | ||||
| Total root tips per tree (no.) | ||||||
| Gala’ | 39343.3 ± 7650 | 10517.6 ± 2551 | *** | 70017.2 ± 13665 | 24991.9 ± 1136 | *** |
| Fuji’ | 11341.0 ± 2223 | 9412.6 ± 1638 | 0.47 | 51392.0 ± 9276 | 44089.4 ± 6759 | 0.41 |
| Honeycrisp’ | 16394.9 ± 4986 | 14609.0 ± 635 | 0.53 | 50033.7 ± 6546 | 30337.3 ± 3297 | *** |
| Cultivar | *** | 0.40 | ||||
| System | *** | *** | ||||
| Cultivar*system | *** | ** | ||||
| 15-August-17 | 15-November-17 | |||||
|---|---|---|---|---|---|---|
| Air-Pruning | Field-Grown | Sig. | Air-Pruning | Field-Grown | Sig. | |
| Container | Liners | Container | Liners | |||
| Scion (wood) DMC per tree (g) | ||||||
| Gala’ | 5.6 ± 0.49 | 5.4 ± 0.55 | 0.67 | 17.0 ± 1.2 | 15.1 ± 2.8 | 0.10 |
| Fuji’ | 5.3 ± 0.9 | 4.0 ± 1.1 | 0.06 | 16.8 ± 1.7 | 10.1 ± 2.1 | *** |
| Honeycrisp’ | 5.5 ± 0.78 | 6.0 ± 0.54 | 0.37 | 11.9 ± 0.4 | 5.6 ± 1.3 | *** |
| Cultivar | * | *** | ||||
| System | 0.36 | *** | ||||
| Cultivar*system | 0.21 | 0.08 | ||||
| Scion total length per tree (cm) | ||||||
| Gala’ | 61.2 ± 2.6 | 42.5 ± 4.4 | *** | 85.0 ± 1.5 | 83.8 ± 5.2 | 0.44 |
| Fuji’ | 49.6 ± 6 | 27.7 ± 3.4 | *** | 88.3 ± 0.56 | 75.0 ± 9.8 | * |
| Honeycrisp’ | 51.8 ± 4.4 | 44.8 ± 2.1 | * | 62.5 ± 3.1 | 41.3 ± 4.6 | *** |
| Cultivar | *** | *** | ||||
| System | *** | *** | ||||
| Cultivar*system | * | * | ||||
| Canopy leaf DMC per tree (g) | ||||||
| Gala’ | 9.8 ± 0.4 | 7.1 ± 0.67 | *** | 16.5 ± 1.2 | 16.6 ± 2. | 0.66 |
| Fuji’ | 6.1 ± 0.98 | 4.5 ± 0.25 | * | 12.9 ± 0.5 | 10.4 ± 1.4 | ** |
| Honeycrisp’ | 9.4 ± 0.66 | 12.3 ± 0.35 | *** | 14.7 ± 0.4 | 10.9 ± 1.7 | ** |
| Cultivar | *** | *** | ||||
| System | 0.11 | *** | ||||
| Cultivar*system | *** | 0.15 | ||||
| Canopy leaf area per tree (cm2) | ||||||
| Gala’ | 878.7 ± 19.2 | 575.3 ± 72.5 | *** | 1227.6 ± 77 | 1161.9 ± 87 | 0.32 |
| Fuji’ | 602.3 ± 87.9 | 347.8 ± 41.3 | *** | 1027.0 ± 28 | 827.5 ± 129 | ** |
| Honeycrisp’ | 843.8 ± 40.2 | 865.8 ± 9.9 | 0.6 | 951.6 ± 24 | 617.4 ± 98 | *** |
| Cultivar | *** | *** | ||||
| System | *** | *** | ||||
| Cultivar*system | 0.18 | 0.06 | ||||
| Leaves per tree (no) | ||||||
| Gala’ | 45.7 ± 3.7 | 40.9 ± 4.2 | 0.36 | 45.8 ± 2.9 | 48.3 ± 2.3 | 0.42 |
| Fuji’ | 37.9 ± 2 | 35.0 ± 2.5 | 0.36 | 52.0 ± 1.1 | 50.3 ± 1.9 | 0.21 |
| Honeycrisp’ | 38.1 ± 1.4 | 41.6 ± 2.1 | 0.18 | 41.0 ± 1.2 | 34.2 ± 3 | * |
| Cultivar | * | *** | ||||
| System | 0.51 | 0.15 | ||||
| Cultivar*system | 0.30 | 0.07 | ||||
| Individual leaf DMC (g) | ||||||
| Gala’ | 0.23 ± 0.02 | 0.18 ± 0.02 | * | 0.38 ± 0.03 | 0.35 ± 0.05 | 0.27 |
| Fuji’ | 0.16 ± 0.02 | 0.14 ± 0.02 | 0.35 | 0.25 ± 0.01 | 0.20 ± 0.03 | * |
| Honeycrisp’ | 0.25 ± 0.01 | 0.30 ± 0.01 | ** | 0.36 ± 0.01 | 0.32 ± 0.03 | * |
| Cultivar | *** | *** | ||||
| System | 0.35 | ** | ||||
| Cultivar*system | ** | 1.0 | ||||
| Individual leaf area (cm2) | ||||||
| Gala’ | 20.2 ± 1.6 | 14.3 ± 1.1 | *** | 27.8 ± 2.1 | 24.5 ± 2.5 | 0.09 |
| Fuji’ | 16.1 ± 2.2 | 10.4 ± 0.7 | *** | 19.8 ± 0.9 | 16.2 ± 2.7 | * |
| Honeycrisp’ | 22.3 ± 0.75 | 21.0 ± 0.9 | 0.4 | 23.4 ± 0.7 | 17.9 ± 1.6 | *** |
| Cultivar | *** | *** | ||||
| System | *** | *** | ||||
| Cultivar*system | * | 0.06 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsysy, M.; Einhorn, T.C. Air-Pruning Containers Modify Root and Scion Growth and Alter Resource Allocation of Bench-Grafted Apple Plants. Horticulturae 2022, 8, 797. https://doi.org/10.3390/horticulturae8090797
Elsysy M, Einhorn TC. Air-Pruning Containers Modify Root and Scion Growth and Alter Resource Allocation of Bench-Grafted Apple Plants. Horticulturae. 2022; 8(9):797. https://doi.org/10.3390/horticulturae8090797
Chicago/Turabian StyleElsysy, Mokhles, and Todd C. Einhorn. 2022. "Air-Pruning Containers Modify Root and Scion Growth and Alter Resource Allocation of Bench-Grafted Apple Plants" Horticulturae 8, no. 9: 797. https://doi.org/10.3390/horticulturae8090797
APA StyleElsysy, M., & Einhorn, T. C. (2022). Air-Pruning Containers Modify Root and Scion Growth and Alter Resource Allocation of Bench-Grafted Apple Plants. Horticulturae, 8(9), 797. https://doi.org/10.3390/horticulturae8090797