
Combined Nano Silver, α-Aminoisobutyric Acid, and 1-Methylcyclopropene Treatment Delays the Senescence of Cut Roses with Different Ethylene Sensitivities



Abstract
:1. Introduction
2. Material and Method
2.1. Plant Materials
2.2. NS, AIB, and 1-MCP Treatments
2.3. Ethylene Treatment
2.4. Measurements
2.4.1. Anti-Bacterial Activity
2.4.2. Flower Diameter (FD) and Water Relations
2.4.3. Maximal PSII Quantum Yield (Fv/Fm), Soluble Solids Content (SSC), and Chlorophyll Content
2.4.4. Vase life (VL) and Senescence Symptoms
2.5. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.6. Experiment Design and Data Analysis
3. Results
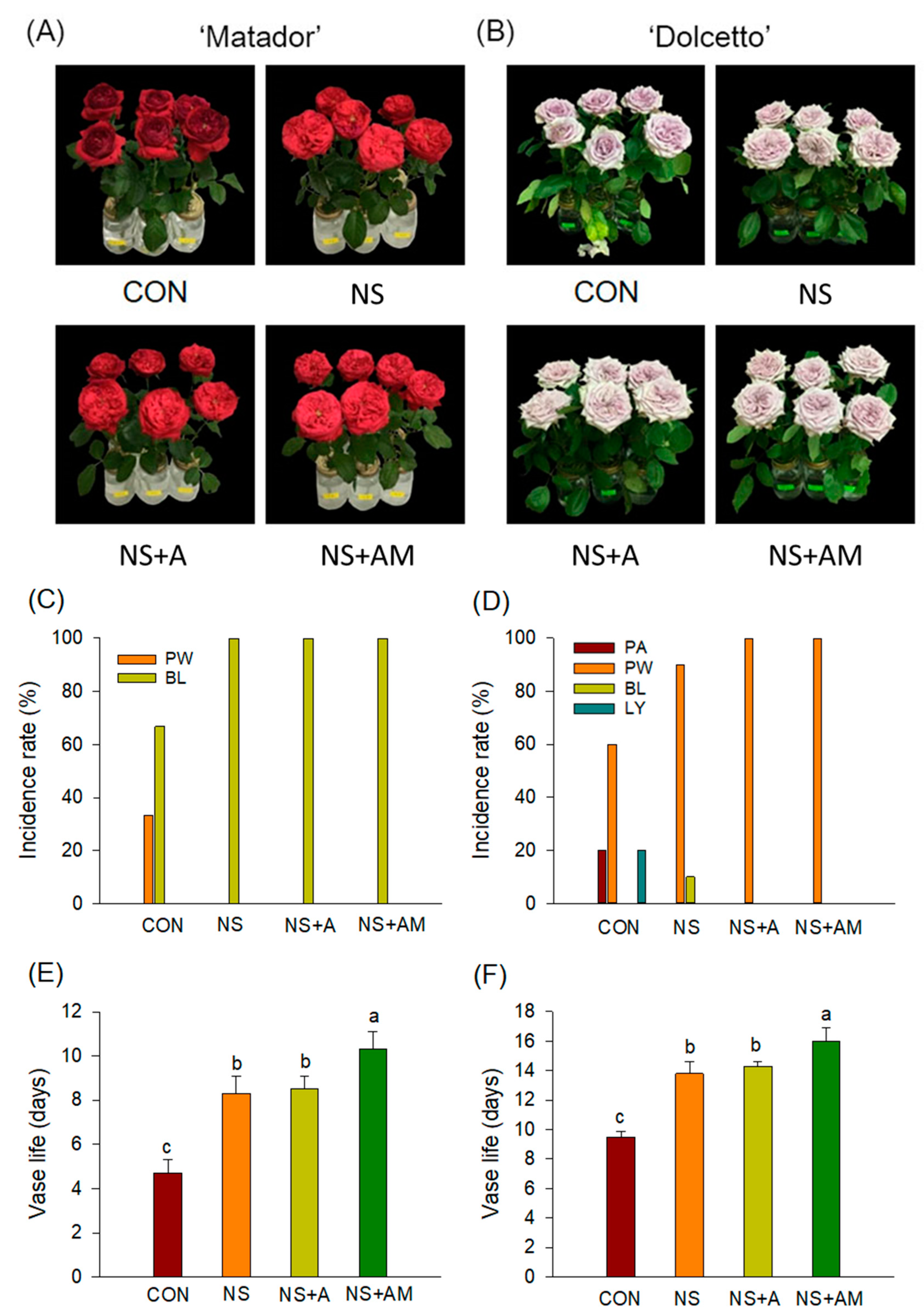
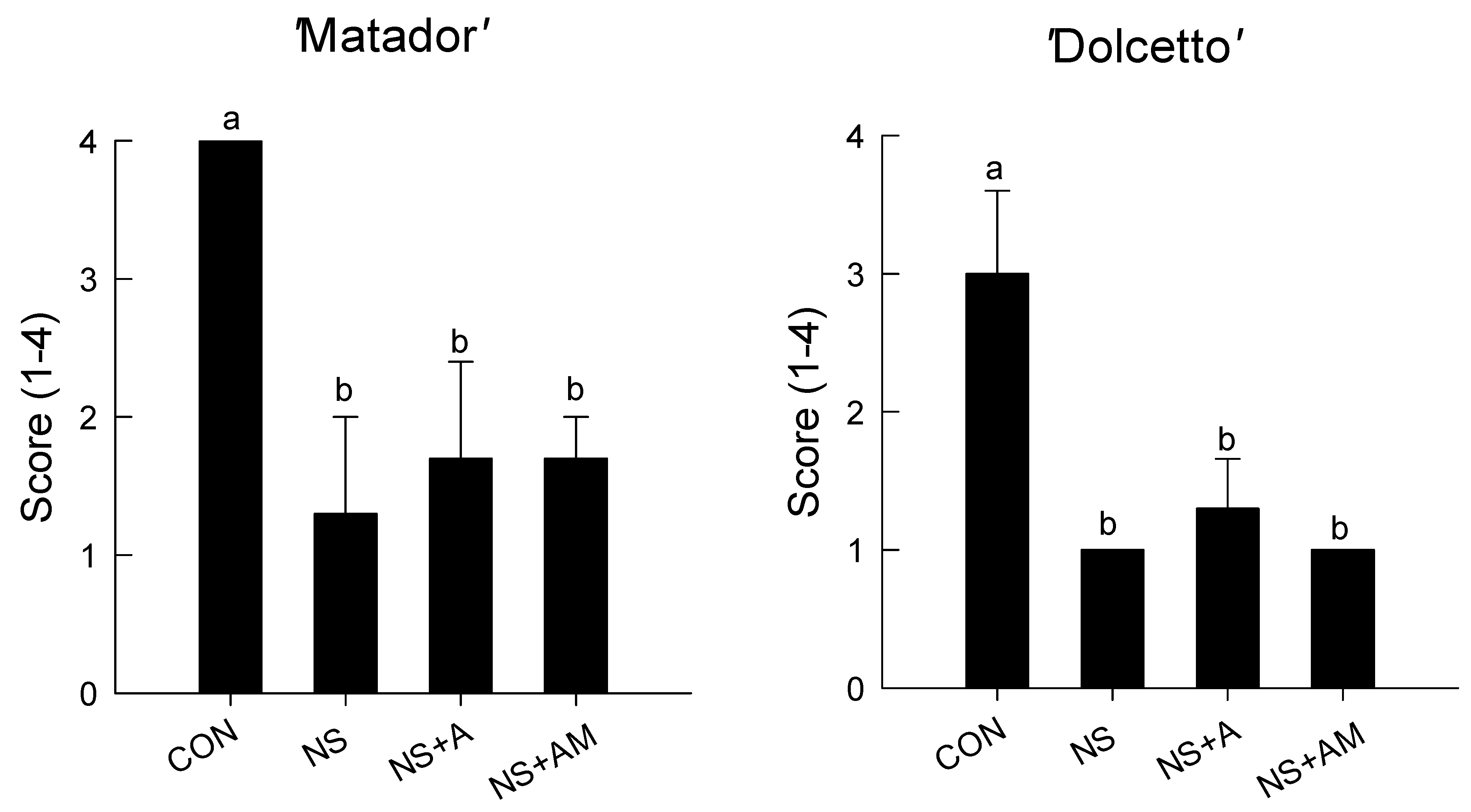
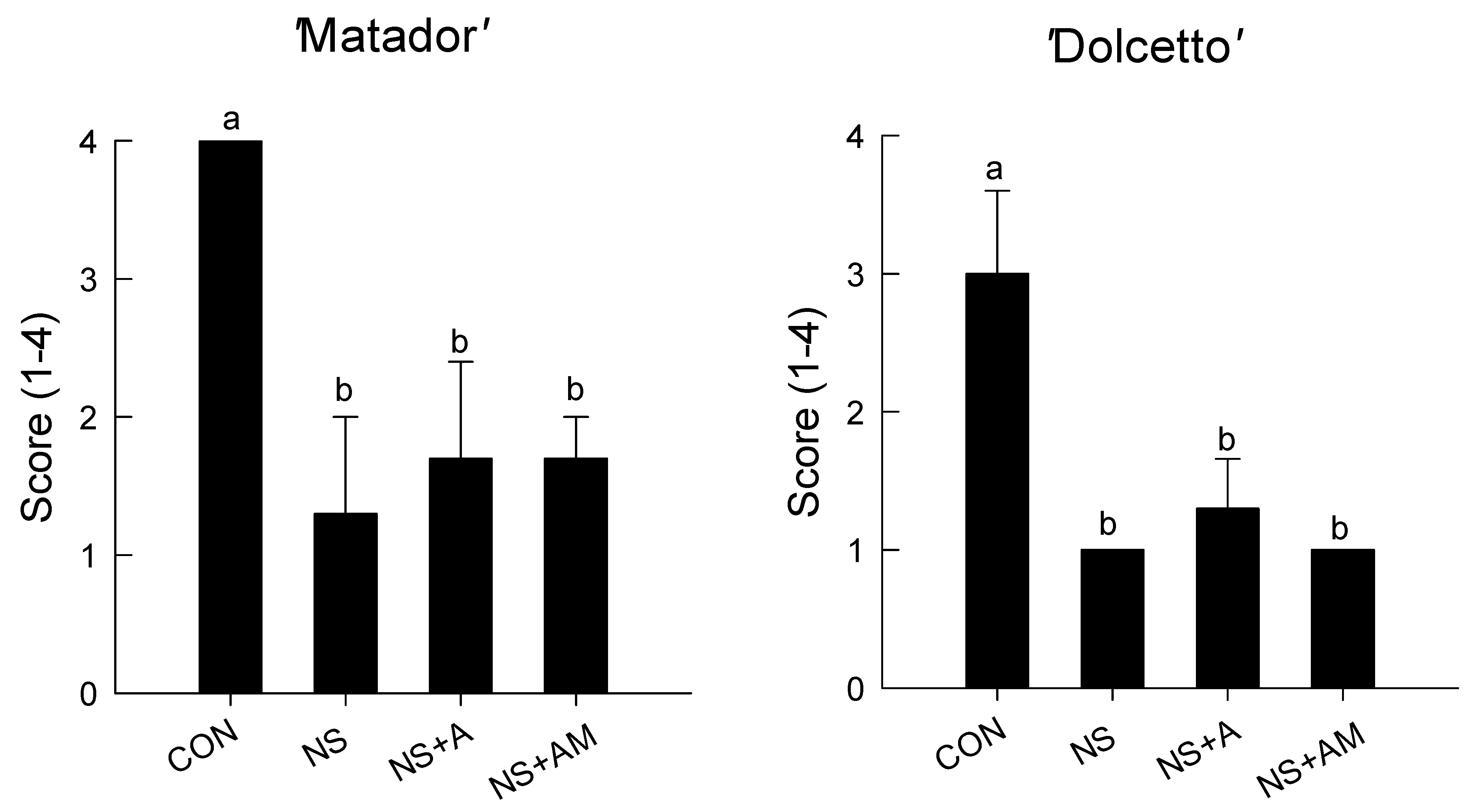
3.1. Senescence Symptoms and Longevity of Cut Roses
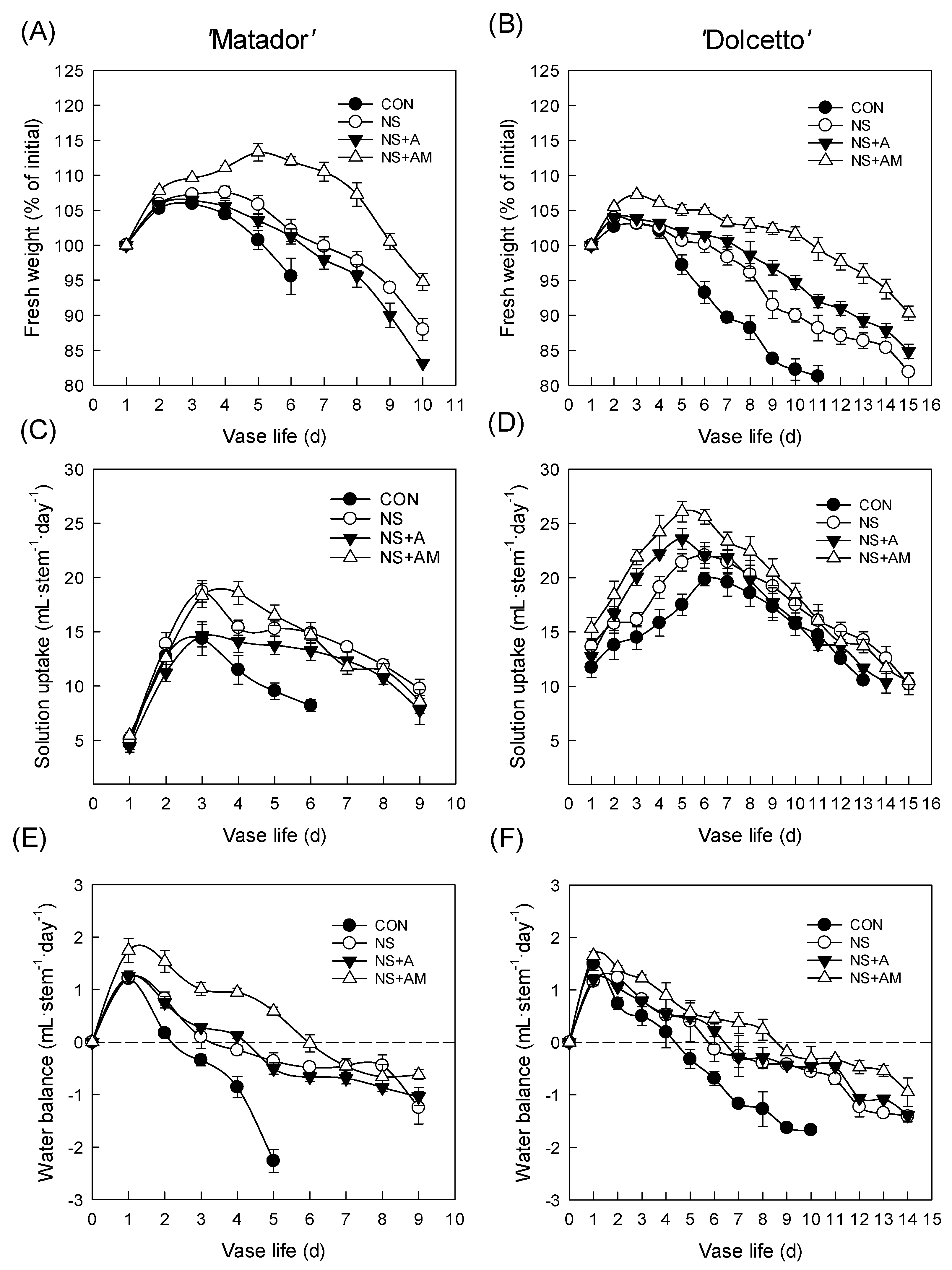
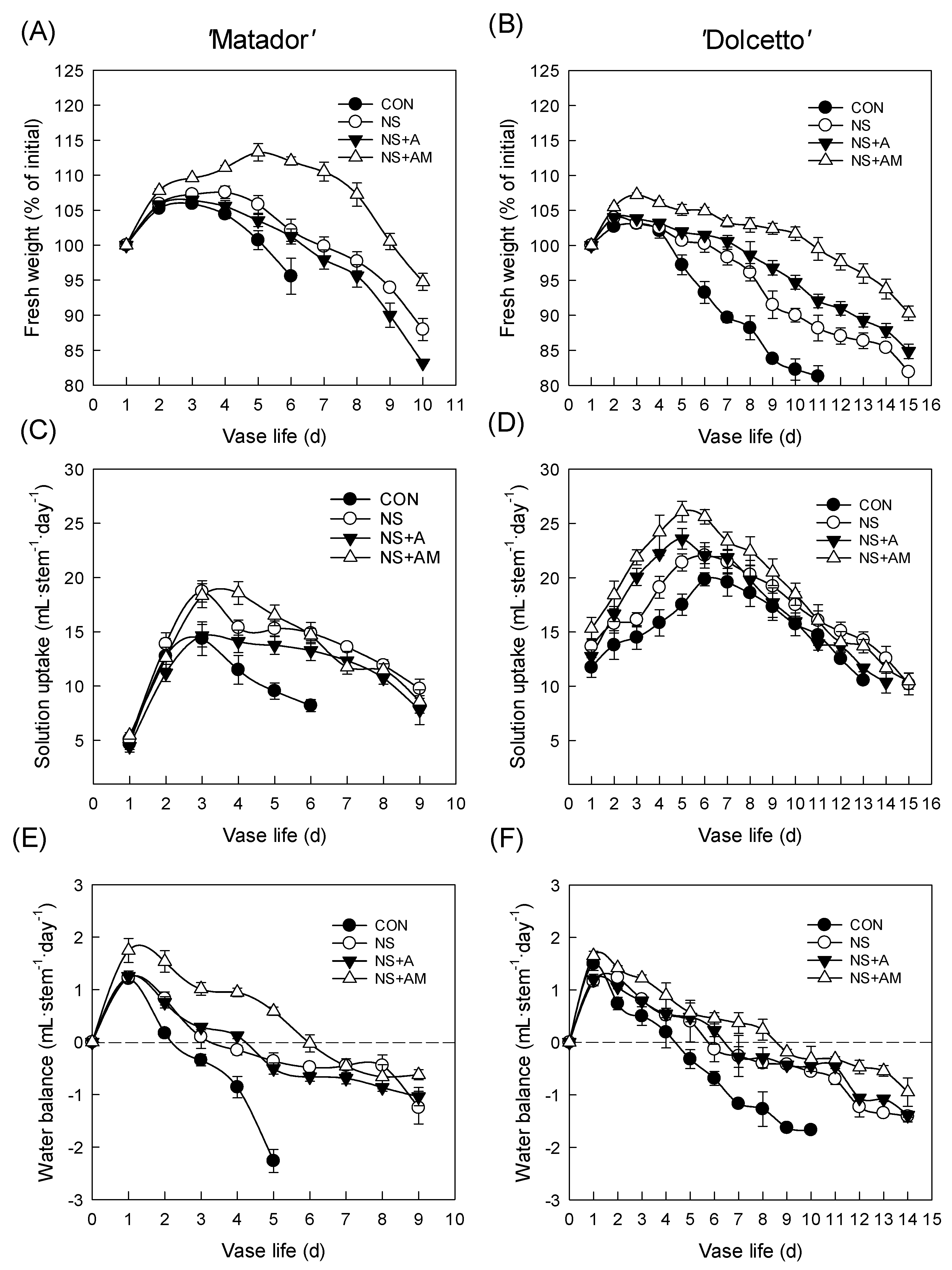
3.2. Antibacterial Activity of the Treatments and Water Relations of Cut Flowers
3.3. FD and SSC of Cut Roses
3.4. Fv/Fm and Chlorophyll Content (SPAD)
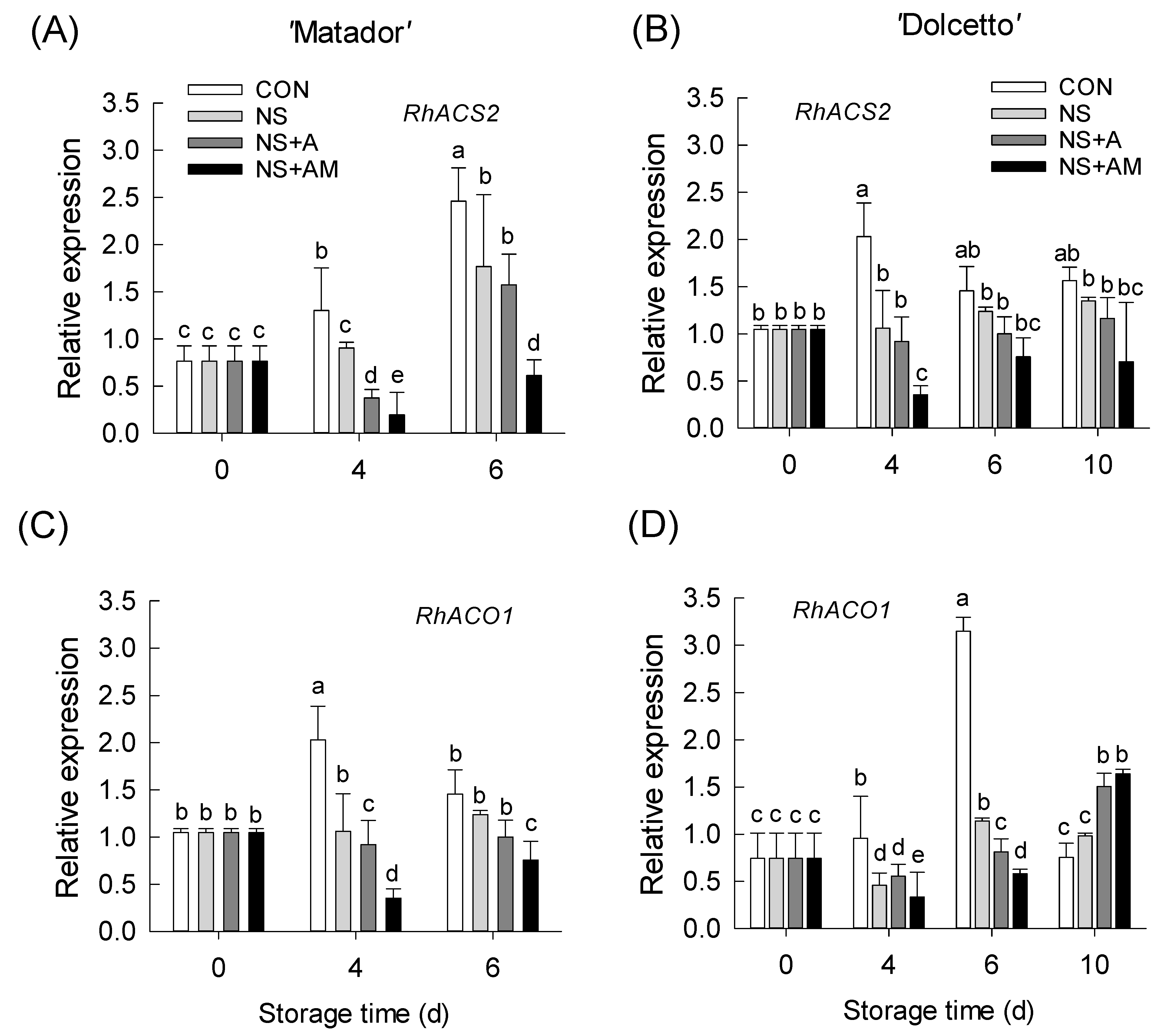
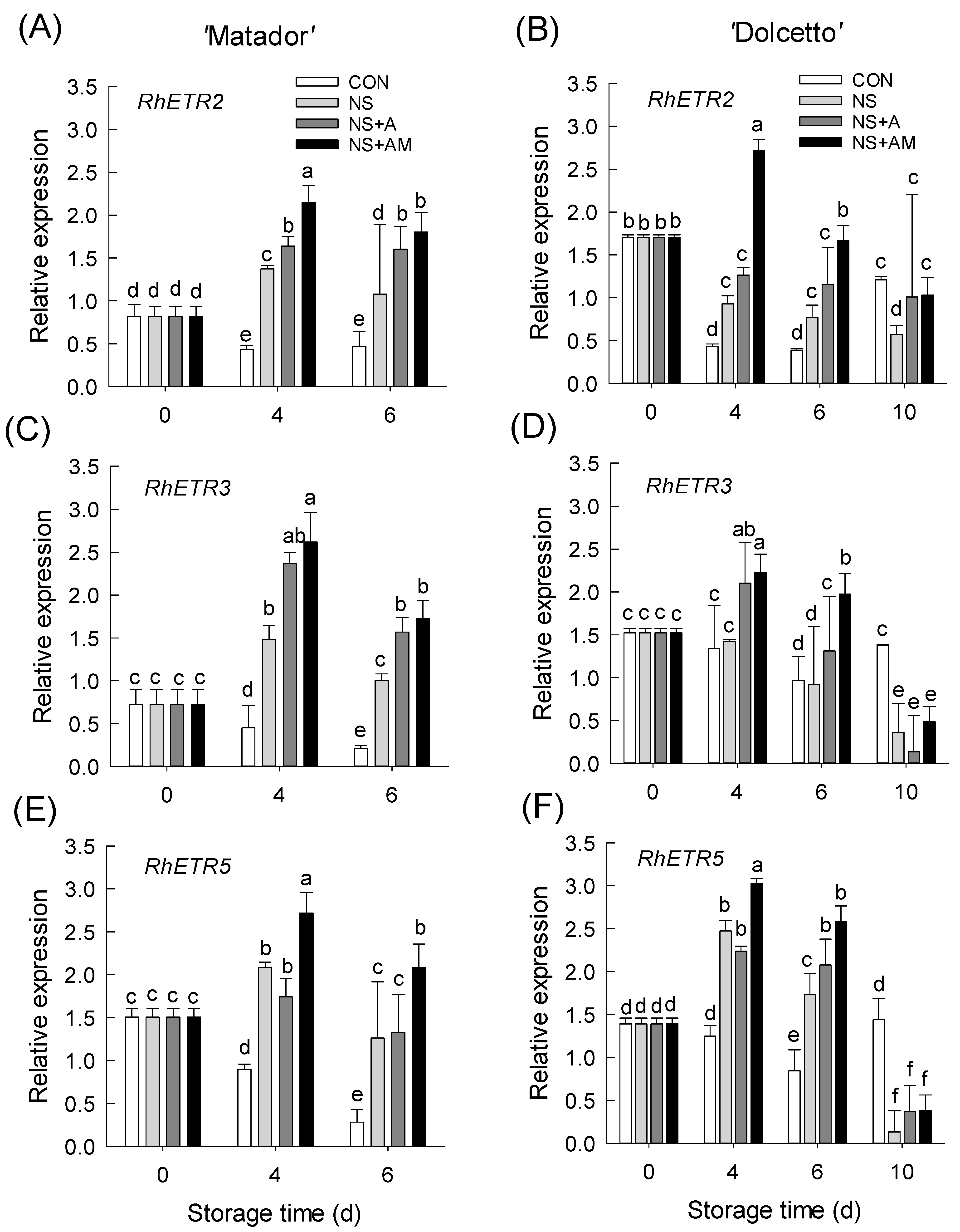
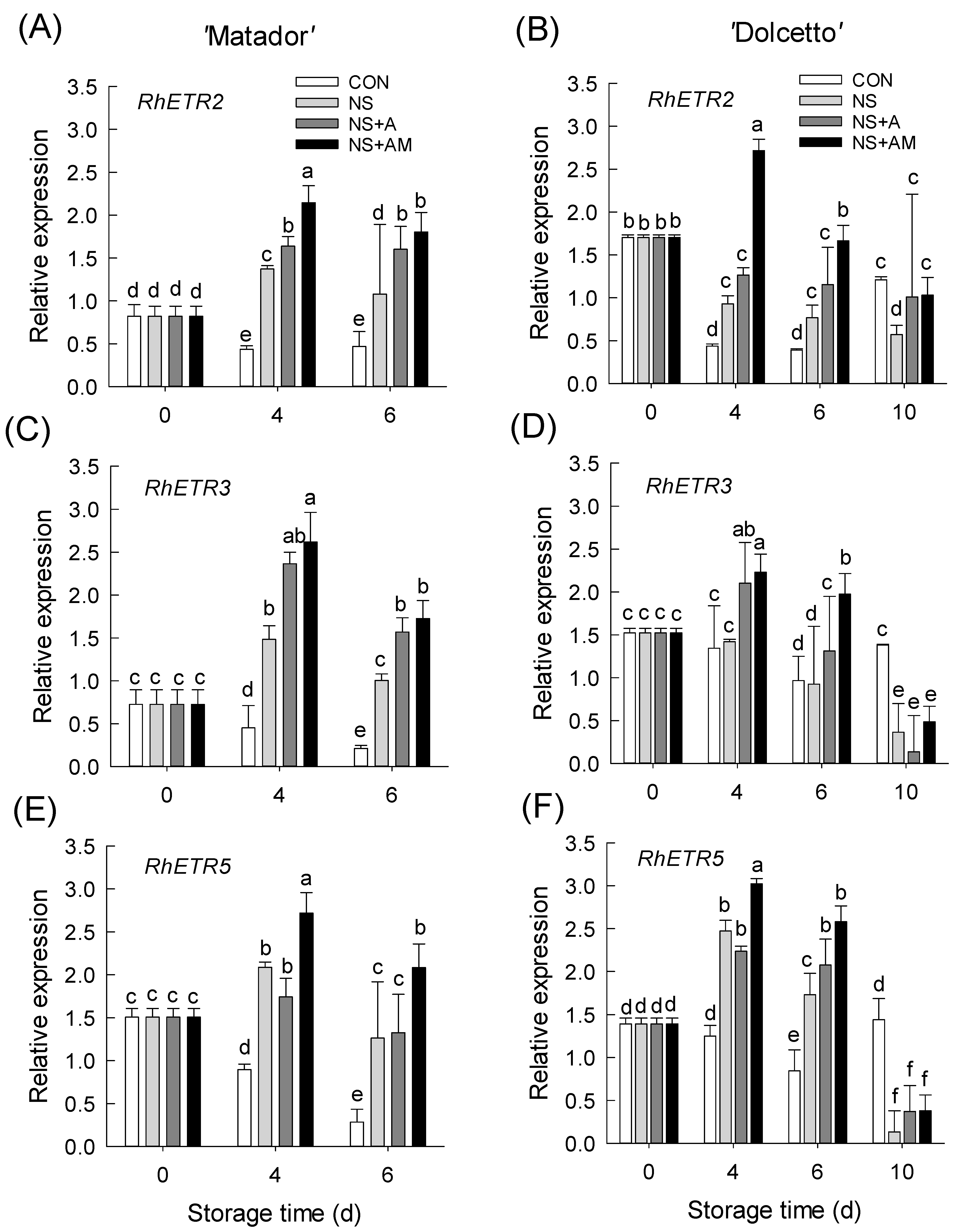
3.5. Changes in mRNA Levels of Ethylene Biosynthesis and Receptor Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- In, B.-C.; Ha, S.T.T.; Lee, Y.S.; Lim, J.H. Relationships between the longevity, water relations, ethylene sensitivity, and gene expression of cut roses. Postharvest Biol. Technol. 2017, 131, 74–83. [Google Scholar] [CrossRef]
- Macnish, A.J.; Leonard, R.T.; Borda, A.M.; Nell, T.A. Genotypic variation in the postharvest performance and ethylene sensitivity of cut rose flowers. HortScience Horts 2010, 45, 790–796. [Google Scholar] [CrossRef] [Green Version]
- Van Doorn, W.G.; Witte, Y.d.; Harkema, H. Effect of high numbers of exogenous bacteria on the water relations and longevity of cut carnation flowers. Postharvest Biol. Technol. 1995, 6, 111–119. [Google Scholar] [CrossRef]
- Van Doorn, W.G. Water relations of cut flowers: An update. Hortic. Rev. 2012, 40, 55–106. [Google Scholar]
- Müller, R.; Andersen, A.S.; Serek, M. Differences in display life of miniature potted roses (Rosa hybrida L.). Sci. Hortic. 1998, 76, 59–71. [Google Scholar] [CrossRef]
- Gong, B.; Huang, S.; Ye, N.; Yuan, X.; Ma, H. Pre-harvest ethylene control affects vase life of cut rose ‘Carola’ by regulating energy metabolism and antioxidant enzyme activity. Hortic. Environ. Biotechnol. 2018, 59, 835–845. [Google Scholar] [CrossRef]
- Chamani, E.; Khalighi, A.; Joyce, D.C.; Irving, D.E.; Zamani, Z.; Mostofi, Y.; Kafi, M. Ethylene and anti-ethylene treatment effects on cut ‘First Red’ rose. J. Appl. Hortic. Lucknow 2005, 7, 3–7. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.-B.; Zhang, M.-L.; Yu, J.-H. Role of nitric oxide in delaying senescence of cut rose flowers and its interaction with ethylene. Sci. Hortic. 2013, 155, 30–38. [Google Scholar] [CrossRef]
- Ha, S.T.T.; Lim, J.-H.; In, B.-C. Simultaneous inhibition of ethylene biosynthesis and binding using AVG and 1-MCP in two rose cultivars with different sensitivities to ethylene. J. Plant Growth Regul. 2020, 39, 553–563. [Google Scholar] [CrossRef]
- Ha, S.T.T.; In, B.C.; Choi, H.W.; Jong, Y.O.; Lim, J.H. Assessment of pretreatment solutions for improving the vase life and postharvest quality of cut roses (Rosa hybrida L. ‘Jinny’). Flower Res. J. 2017, 25, 101–109. [Google Scholar] [CrossRef]
- Satoh, S.; Esashi, Y. α-Aminoisobutyric acid: A probable competitive inhibitor of conversion of 1-aminocyclopropane-1-carboxylic acid to ethylene. Plant Cell Physiol. 1980, 21, 939–949. [Google Scholar] [CrossRef]
- Satoh, S.; Esashi, Y. Effects of Ñ aminoisobutyric acid and d and l amino acids on ethylene production and content of 1 aminocyclopropane Q carboxylic acid in cotyledonary segments of cocklebur seeds. Physiol. Plant. 1982, 54, 147–152. [Google Scholar] [CrossRef]
- Ha, S.T.T.; Lim, J.H.; In, B.C. Extension of the vase life of cut roses by both improving water relations and repressing ethylene responses. Korean J. Hortic. Sci. Technol. 2019, 37, 65–77. [Google Scholar]
- Serek, M.; Sisler, E.C.; Reid, M.S. Effects of 1-MCP on the vase life and ethylene response of cut flowers. Plant Growth Regul. 1995, 16, 93–97. [Google Scholar] [CrossRef]
- Golding, J.B.; Singh, S.P. Use of 1-MCP in the storage life extension of fruit. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Lok, C.-N.; Ho, C.-M.; Chen, R.; He, Q.-Y.; Yu, W.-Y.; Sun, H.; Tam, P.K.-H.; Chiu, J.-F.; Che, C.-M. Proteomic analysis of the mode of antibacterial action of silver nanoparticles. J. Proteome Res. 2006, 5, 916–924. [Google Scholar] [CrossRef]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2009, 27, 76–83. [Google Scholar] [CrossRef]
- Liu, J.; Lai, L.; Liu, H.; Li, H.; Yu, G.; Sun, Y.; He, S. Nano-silver treatment reduces bacterial proliferation and stem bending in cut gerbera flowers: An in vitro and in vivo evaluation. Postharvest Biol. Technol. 2021, 180, 111595. [Google Scholar] [CrossRef]
- Ha, S.T.T.; Kim, Y.-T.; Jeon, Y.H.; Choi, H.W.; In, B.-C. Regulation of Botrytis cinerea infection and gene expression in cut roses by using nano silver and salicylic acid. Plants 2021, 10, 1241. [Google Scholar] [CrossRef]
- Skutnik, E.; Jędrzejuk, A.; Rabiza-Świder, J.; Rochala-Wojciechowska, J.; Latkowska, M.; Łukaszewska, A. Nanosilver as a novel biocide for control of senescence in garden cosmos. Sci. Rep. 2020, 10, 10274. [Google Scholar] [CrossRef]
- Park, D.Y.; Naing, A.H.; Ai, T.N.; Han, J.-S.; Kang, I.-K.; Kim, C.K. Synergistic effect of nano-sliver with sucrose on extending vase life of the carnation cv. Edun. Front. Plant Sci. 2017, 8, 1601. [Google Scholar] [CrossRef] [Green Version]
- In, B.-C.; Lim, J.H. Potential vase life of cut roses: Seasonal variation and relationships with growth conditions, phenotypes, and gene expressions. Postharvest Biol. Technol. 2018, 135, 93–103. [Google Scholar] [CrossRef]
- VBN. Evaluation Cards for Rosa; FloraHollandAalsmeer: Aalsmeer, The Netherlands, 2014. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Halevy, A.H.; Mayak, S. Senescence and postharvest physiology of cut flowers—Part 2. Hortic. Rev. 1981, 3, 59–143. [Google Scholar]
- Paulin, A. Influence of exogenous sugars on the evolution of some senescence parameters of petals. Acta Hortic. 1986, 181, 183–194. [Google Scholar] [CrossRef]
- Shibuya, K.; Yoshioka, T.; Hashiba, T.; Satoh, S. Role of the gynoecium in natural senescence. Influence of exogenous sugars on the evolution of some senescence parameters of petalsof carnation (Dianthus caryophyllus L.) flowers. J. Exp. Bot. 2000, 51, 2067–2073. [Google Scholar] [CrossRef] [Green Version]
- In, B.-C.; Binder, B.M.; Falbel, T.G.; Patterson, S.E. Analysis of gene expression during the transition to climacteric phase in carnation flowers (Dianthus caryophyllus L.). J. Exp. Bot. 2013, 64, 4923–4937. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L. Ethylene biosynthetic genes are differentially regulated by ethylene and ACC in carnation styles. Plant Growth Regul. 2003, 40, 129–138. [Google Scholar] [CrossRef]
- Ha, S.T.T.; In, B.C. Ethylene biosynthesis, signaling, and regulation in roses. Flower Res. J. 2021, 29, 1–10. [Google Scholar]
- Rodríguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef]
- Syu, Y.Y.; Hung, J.H.; Chen, J.C.; Chuang, H.W. Impacts of size and shape of silver nanoparticles on Arabidopsis plant growth and gene expression. Plant Physiol. Biochem. PPB 2014, 83, 57–64. [Google Scholar] [CrossRef]
- Yang, Y.; Qin, N.; Huang, J.; Guo, A.; Kang, X.; Li, S.; Xing, G. Dynamic changes of pectin epitopes and daylily tepals during flower opening and senescence of Hemerocallis citrina. Sci. Hortic. 2021, 288, 110367. [Google Scholar] [CrossRef]
- Secchi, F.; Gillbert, M.E.; Zwieniecki, M.A. Transcriptome response to embolism formation in stems of Populus trichocarpa provides insight into signaling and the biology and refilling. Plant Physiol. 2011, 157, 1419–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lear, B.; Casey, M.; Stead, A.D.; Rogers, H.J. Peduncle necking in Rosa hybrida induces stress-related transcription factors, upregulates galactose metabolism, and downregulates phenylpropanoid biosynthesis genes. Front. Plant Sci 2022, 13, 874590. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Forward Primer | Reverse Primer | Size |
|---|---|---|---|
| RhACS2 (AY803737.1) | 5′-GCGAACAGGGGTACAACTTC-3′ | 5′-GGGTTTGAGGGGTTGGTAAT-3′ | 147 |
| RhACO1 (AF441282.1) | 5′-CGTTCTACAACCCAGGCAAT-3′ | 5′-TTGAGGCCTGCATAGAGCTT-3′ | 130 |
| RhETR2 (AF127220.1) | 5′-CTGCGTTAGAGCAGCAACTG-3′ | 5′-GGAATTCGGCGATATCTTCA-3′ | 131 |
| RhETR3 (AY953392.1) | 5′-CCATGAGTTGAAAGGGAGGA-3′ | 5′-GGCTCACCAAAATCACCACT-3′ | 156 |
| RhETR5 (AF441283.1) | 5′-TGTGTGGAGCGACACATCTT-3′ | 5′-TGAGGGCAGTAGCACATGAC-3′ | 120 |
| RhACT1 (KC514918.1) | 5′-GTTCCCAGGAATCGCTGATA-3′ | 5′-ATCCTCCGATCCAAACACTG-3′ | 116 |
| Cultivar | Treatment | MFD (% of Initial) | SSC (%) | Fv/Fm | Chlorophyll Content (SPAD Value) |
|---|---|---|---|---|---|
| ‘Matador’ | CON | 114.8 ± 4.4 c ˣ | 0.22 ± 0.00 c | 0.62 ± 0.03 b | 48.7 ± 0.2 b |
| NS | 116.0 ± 3.3 b | 0.22 ± 0.00 c | 0.72 ± 0.02 ab | 49.9 ± 0.3 b | |
| NS+A | 118.6 ± 6.8 b | 0.30 ± 0.03 b | 0.74 ± 0.01 ab | 49.8 ± 0.4 b | |
| NS+AM | 129.7 ± 2.5 a | 0.33 ± 0.01 a | 0.75 ± 0.01 a | 51.5 ± 0.2 a | |
| ‘Dolcetto’ | CON | 160.2 ± 4.6 c | 0.10 ± 0.00 c | 0.58 ± 0.06 c | 35.5 ± 5.8 c |
| NS | 162.7 ± 6.7 c | 0.27 ± 0.06 b | 0.77 ± 0.06 ab | 48.4 ± 0.4 b | |
| NS+A | 169.9 ± 6.4 b | 0.27 ± 0.03 b | 0.76 ± 0.01 b | 48.3 ± 0.4 b | |
| NS+AM | 178.8 ± 5.2 a | 0.32 ± 0.06 a | 0.78 ± 0.01 a | 53.1 ± 4.7 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, S.T.T.; In, B.-C. Combined Nano Silver, α-Aminoisobutyric Acid, and 1-Methylcyclopropene Treatment Delays the Senescence of Cut Roses with Different Ethylene Sensitivities. Horticulturae 2022, 8, 482. https://doi.org/10.3390/horticulturae8060482
Ha STT, In B-C. Combined Nano Silver, α-Aminoisobutyric Acid, and 1-Methylcyclopropene Treatment Delays the Senescence of Cut Roses with Different Ethylene Sensitivities. Horticulturae. 2022; 8(6):482. https://doi.org/10.3390/horticulturae8060482
Chicago/Turabian StyleHa, Suong Tuyet Thi, and Byung-Chun In. 2022. "Combined Nano Silver, α-Aminoisobutyric Acid, and 1-Methylcyclopropene Treatment Delays the Senescence of Cut Roses with Different Ethylene Sensitivities" Horticulturae 8, no. 6: 482. https://doi.org/10.3390/horticulturae8060482
APA StyleHa, S. T. T., & In, B.-C. (2022). Combined Nano Silver, α-Aminoisobutyric Acid, and 1-Methylcyclopropene Treatment Delays the Senescence of Cut Roses with Different Ethylene Sensitivities. Horticulturae, 8(6), 482. https://doi.org/10.3390/horticulturae8060482