
Cold Treatment Modulates Changes in Primary Metabolites and Flowering of Cut Flower Tulip Hybrids

 ,
, 

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture
2.2. Plant Growth and Flowering
2.3. Macronutrients Content in Leaves
2.4. Metabolic Profile
2.4.1. Starch and Soluble Carbohydrate Analysis
2.4.2. Soluble Proteins: Free Amino Acid Analysis
2.4.3. Polyphenols Analysis
2.4.4. Chlorophylls and Carotenoids Analysis
2.5. Statistical Analysis
3. Results
3.1. Plant Growth and Flowering
3.2. Analyzed Metabolites
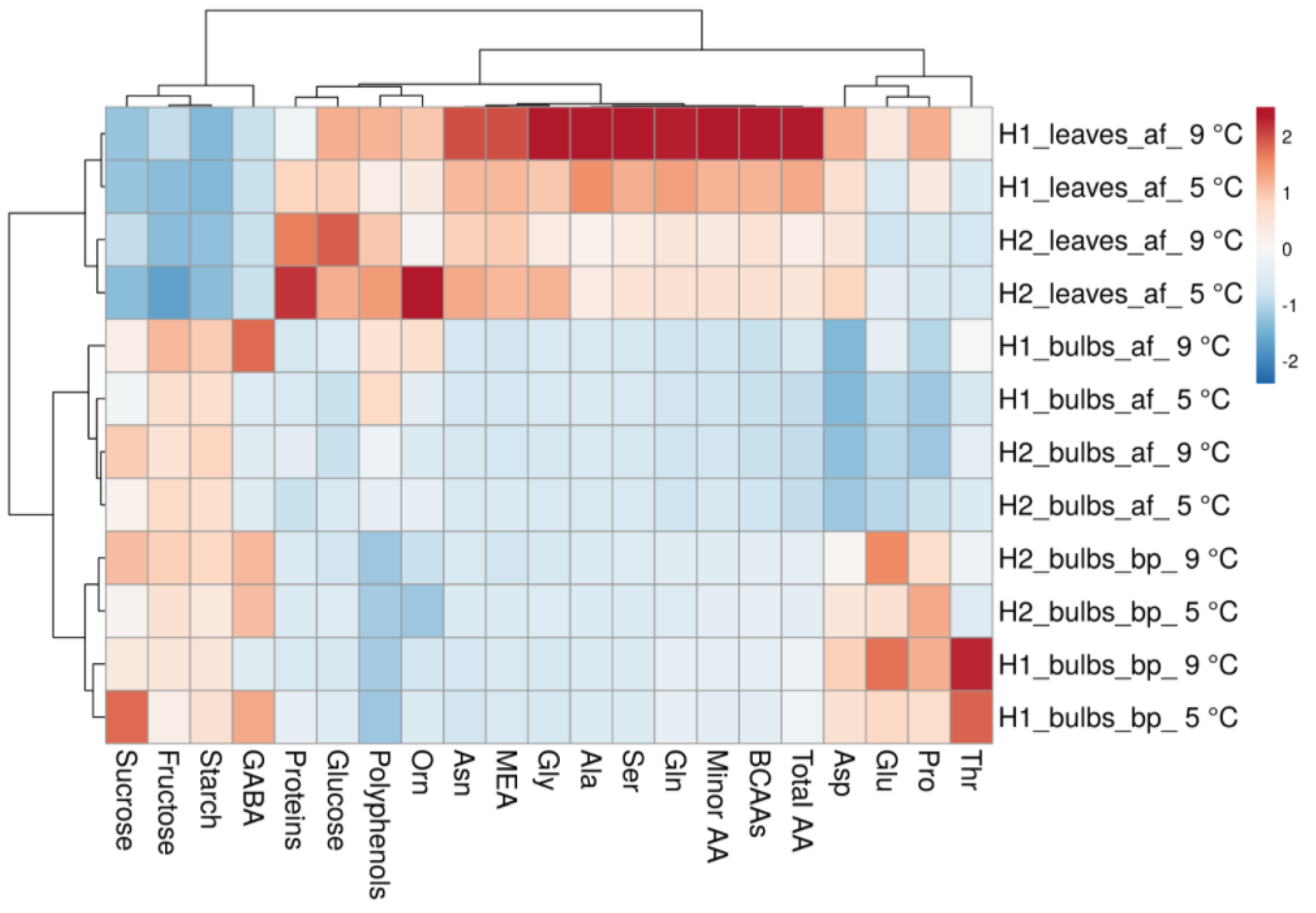
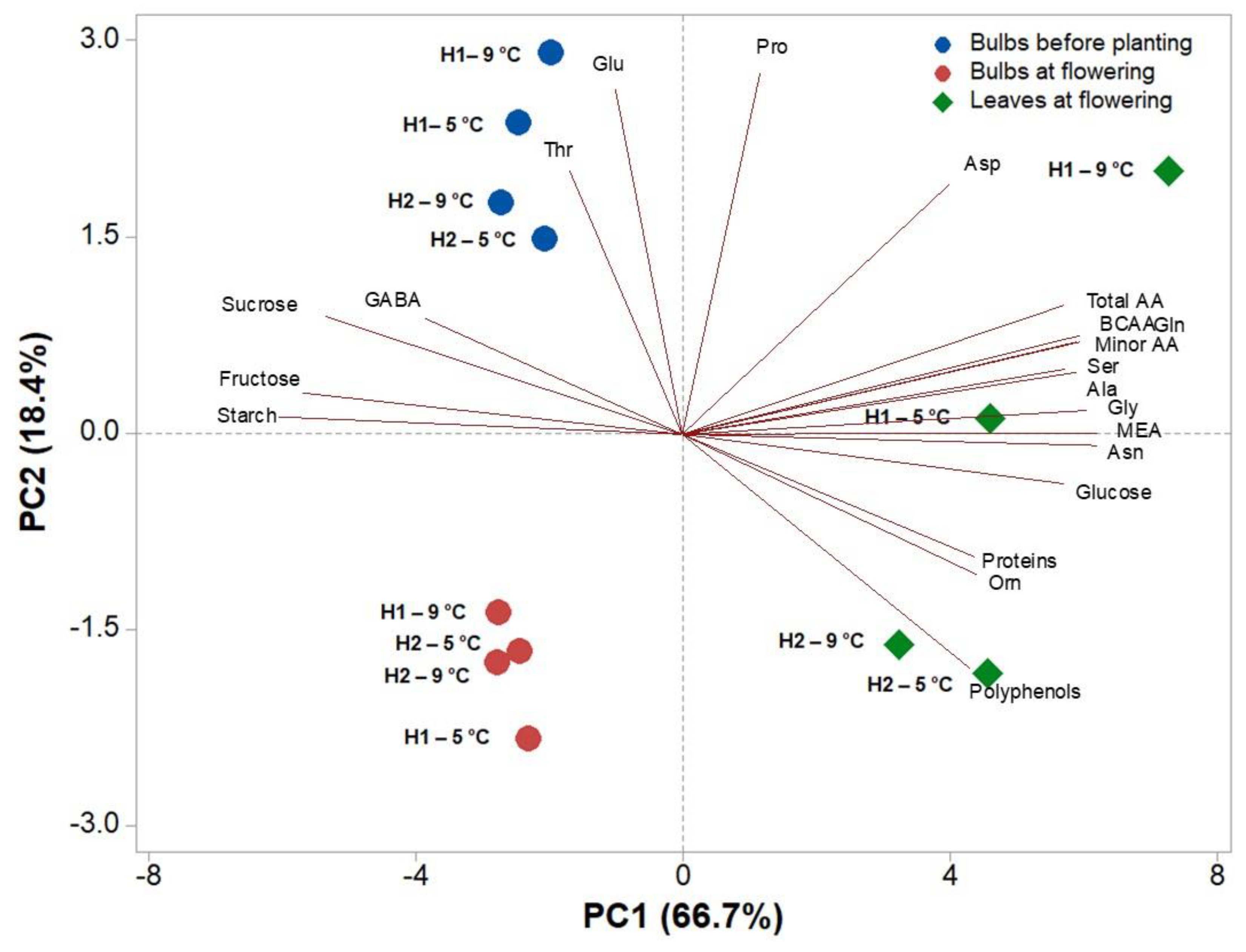
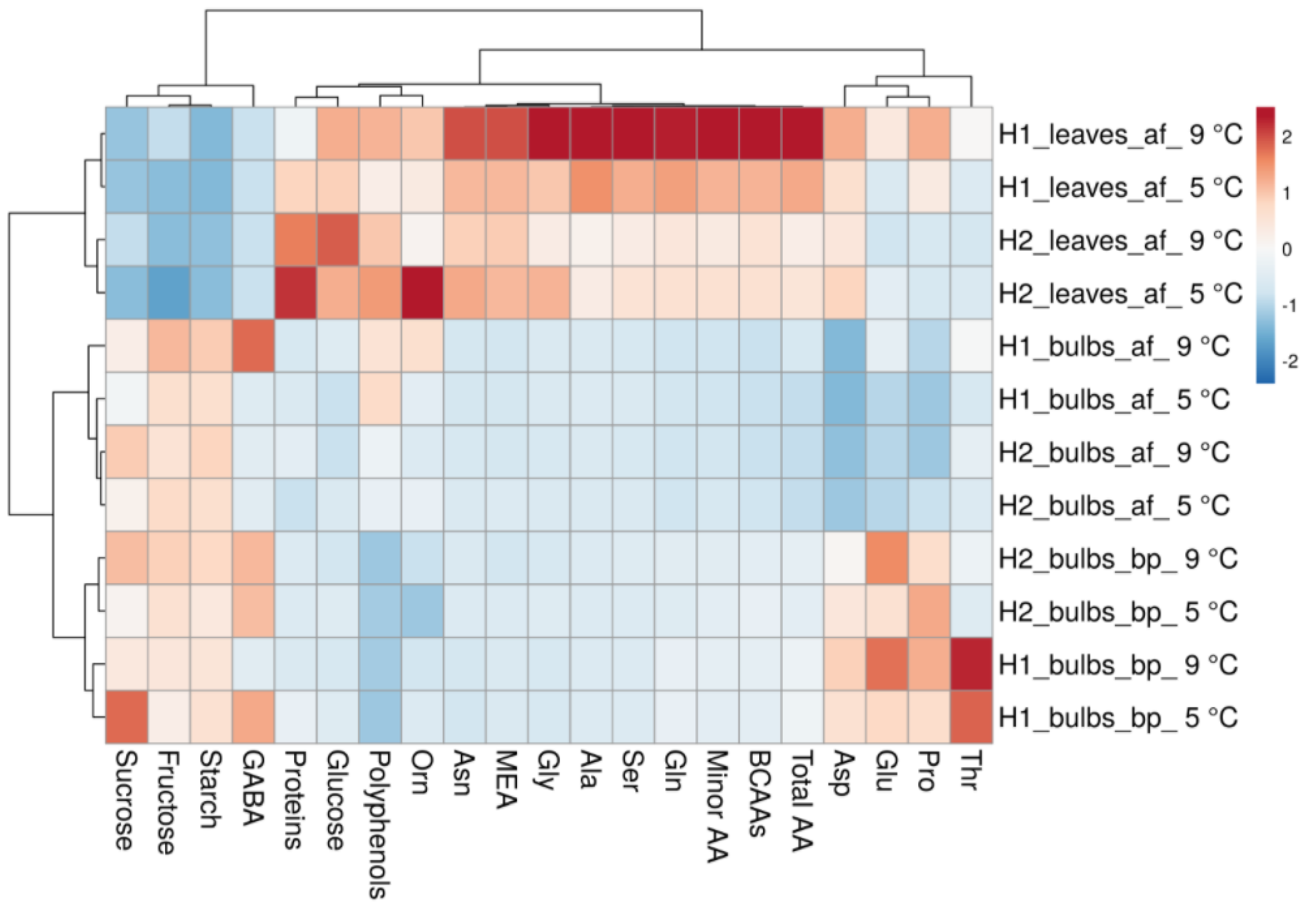
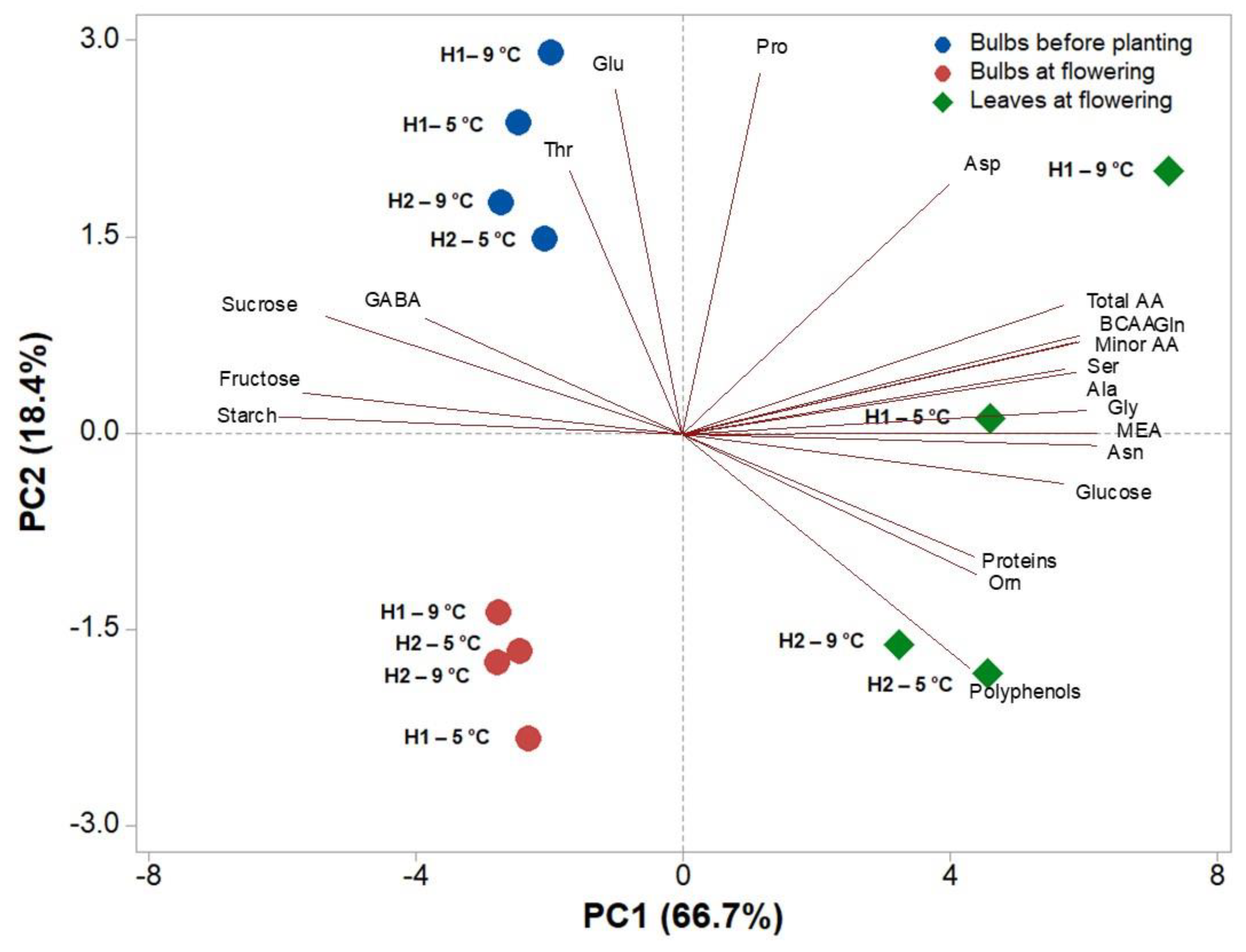
3.3. Heat Map and Principal and Component Analysis of the Analysed Biochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benschop, M.; Kamenetsky, R.; Le Nard, M.; Okubo, H.; De Hertogh, A. The global flower bulb industry: Production, utilization, research. Hortic. Rev. 2010, 36, 1–115. [Google Scholar]
- Le Nard, M.; De Hertogh, A. Tulipa. In The Physiology of Flower Bulbs; Elsevier Science: Amsterdam, The Netherlands, 1993; pp. 617–682. [Google Scholar]
- Mulder, R.; Luyten, I. De periodieke ontwikkeling van de Darwin tulp. Verh. K. Ned. Akad. Wet. Afd. Natuurkd 1928, 2, 1–64. [Google Scholar]
- Kamenetsky, R.; Zemah, H.; Ranwala, A.P.; Vergeldt, F.; Ranwala, N.K.; Miller, W.B.; Van As, H.; Bendel, P. Water status and carbohydrate pools in tulip bulbs during dormancy release. New Phytol. 2003, 158, 109–118. [Google Scholar] [CrossRef]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valluru, R.; Van den Ende, W. Plant fructans in stress environments: Emerging concepts and future prospects. J. Exp. Bot. 2008, 59, 2905–2916. [Google Scholar] [CrossRef] [Green Version]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, H.; Rook, F.; Kolloffel, C. Carbohydrate status of tulip bulbs during cold-induced flower stalk elongation and flowering. Plant Physiol. 1994, 104, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, H.; Wang, Y.; Yu, S.; Zheng, Y.; Chan, Z. Comparative physiological and metabolomic analyses reveal natural variations of tulip in response to storage temperatures. Planta 2019, 249, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Seng, S.; Li, D.; Zhang, F.; Liu, Y.; Yao, T.; Liang, J.; Yi, M.; Wu, J. Antagonism between abscisic acid and gibberellin regulates starch synthesis and corm development in Gladiolus hybridus. Hortic. Res. 2021, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Royal General Dutch Bulbgrowers Society. Classified List and International Register of Tulip Names; Hillegom, KAVB: Hillegom, The Netherlands, 1996; 188p. [Google Scholar]
- De Hertogh, A.; Le Nard, M. (Eds.) The Physiology of Flower Bulbs; Elsevier Science: Amsterdam, The Netherlands, 1993; 795p. [Google Scholar]
- Walinga, I.J.J.; van der Lee, V.J.G.; van Vark, W.; Novozamsky, I. Plant Analysis Manual; KAP: Wageningen, The Netherlands, 1995. [Google Scholar]
- Kjeldahl, J. A new method for the determination of nitrogen in organic matter. Z. Anal. Chem. 1883, 22, 366. [Google Scholar] [CrossRef] [Green Version]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity duration differently modulates physiological parameters and metabolites profile in roots of two contrasting barley genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; De Pascale, S.; Rouphael, Y. Sensory and functional quality characterization of protected designation of origin ’Piennolo del Vesuvio’ cherry tomato landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of combined applications of Trichoderma virens and a biopolymer-based biostimulant on lettuce agronomical, physiological, and qualitative properties under variable N regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Ciarmiello, L.F.; Piccirillo, P.; Carillo, P.; De Luca, A.; Woodrow, P. Determination of the genetic relatedness of fig (Ficus carica L.) accessions using RAPD fingerprint and their agro-morphological characterization. Afr. J. Bot. 2015, 97, 40–47. [Google Scholar] [CrossRef]
- Carillo, P.; Arena, C.; Modarelli, G.C.; De Pascale, S.; Paradiso, R. Photosynthesis in Ranunculus asiaticus L.: The influence of the hybrid and the preparation procedure of tuberous roots. Front. Plant Sci. 2019, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Dell’Aversana, E.; Modarelli, G.C.; Fusco, G.M.; De Pascale, S.; Paradiso, R. Metabolic profile and performance responses of Ranunculus asiaticus L. hybrids as affected by light quality of photoperiodic lighting. Front. Plant Sci. 2020, 11, 597823. [Google Scholar] [CrossRef]
- Carillo, P.; Modarelli, G.C.; Fusco, G.M.; dell’Aversana, E.; Arena, C.; De Pascale, S.; Paradiso, R. Light spectral composition affects metabolic response and flowering in non-vernalized Ranunculus asiaticus L. Environ. Exp. Bot. 2021, 192, 104649. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Krishnan, S.R.; Pandian, S.; Mareeswaran, N.; Aruni, W.; Pandian, S.K.; Ramesh, M. Global analysis of threonine metabolism genes unravel key players in rice to improve the abiotic stress tolerance. Sci. Rep. 2018, 8, 9270. [Google Scholar] [CrossRef] [PubMed]
- Strauss, G.; Hauser, H. Stabilization of lipid bilayer vesicles by sucrose during freezing. Proc. Natl. Acad. Sci. USA 1986, 83, 2422–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, W.; Secor, J.; Schrader, L.E. Rapid accumulation of γ-aminobutyric acid and alanine in soybean leaves in response to an abrupt transfer to lower temperature, darkness, or mechanical manipulation. Plant Physiol. 1984, 75, 170–175. [Google Scholar] [CrossRef]
- Rekarte-Cowie, I.; Ebshish, O.S.; Mohamed, K.S.; Pearce, R.S. Sucrose helps regulate cold acclimation of Arabidopsis thaliana. J. Exp. Bot. 2008, 59, 4205–4217. [Google Scholar] [CrossRef] [Green Version]
- Klein, W.; Weber, M.H.; Marahiel, M.A. Cold shock response of Bacillus subtilis: Isoleucine-dependent switch in the fatty acid branching pattern for membrane adaptation to low temperatures. J. Bacteriol. Res. 1999, 181, 5341–5349. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Xie, S.; Nie, L.; Zheng, Y.; Wang, J.; Huang, J.; Zhao, M.; Zhu, S.; Hou, J.; Chen, G.; et al. Comparative proteomics reveals cold acclimation machinery through enhanced carbohydrate and amino acid metabolism in Wucai (Brassica campestris L.). Plants 2019, 8, 474. [Google Scholar] [CrossRef] [Green Version]
- Castander-Olarieta, A.; Montalbán, I.A.; De Medeiros Oliveira, E.; Dell’Aversana, E.; D’Amelia, L.; Carillo, P.; Steiner, N.; Fraga, H.; Guerra, M.P.; Goicoa, T.; et al. Effect of thermal stress on tissue ultrastructure and metabolite profiles during initiation of radiata pine somatic embryogenesis. Front. Plant Sci. 2019, 9, 2004. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing sustainability by improving plant salt tolerance through macro- and micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Plant Leaf Area (cm2/Plant) | Flower Stem Height (cm) | Flower Stem Diameter (mm) | Flower Stem Total FW (g/Flower Stem) | Flower Stem Total DW (g/Flower Stem) | Flower Bud DW (g) | Stem DW (g) | Leaf DW (g) | Bulb DW (g) | |
|---|---|---|---|---|---|---|---|---|---|
| ‘Royal Virgin’ (H1) | |||||||||
| 5 °C | 122.9 ± 34.4 | 40.7 ± 0.3 | 8.0 ± 0.6 | 18.4 ± 2.0 | 1.56 ± 0.08 | 0.40 ± 0.06 | 1.16 ± 0.06 | 0.96 ± 0.23 | 7.05 ± 0.94 |
| 9 °C | 147.3 ± 22.3 | 42.2 ± 1.2 | 8.3 ± 0.9 | 20.9 ± 1.7 | 2.00 ± 0.15 | 0.40 ± 0.02 | 1.60 ± 0.13 | 1.11 ± 0.17 | 8.07 ± 1.15 |
| ‘Ad Rem’ (H2) | |||||||||
| 5 °C | 144.6 ± 40.6 | 31.5 ± 0.3 | 9.3 ± 0.9 | 17.9 ± 1.6 | 1.82 ± 0.27 | 0.43 ± 0.02 | 1.39 ± 0.27 | 0.91 ± 0.10 | 6.61 ± 0.96 |
| 9 °C | 119.3 ± 5.9 | 43.0 ± 1.5 | 9.3 ± 0.1 | 20.2 ± 0.8 | 1.93 ± 0.03 | 0.53 ± 0.03 | 1.40 ± 0.02 | 0.84 ± 0.08 | 10.25 ± 0.10 |
| Leaves | Bulb | |||||
|---|---|---|---|---|---|---|
| N tot % | p % | K % | N tot % | p % | K % | |
| ‘Royal Virgin’ (H1) | ||||||
| 5 °C | 3.17 ± 0.07 | 0.48 ± 0.08 | 18.03 ± 1.83 | 0.70 ± 0.07 | 0.07 ± 0.01 | 7.51 ± 0.50 |
| 9 °C | 3.62 ± 0.53 | 0.61 ± 0.06 | 7.64 ± 1.32 | 0.71 ± 0.07 | 0.07 ± 0.01 | 7.15 ± 0.41 |
| ‘Ad Rem’ (H2) | ||||||
| 5 °C | 2.79 ± 0.13 | 0.66 ± 0.01 | 17.36 ± 4.62 | 0.74 ± 0.19 | 0.11 ± 0.01 | 7.62 ± 0.52 |
| 9 °C | 2.83 ± 0.07 | 0.62 ± 0.03 | 19.17 ± 1.60 | 1.03 ± 0.38 | 0.09 ± 0.01 | 6.52 ± 0.24 |
| ‘Royal Virgin’ (H1) | ‘Ad Rem’ (H2) | Significance | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Source of Variance | 5 °C | 9 °C | Mean | 5 °C | 9 °C | Mean | Hybrid (H) | Preparation Procedure (P) | H × P |
| Bulbs before planting | |||||||||
| Soluble proteins | 46.7 | 43.2 | 45 | 43.7 | 43.9 | 43.8 | ns | ns | ns |
| Glucose | 37.94 a | 35.44 b | 36.7 | 37.85 a | 33.38 a | 35.6 | ns | * | ns |
| Fructose | 25.62 a | 27.99 b | 26.8 A | 29.66 ab | 33.11 c | 31.4 B | ** | ns | ** |
| Sucrose | 16.17 a | 9.18 b | 12.67 | 7.85 b | 12.59 b | 10.22 | ns | * | ns |
| Starch | 66.33 a | 63.49 a | 64.9 | 61.63 a | 71.95 b | 66.8 | ns | ns | * |
| Polyphenols | 0.92 a | 0.98 a | 0.96 | 0.98 a | 0.91 b | 0.95 | ns | ns | ns |
| Bulbs at flowering | |||||||||
| Soluble proteins | 43.47 | 42.43 | 42.95 | 39.94 | 45.79 | 42.86 | ns | ns | ns |
| Glucose | 31.01 a | 37.88 b | 34.45 | 36.30 ab | 31.79 a | 34.05 | ns | ns | * |
| Fructose | 30.66 | 36.07 | 33.37 | 31.48 | 29.12 | 30.3 | ns | ns | ns |
| Sucrose | 6.54 a | 8.63 b | 7.59 A | 8.08 a | 11.94 b | 10.01 B | * | ** | ns |
| Starch | 68.82 a | 76.71 b | 72.76 | 68.48 a | 74.19 b | 71.33 | ns | *** | * |
| Polyphenols | 4.46 | 4.12 | 4.29 A | 2.49 | 2.68 | 2.58 B | *** | ns | ns |
| Leaves at flowering | |||||||||
| Soluble proteins | 62.49 a | 48.87 ab | 55.68 A | 80.99 bc | 73.63 c | 77.31 B | * | ns | ** |
| Glucose | 66.49 a | 73.91 ab | 70.2 | 73.44 ab | 87.18 b | 80.31 | ns | ns | * |
| Fructose | 6.35 | 11.69 | 9.02 | 2.34 | 6.34 | 4.34 | ns | ns | ns |
| Sucrose | 0.99 a | 1.12 a | 1.06 | 0.65 a | 2.71 b | 1.68 | ns | * | *** |
| Starch | 11.07 | 10.03 | 10.55 A | 11.49 | 12.54 | 12.02 B | * | ns | ns |
| Polyphenols | 3.57 a | 5.26 b | 4.42 | 5.67 b | 4.09 b | 5.29 | ns | ns | ** |
| Chlorophyll a | 98.22 | 85.92 | 92.07 | 83.38 | 11.02 | 97.20 | ns | ns | ns |
| Chlorophyll b | 34.06 | 28.92 | 31.49 | 24.93 | 40.29 | 32.61 | ns | ns | ns |
| Carotenoids | 18.24 | 17.53 | 23.37 | 15.15 | 23.37 | 19.26 | ns | ns | ns |
| Source of Variance | Ala | Asn | Asp | GABA | Glu | Gln | Gly | MEA | Orn | Pro | Ser | Thr | Total AA | Minor AA | BCAAs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bulbs before planting | |||||||||||||||
| ‘Royal Virgin’ (H1) | |||||||||||||||
| 5 °C | 2.48 a | 0.57 a | 6.05 | 5.38 a | 10.9 a | 31.3 a | 0.12 a | 0.41 a | 0.3 a | 14.9 | 2.67 | 25.3 a | 134 a | 13.07 | 5.85 |
| 9 °C | 2.73 a | 0.7 a | 6.59 | 31.6 b | 15 b | 30.4 a | 0.12 a | 0.41 a | 0.27 a | 18.3 | 2.75 | 29.5 a | 126 a | 14.09 | 6.27 |
| Mean | 2.61 A | 0.64 A | 6.32 A | 15.5 A | 13 | 30.9 A | 0.12 | 0.41 | 0.29 A | 16.6 | 2.71 | 27.4 A | 130 A | 13.58 | 6.06 |
| ‘Ad Rem’ (H2) | |||||||||||||||
| 5 °C | 1.77 b | 3.18 b | 5.71 | 23.3 b | 10.1 a | 19.5 b | 0.56 b | 0.43 a | 0.2 b | 18.6 | 2.68 | 3.21 b | 103 b | 13.44 | 6.61 |
| 9 °C | 2.24 a | 2.84 b | 4.93 | 23.8 b | 14.3 b | 22.4 b | 0.1 a | 0.31 b | 0.25 ab | 14.9 | 3.16 | 5.66 b | 108 b | 12.52 | 5.87 |
| Mean | 2.01 B | 3.01 B | 5.32 B | 23.6 B | 12.2 | 20.9 B | 0.33 | 0.37 | 0.22 B | 16.7 | 2.92 | 4.43 B | 105 B | 12.98 | 6.24 |
| Hybrid (H) | ** | ** | * | * | ns | * | ns | ns | ** | ns | ns | ** | *** | ns | ns |
| Preparation procedure (P) | ns | ns | ns | ns | *** | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| H × P | * | * | ns | ** | ns | ** | * | * | * | ns | ns | *** | ** | ns | ns |
| Bulbs at flowering | |||||||||||||||
| ‘Royal Virgin’ (H1) | |||||||||||||||
| 5 °C | 0.99 a | 1.26 a | 2.07 | 5.05 | 3.22 b | 5.69 a | 0.19 a | 0.33 | 0.32 a | 2.97 a | 0.99 a | 1.64 a | 30.4 a | 5.35 a | 2.46 |
| 9 °C | 0.79 ab | 0.94 a | 2.04 | 5.38 | 5.91 a | 4.27 b | 0.19 a | 0.3 | 0.48 b | 4.23 ab | 0.91 b | 7.81 ab | 65.4 b | 5.42 a | 2.49 |
| Mean | 0.89 A | 1.1 | 2.05 | 5.21 | 4.57 | 4.98 A | 0.19 | 0.32 | 0.4 | 3.6 | 0.95 | 4.72 | 47.9 | 5.39 | 2.47 |
| ‘Ad Rem’ (H2) | |||||||||||||||
| 5 °C | 0.74 b | 2.59 b | 2.41 | 5.37 | 3.19 b | 2.47 c | 0.22 a | 0.43 | 0.33 a | 4.94 b | 1.18 a | 2.78 ab | 35.3 a | 8.35 b | 2.76 |
| 9 °C | 0.59 b | 1.46 a | 2.19 | 5.72 | 3.14 b | 2.45 c | 0.07 b | 0.35 | 0.3 a | 2.95 a | 0.73 b | 4.72 b | 30.8 a | 5.82 a | 2.16 |
| Mean | 0.66 B | 2.02 B | 2.3 | 5.55 | 3.16 | 2.46 B | 0.15 | 0.39 | 0.32 | 3.94 | 0.95 | 3.75 | 33.1 | 7.09 | 2.46 |
| Hybrid (H) | * | * | ns | ns | ns | *** | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Preparation procedure (P) | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | * | ** | ns | ns | ns |
| H × P | * | * | ns | ns | * | *** | *** | ns | *** | * | ns | *** | * | * | ns |
| Leaves at flowering | |||||||||||||||
| ‘Royal Virgin’ (H1) | |||||||||||||||
| 5 °C | 79 a | 32.3 | 6.16 | 1.09 | 4.91 a | 126 a | 4.99 ab | 1.57 | 0.44 a | 12.9 a | 38.7 a | 2.7 a | 359 a | 47.35 a | 18.4 a |
| 9 °C | 116 b | 47.3 | 7.3 | 1.13 | 9.29 b | 178 b | 9.13 a | 2.14 | 0.53 a | 18.3 b | 65 a | 7.9 b | 537 b | 74.73 b | 28.2 b |
| Mean | 97.5 A | 39.8 | 6.73 | 1.11 | 7.1 | 152 A | 7.06 A | 1.86 | 0.48 | 15.6 A | 51.8 A | 5.3 A | 448 A | 61.04 A | 23.3 A |
| ‘Ad Rem’ (H2) | |||||||||||||||
| 5 °C | 35.3 c | 34.7 | 6.55 | 1.11 | 5.64 a | 82.5 a | 5.61 ab | 1.57 | 0.76 b | 6.16 c | 24.6 b | 2.29 a | 242 c | 34.51 a | 14 a |
| 9 °C | 29.9 c | 28.5 | 5.72 | 0.85 | 4.02 a | 73.4 c | 2.94 b | 1.43 | 0.4 a | 6.42 c | 20.3 c | 1.3 a | 205 d | 29.5 a | 13.5 a |
| Mean | 32.6 B | 31.6 | 6.13 | 0.98 | 4.83 | 77.9 B | 4.28 B | 1.5 | 0.58 | 6.29 B | 22.5 B | 1.8 B | 224 B | 32.01 B | 13.8 B |
| Hybrid (H) | ** | ns | ns | ns | ns | ** | * | ns | ns | ** | ** | * | ** | ** | ** |
| Preparation procedure (P) | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| H × P | *** | ns | ns | ns | ** | * | * | ns | *** | *** | * | *** | * | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carillo, P.; Fusco, G.M.; Del Gaudio, R.S.; De Pascale, S.; Paradiso, R. Cold Treatment Modulates Changes in Primary Metabolites and Flowering of Cut Flower Tulip Hybrids. Horticulturae 2022, 8, 371. https://doi.org/10.3390/horticulturae8050371
Carillo P, Fusco GM, Del Gaudio RS, De Pascale S, Paradiso R. Cold Treatment Modulates Changes in Primary Metabolites and Flowering of Cut Flower Tulip Hybrids. Horticulturae. 2022; 8(5):371. https://doi.org/10.3390/horticulturae8050371
Chicago/Turabian StyleCarillo, Petronia, Giovanna Marta Fusco, Rosaria Serena Del Gaudio, Stefania De Pascale, and Roberta Paradiso. 2022. "Cold Treatment Modulates Changes in Primary Metabolites and Flowering of Cut Flower Tulip Hybrids" Horticulturae 8, no. 5: 371. https://doi.org/10.3390/horticulturae8050371
APA StyleCarillo, P., Fusco, G. M., Del Gaudio, R. S., De Pascale, S., & Paradiso, R. (2022). Cold Treatment Modulates Changes in Primary Metabolites and Flowering of Cut Flower Tulip Hybrids. Horticulturae, 8(5), 371. https://doi.org/10.3390/horticulturae8050371