
Characterization of Young Shoot Population, Yield, and Nitrogen Demands of Tea (Camellia sinensis L.) Harvested under Different Standards




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Tea Plantation
2.2. Harvest Standard
2.3. Fertilization
2.4. Sampling and Measurement of Parameters
2.5. Statistical Analysis
3. Results
3.1. Weight per Young Shoot
3.2. Young Shoot Density
3.3. Shoot Yield
3.4. Shoot N Concentrations and N Contents
3.5. 15N Absorption
3.6. Young Shoot Harvest Index (Shi) and N Harvest Index (NHI)
4. Discussion
4.1. Characteristics of Shoot Populations under Different Harvest Standards
4.2. Biomass Allocation and Young Shoot Yields under Different Harvest Standards
4.3. N Absorption and Allocation under Different Harvest Standards
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.E.; Lee, B.J.; Hwang, J.A.; Ko, K.S.; Chung, J.O.; Kim, E.H.; Lee, S.J.; Hong, Y.S. Metabolic dependence of green tea on plucking positions revisited: A metabolomic study. J. Agric. Food Chem. 2011, 59, 10579–10585. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-Y.; Fang, Z.-T.; Lin, J.-K.; Sun, Y.; Du, Z.-Z.; Guo, Y.-L.; Liu, J.-H.; Liang, Y.-R.; Ye, J.-H. Complementary iTRAQ proteomic and transcriptomic analyses of leaves in tea plant (Camellia sinensis L.) with different maturity and regulatory network of flavonoid biosynthesis. J. Proteome Res. 2019, 18, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Samanta, T.; Kotamreddy, J.N.R.; Ghosh, B.C.; Mitra, A. Changes in targeted metabolites, enzyme activities and transcripts at different developmental stages of tea leaves: A study for understanding the biochemical basis of tea shoot plucking. Acta Physiol. Plant. 2016, 39, 11. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Li, W.; Zhao, L.; Meng, F.; Wang, Y.; Tan, H.; Yang, H.; Wei, C.; Wan, X.; et al. Tissue-specific, development-dependent phenolic compounds accumulation profile and gene expression pattern in tea plant [Camellia sinensis]. PLoS ONE 2013, 8, e62315. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Asil, M. Determination of optimum harvestable length of shoots in tea (Camellia sinensis L.) based on the current shoot growth, rather than interval plucking. J. Food Agric. Env. 2007, 5, 122–124. [Google Scholar]
- Cloughley, J.B. Effects of harvesting policy and nitrogen application rates on the production of tea in Central Africa. II. Quality and total value of the crop. Exp. Agric. 1983, 19, 47–54. [Google Scholar] [CrossRef]
- Owuor, P.O.; Ng’etich, W.K.; Obanda, M. Quality response of clonal black tea to nitrogen fertilizer, plucking interval and plucking standard. J. Sci. Food Agric. 2000, 80, 439–446. [Google Scholar] [CrossRef]
- Mouli, M.R.; Onsando, J.M.; Corley, R.H. Intensity of harvesting in tea. Exp. Agric. 2007, 43, 41–50. [Google Scholar] [CrossRef]
- Xu, W.; Song, Q.; Li, D.; Wan, X. Discrimination of the production season of Chinese green tea by chemical analysis in combination with supervised pattern recognition. J. Agric. Food Chem. 2012, 60, 7064–7070. [Google Scholar] [CrossRef]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Xie, D.; Tan, J.; et al. Nontargeted analysis using ultraperformance liquid chromatography–quadrupole time-of-flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality of tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef]
- Liu, J.W.; Zhang, Q.F.; Liu, M.Y.; Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Metabolomic analyses reveal distinct change of metabolites and quality of green tea during the short duration of a single spring season. J. Agric. Food Chem. 2016, 64, 3302–3309. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Yang, J.; Xie, X.; Lin, C. Over-supply of tea in China—A marketing analysis. Acta Tea Sinica 2017, 58, 75–79. [Google Scholar]
- Ruan, J.Y.; Gerendas, J. Absorption of foliar-applied urea-N-15 and the impact of low nitrogen, potassium, magnesium and sulfur nutritional status in tea (Camellia sinensis L.) plants. Soil Sci. Plant Nutr. 2015, 61, 653–663. [Google Scholar] [CrossRef]
- Chen, P.-A.; Lin, S.-Y.; Liu, C.-F.; Su, Y.-S.; Cheng, H.-Y.; Shiau, J.-H.; Chen, I.-Z. Correlation between nitrogen application to tea flushes and quality of green and black teas. Sci. Hortic. 2015, 181, 102–107. [Google Scholar] [CrossRef]
- Dutta, R.; Stein, A.; Smaling, E.M.A.; Bhagat, R.M.; Hazarika, M. Effects of plant age and environmental and management factors on tea yield in Northeast India. Agron. J. 2010, 102, 1290–1301. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Gerendas, J.; Hardter, R.; Sattelmacher, B. Effect of root zone pH and form and concentration of nitrogen on accumulation of quality-related components in green tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Gerendas, J.; Hardter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef]
- Stephens, W.; Carr, M.K. Responses of tea (Camellia sinensis) to irrigation and fertilizer. III. Shoot extension and development. Exp. Agric. 1993, 29, 323–339. [Google Scholar] [CrossRef]
- Stephens, W.; Carr, M.K. Responses of tea (Camellia sinensis) to irrigation and fertilizer. IV. Shoot population density, size and mass. Exp. Agric. 1994, 30, 189–205. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Haerdter, R.; Gerendas, J. Impact of nitrogen supply on carbon/nitrogen allocation: A case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biol. 2010, 12, 724–734. [Google Scholar] [CrossRef]
- Kamau, D.M.; Spiertz, J.H.J.; Oenema, O.; Owuor, P.O. Productivity and nitrogen use of tea plantations in relation to age and genotype. Field Crops Res. 2008, 108, 60–70. [Google Scholar] [CrossRef]
- Stephens, W.; Carr, M.K. Responses of tea (Camellia sinensis) to irrigation and fertilizer. I. Yield. Exp. Agric. 1991, 27, 177–191. [Google Scholar] [CrossRef]
- Möller, M.; Weatherhead, E.K. Evaluating drip irrigation in commercial tea production in Tanzania. Irrig. Drain. Syst. 2007, 21, 17–34. [Google Scholar] [CrossRef]
- Venkatesan, S.; Murugesan, S.; Ganapathy, M.N.K.; Verma, D.P. Long-term impact of nitrogen and potassium fertilizers on yield, soil nutrients and biochemical parameters of tea. J. Sci. Food Agric. 2004, 84, 1939–1944. [Google Scholar] [CrossRef]
- Cloughley, J.B.; Grice, W.J.; Ellis, R.T. Effects of harvesting policy and nitrogen application rates on the production of tea in Central Africa. I. Yield and crop distribution. Exp. Agric. 1983, 19, 33–46. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Ma, L.; Jiang, S.; Deng, M.; Lv, L.; Xu, Z.; Ruan, J. Thermo condition determines the uptake of autumn and winter applied nitrogen and subsequent utilization in spring tea (Camellia sinensis L.). Horticulturae 2021, 7, 544. [Google Scholar] [CrossRef]
- Lemaire, G.; Oosterom, E.v.; Sheehy, J.; Jeuffroy, M.H.; Massignam, A.; Rossato, L. Is crop N demand more closely related to dry matter accumulation or leaf area expansion during vegetative growth? Field Crops Res. 2007, 100, 91–106. [Google Scholar] [CrossRef]
- Lemaire, G.; Jeuffroy, M.H.; Gastal, F. Diagnosis tool for plant and crop N status in vegetative stage theory and practices for crop N management. Eur. J. Agron. 2008, 28, 614–624. [Google Scholar] [CrossRef]
- Balasuriya, J. Shoot population density and shoot weight of clonal tea (Camellia sinensis) at different altitudes in Sri Lanka. Eur. J. Agron. 1999, 11, 123–130. [Google Scholar] [CrossRef]
- De Costa, W.A.J.M.; Navaratne, D.M.S.; Anandacoomaraswamy, A. Physiological basis of yield variation of tea (Camellia sinensis) during different years of the pruning cycle in the central highlands of Sri Lanka. Exp. Agric. 2009, 45, 429–450. [Google Scholar] [CrossRef]
- Madamombe, G.; Tesfamariam, E.; Taylor, N. Yield decline in mechanically harvested clonal tea (Camellia sinensis (L.) O. Kuntze) as influenced by changes in source/sink and radiation interception dynamics in the canopy. Sci. Hortic. 2015, 194, 286–294. [Google Scholar] [CrossRef]
- Kalita, R.M.; Das, A.K.; Sileshi, G.W.; Nath, A.J. Ecosystem carbon stocks in different aged tea agroforestry systems: Implications for regional ecosystem management. Trop. Ecol. 2020, 61, 203–214. [Google Scholar] [CrossRef]
- Kamau, D.M.; Spiertz, J.H.J.; Oenema, O. Carbon and nutrient stocks of tea plantations differing in age, genotype and plant population density. Plant Soil 2008, 307, 29–39. [Google Scholar] [CrossRef]
- Li, S.; Wu, X.; Xue, H.; Gu, B.; Cheng, H.; Zeng, J.; Peng, C.; Ge, Y.; Chang, J. Quantifying carbon storage for tea plantations in China. Agric. Ecosyst. Envrion. 2011, 141, 390–398. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, Y.; Fan, D.; Zhu, Q.; Pan, Z.; Fan, K.; Wang, X. Temporal evolution of carbon storage in Chinese tea plantations from 1950 to 2010. Pedosphere 2017, 27, 121–128. [Google Scholar] [CrossRef]
- Ahmed, A.; de Costa, W.A.J.M.; Wijeratne, M.A. Effect of different plucking systems on yield and root starch reserve in two cultivars of tea (Camellia sinensis L.). J. Environ. Sci. Nat. Resour. 2016, 9, 91–95. [Google Scholar] [CrossRef][Green Version]
- Botwright, T.L.; Menary, R.C.; Brown, P.H. Photosynthesis and assimilate partitioning during rhythmic growth of green tea (Camellia sinensis var sinensis). J. Hortic. Sci. Biotechnol. 1998, 73, 806–811. [Google Scholar] [CrossRef]
- Magambo, M.J.; Cannell, M.G. Dry matter production and partition in relation to yield of tea. Exp. Agric. 1981, 17, 33–38. [Google Scholar] [CrossRef]
- Fageria, N.K. Nitrogen harvest index and its association with crop yields. J. Plant Nutr. 2014, 37, 795–810. [Google Scholar] [CrossRef]
- Fan, K.; Zhang, Q.; Tang, D.; Shi, Y.; Ma, L.; Liu, M.; Ruan, J. Dynamics of nitrogen translocation from mature leaves to new shoots and related gene expression during spring shoots development in tea plants (Camellia sinensis L.). J. Plant Nutr. Soil Sci. 2020, 183, 180–191. [Google Scholar] [CrossRef]
- Zhu, F.; Dai, L.; Hobbie, E.A.; Qu, Y.; Huang, D.; Gurmesa, G.A.; Zhou, X.; Wang, A.; Li, Y.; Fang, Y. Quantifying nitrogen uptake and translocation for mature trees: An in situ whole tree paired 15N labeling method. Tree Physiol. 2021, 41, 2109–2125. [Google Scholar] [CrossRef] [PubMed]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.F.; Shi, Y.Z.; Ruan, J.Y. Nitrogen absorption by field-grown tea plants (Camellia sinensis) in winter dormancy and utilization in spring shoots. Plant Soil 2019, 442, 127–140. [Google Scholar] [CrossRef]
- Dong, F.; Hu, J.; Shi, Y.; Liu, M.; Zhang, Q.; Ruan, J. Effects of nitrogen supply on flavonol glycoside biosynthesis and accumulation in tea leaves (Camellia sinensis). Plant Physiol. Biochem. 2019, 138, 48–57. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
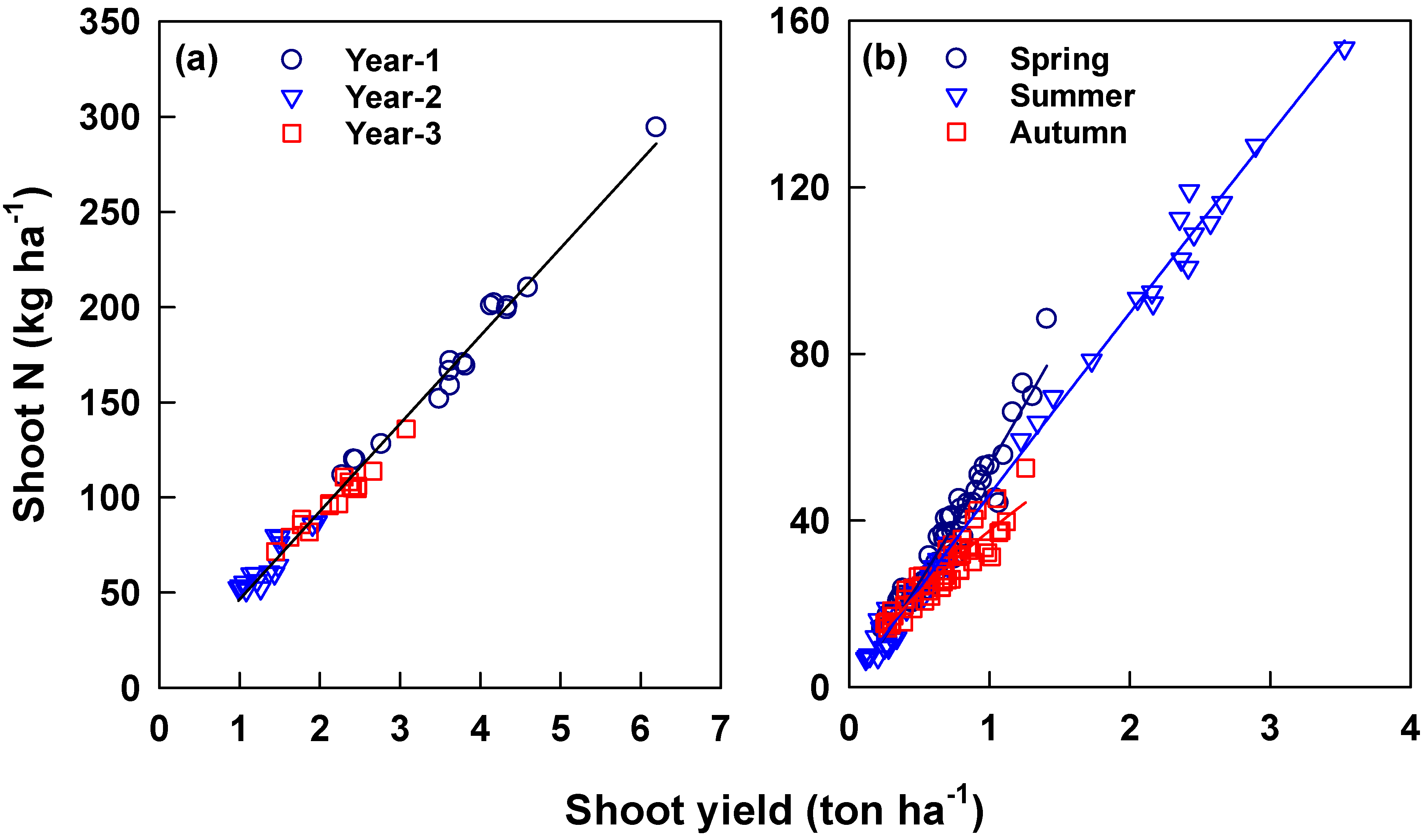
| Season | a | b | R2 | SE of Estimate |
|---|---|---|---|---|
| Early spring | 0.194 | 1.600 | 0.942 *** | 0.339 |
| Late spring | 0.840 | 1.926 | 0.914 *** | 0.505 |
| Spring | 0.626 | 1.776 | 0.966 *** | 0.285 |
| Summer | −0.226 | 3.330 | 0.825 *** | 1.306 |
| Autumn | −0.380 | 2.506 | 0.950 *** | 0.489 |
| Season | a | b | R2 | SE of Estimate |
|---|---|---|---|---|
| Spring tea | 53.216 | 1.085 | 0.925 **** | 4.364 |
| Summer tea | 46.157 | 0.962 | 0.993 **** | 3.396 |
| Autumn tea | 37.536 | 0.725 | 0.845 **** | 3.467 |
| Whole year | 46.435 | 0.997 | 0.986 **** | 6.300 |
| Organ | Harvest Standard | |||
|---|---|---|---|---|
| B + 1L | B + 2L | B + 3L | B + 1L/3L | |
| Plant biomass (ton ha−1) | 60.2 ± 3.7 | 62.5 ± 3.5 | 63.6 ± 8.1 | 65.2 ± 4.1 |
| Fibrous root biomass (ton ha−1) | 1.6 ± 0.4 | 2.5 ± 0.4 | 2.1 ± 0.7 | 2.9 ± 0.3 |
| Brown root biomass (ton ha−1) | 23.1 ± 2.2 b | 19.9 ± 1.0 b | 15.2 ± 1.7 a | 22.5 ± 2.9 b |
| Root/above-ground ratio | 0.71 ± 0.06 b | 0.56 ± 0.02 b | 0.38 ± 0.03 a | 0.64 ± 0.07 b |
| Shoot harvest index (SHI) (%) | 0.046 ± 0.004 | 0.058 ± 0.002 | 0.061 ± 0.006 | 0.058 ± 0.002 |
| Total 15N absorption (kg ha−1) | 42.4 ± 5.4 | 43.0 ± 5.1 | 43.0 ± 2.9 | 49.5 ± 4.1 |
| Shoot 15N (kg ha−1) | 12.0 ± 0.3 | 15.5 ± 1.4 | 16.0 ± 2.4 | 14.3 ± 0.9 |
| Share of shoot 15N in total 15N absorption (%) | 29.7 ± 3.4 | 36.8 ± 2.9 | 37.1 ± 4.5 | 29.0 ± 1.1 |
| 15N in final plant (kg ha−1) | 27.8 ± 5.3 | 25.1 ± 3.6 | 25.0 ± 2.8 | 33.1 ± 3.4 |
| Share of final plant 15N in total 15N absorption (%) | 63.8 ± 4.9 | 57.9 ± 1.8 | 58.3 ± 4.9 | 66.5 ± 1.8 |
| N harvest index (NHI) | 0.19 ± 0.02 | 0.23 ± 0.02 | 0.22 ± 0.02 | 0.18 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, L.; Shi, Y.; Ma, L.; Ruan, J. Characterization of Young Shoot Population, Yield, and Nitrogen Demands of Tea (Camellia sinensis L.) Harvested under Different Standards. Horticulturae 2022, 8, 275. https://doi.org/10.3390/horticulturae8040275
Long L, Shi Y, Ma L, Ruan J. Characterization of Young Shoot Population, Yield, and Nitrogen Demands of Tea (Camellia sinensis L.) Harvested under Different Standards. Horticulturae. 2022; 8(4):275. https://doi.org/10.3390/horticulturae8040275
Chicago/Turabian StyleLong, Lizhi, Yuanzhi Shi, Lifeng Ma, and Jianyun Ruan. 2022. "Characterization of Young Shoot Population, Yield, and Nitrogen Demands of Tea (Camellia sinensis L.) Harvested under Different Standards" Horticulturae 8, no. 4: 275. https://doi.org/10.3390/horticulturae8040275
APA StyleLong, L., Shi, Y., Ma, L., & Ruan, J. (2022). Characterization of Young Shoot Population, Yield, and Nitrogen Demands of Tea (Camellia sinensis L.) Harvested under Different Standards. Horticulturae, 8(4), 275. https://doi.org/10.3390/horticulturae8040275