1. Introduction
Over the last few decades, the recurrent flowering, or everbearing (EB), strawberry cultivars have steadily expanded their share of commercial production due to their high yield potential and long cropping season. While below-average fruit taste quality has limited their use in the past, new cultivars with acceptable fruit quality are now available [
1,
2]. In contrast to the traditional seasonal flowering cultivars, the EBs are quantitative long day (LD) plants at intermediate temperatures (18–21 °C) and obligatory LD plants at high (27 °C) temperatures. Only at low temperatures (≤15 °C) are they day-neutral [
3,
4,
5,
6]. Because of their recurrent-flowering character, EBs are very difficult to propagate in a vegetative state for experimental use, even when using runner tips. Accordingly, most flowering studies with EB cultivars have been started with flowering-induced plants. However, the problem can be circumvented by using seed propagated F1 hybrids [
7], which produce genetically identical plants which are not influenced by previous flowering history [
3,
4,
5].
In Europe, the EB cultivars are mainly used for annual production in plastic tunnels and greenhouses. Runner tips are cut and rooted in late July and raised as tray plants under natural outdoor conditions until early November. Under the relatively low temperature conditions prevailing during the raising period, the plants will initiate flower buds also in short day (SD) cf. [
8], and by the end of the season, they are ready to flower under appropriate forcing conditions. After overwintering in cold stores at −1.5 °C, the plants are commonly marketed as Tray or Mini Tray plants according to the size of the plants and root volume of the trays, or as bare-rooted Frigoplants [
9]. Early rooting will generally produce larger plants with more flower trusses and accordingly, with a larger yield potential. Plant quality grading is usually done by measurement of crown diameter. For early production, the plants are established on so-called table-top gutter systems in greenhouses or plastic tunnels in March–April and cropped under natural light conditions throughout the entire season until late October [
1,
2]. For summer production, the plants are planted in the field in April–May, depending on the local climatic conditions.
However, a serious limitation with this production system is that it does not give a continuous and steady supply of ripe berries but rather produces a series of flowering and fruiting flushes separated by gaps with little or no flowers and ripe fruits. Thus, the first fruit flush, which originates from inflorescences initiated during plant raising in the previous year [
10,
11], is always followed by an off period of 2–3 or more weeks with no flowering and fruiting [
1,
2,
10,
11]. Usually, this is followed by an oscillation in flowering and fruiting, resulting in repeated on and off periods during the summer season. This represents a big challenge for commercial production since it causes non-continuous berry supply and reduced total berry yields.
Due to the SD conditions prevailing during much of the raising period, the plants will also enter a semi-dormant state that is typical for all strawberry types during late fall and winter [
6,
8]. Therefore, several weeks of chilling are needed for dormancy release and re-establishment of normal elongation of flower trusses during cropping [
2].
Obviously, there are several factors that can influence the recurrent flowering tendency and the seasonal patterns of fruiting flushes in EB strawberries. Melis (2020) reported that the larger first flush of the big Tray plants delayed initiation of the recurrent flower flush compared with the smaller Mini Tray plants, which produced a smaller first flush. Therefore, he argued that the presence of a heavy crop load tends to suppress the initiation of a recurrent flower flush. This concurs with the results by Sønsteby et al. (2021) [
11], who found that floral initiation in ‘Favori’ plants was source limited in plants with a high fruit/leaf ratio. However, when comparing the results from a range of experiments, there is no overall positive relationship between fruit load and the length of the non-flowering off period. Accordingly, there seem to be other modifying factors involved, for example, the duration of winter chilling, which was shown to be excessive in the present production practice (1).
In order to obtain a better understanding of these complex issues, we have performed a plant raising experiment with the commonly used EB cultivars ‘Favori’ and ‘Murano’ in which rooting date and temperature and photoperiod during plant raising were varied factorially. Since previous experience with SF (SD) cultivars has shown that it is difficult to produce plants with satisfactory yield potential in the cool Nordic environment [
12], strawberry producers in the Nordic countries usually buy their plants from The Netherlands. The aim of the experiment was to develop optimal protocols for raising plants under Nordic climate conditions and compare the yield potential of plants produced in The Netherlands and in Nordic climates.
2. Materials and Methods
2.1. Plant Material and Cultivation
Rooted runners from certified plants of the cultivars ‘Favori’ and ‘Murano’ were planted in 3-L plastic pots filled with a mixture of 80% limed and fertilized sphagnum peat and 20% granulated perlite (v/v) in early May 2020 and grown as stock plants in a greenhouse maintained at 25 °C and 10 h photoperiod (08.00 h–16.00 h). On 15 July, 1 August, and 15 August, respectively, young runner plants of each cultivar were cut from stock plants and rooted directly in multi-cell trays (Bato Strawberry Trays 9-holes, Bato Plastics B. V., The Netherlands) filled with a peat-based potting soil (Gartnerjord, LOG, Oslo). All runners were rooted in a water-saturated atmosphere in a greenhouse kept at 20 °C and 10-h photoperiod. After 15 days, when all plants had formed roots, the plants were divided into two groups and further raised at the NIBIO Experimental Center Apelsvoll in southeast Norway (60°40′ N, 10°40′ E, 250 m.a.s.l.) in the following conditions:
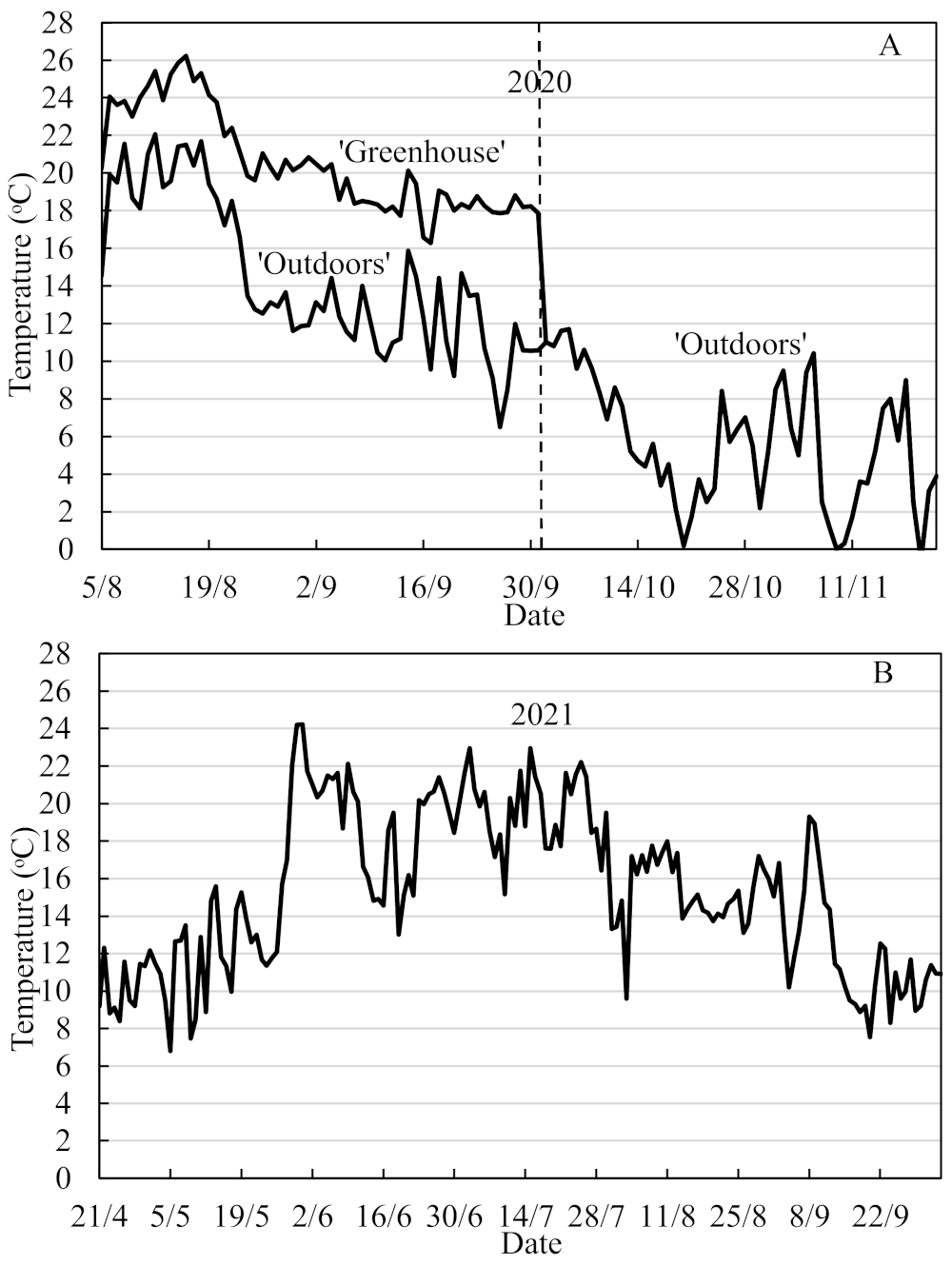
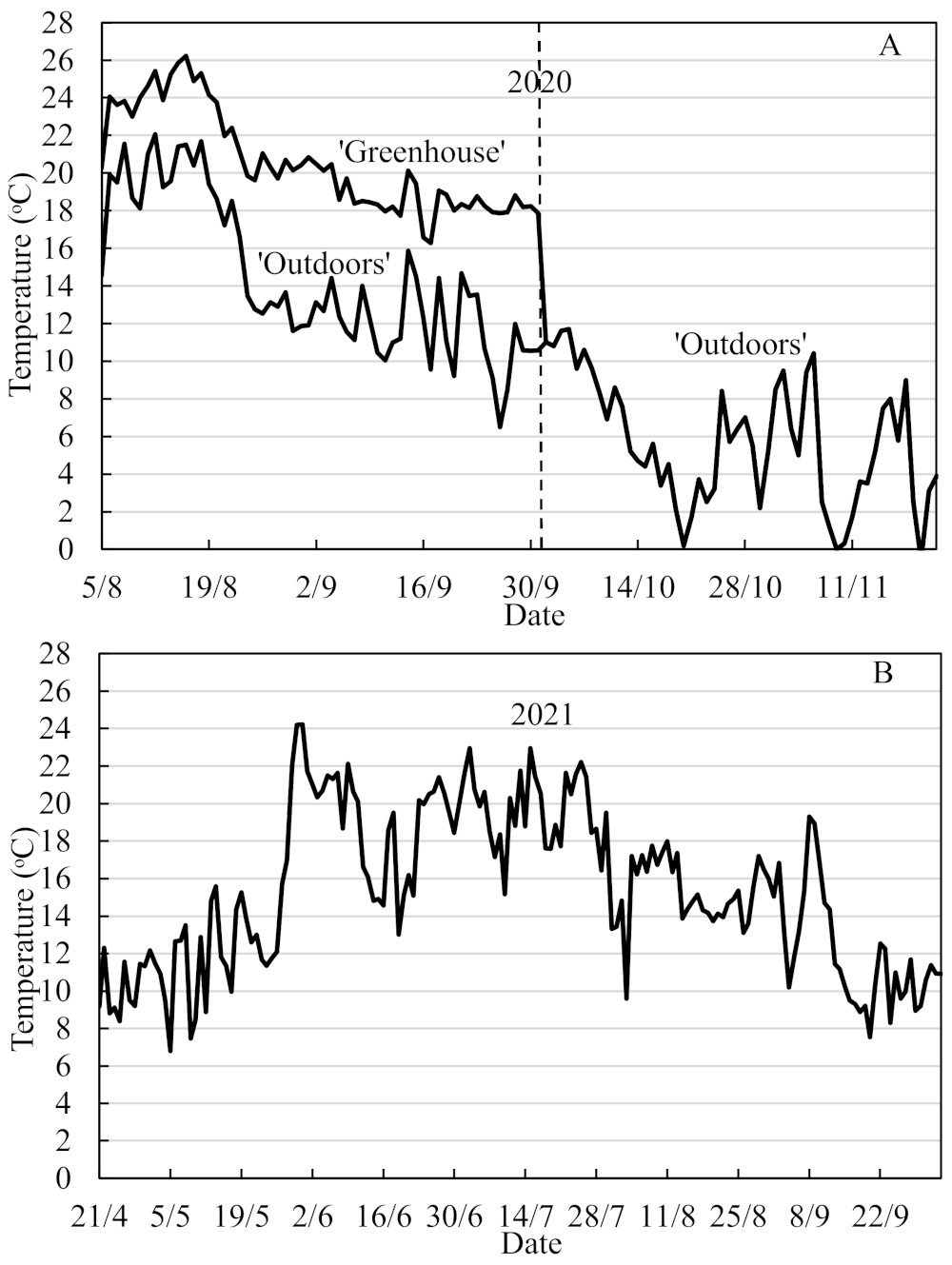
Daylength extension to 20 h was provided by low-intensity light (15 µmol quanta m
−2 s
−1) from 70 W incandescent lamps, and the daily mean temperatures at Apelsvoll during the 5 August–22 November period are shown in
Figure 1A. On 1 October, all plants were moved outdoors where they remained until 22 November when they were partly defoliated and moved into a cold store and overwintered at −1.5 °C in boxes lined with a polyethylene film. ‘Favori’ plants produced in The Netherlands and imported as frozen plants in spring 2021 were also used for comparison. On 14 April 2021, all plants were transplanted into 50 cm long table-top gutter-like containers with 8 L capacity (3 plants in each) in a mixture of 80% limed and fertilized peat and 20% granulated perlite. After a 9 day establishment period in an unheated plastic greenhouse under a double layer of fiber cloth, the plants were then, on 18 May, placed in an open Haygrove plastic tunnel, where they remained for the entire cropping season. From then onwards, the plants were drip-irrigated with a nutrient solution containing a 1:1 mixture of Calcinit™ and Kristalon Indigo™ (Yara, Norway) with electric conductivity of 1.6 mS cm
−1. As a protection against mildew, all plants were sprayed with an elemental sulfur suspension (Thiovit Jet
®, Syngenta, Basel, Switzerland) at planting and weekly after planting until the end of May, and with 3 chemical pesticides until July. Beneficials (
Amblyseius cucumeris) against thrips and spider mites were applied 2–3 times throughout the growing season. The daily mean temperatures in the tunnel during the cropping season are shown in
Figure 1B.
2.2. Experimental Design and Data Collection and Analysis
The experiment had a split-plot design with raising environments as main plots and rooting dates and cultivars as sub-plots. Each treatment had 3 replicates, each consisting of 9 plants grown in 3 adjacent containers and representing 1.5 m running row. Crown diameters and number of crowns were recorded in each plant, and photos of representative plants (top and root clump) were taken in spring before planting (
Figure 2). New runners were removed and counted weekly as soon as they appeared throughout the cropping season. Ripe berries were harvested 2–3 times per week from 16 June to 2 October. The number and weight of all berries, including rotten berries, were recorded as well as the proportion of berries with a diameter >28 mm. At the termination of harvest on 2 October, crop height, number of crowns per plant, and plant fresh weight (excluding runners and roots) were recorded for all plants, as well as the number of flowers and fruits not reaching maturity.
Experimental data were subjected to analysis of variance (ANOVA) using the MiniTab® Statistical Software program package (Release 15; Minitab Inc., State College, PA, USA). Percentage values were always subjected to an arc sin transformation before the performance of the ANOVA, and separation of significant treatment means was performed by Fisher’s least significant difference (LSD) method.
3. Results
The results in
Table 1 show that crown diameter was larger in ‘Favori’ than in ‘Murano’ plants and generally declined with delayed rooting date in both cultivars. However, due to highly significant two-factor interactions, the main effect of the raising environment was not statistically significant. Unexpectedly, plants of both cultivars that were rooted 15 July and subsequently raised in the greenhouse environment had a smaller crown diameter than the corresponding plants raised outdoors, and were smaller than their counterparts rooted on 1 August. The photos in
Figure 2 show that these smaller plants raised in the greenhouse had a smaller and less developed root system as well. The number of crowns per plant was higher in ‘Murano’ and decreased as rooting was delayed. The dimensions of ’Favori’ plants produced in The Netherlands closely matched those of ‘Favori’ plants rooted on 1 August and raised outdoors in Norway.
The results in
Table 2 show that also the total berry yield was significantly higher in ‘Favori’ than in ‘Murano’ plants, while the main effects of rooting dates and raising environment were nonsignificant due to highly significant two-and three-factor interactions of the treatment variables. This was particularly marked in ‘Favori’, in which delayed rooting markedly decreased yield in plants raised outdoors, whereas it had the opposite effect in plants raised in the greenhouse. To a lesser extent, this was also the case in ’Murano’. The same trend was also found for berry size and the percentage of berries with diameter > 28 mm, whereas the number of berries per plant varied only between the cultivars, being highest in ‘Favori’. While earliness of ripening (days to first pick) was less affected by cultivars and treatments (
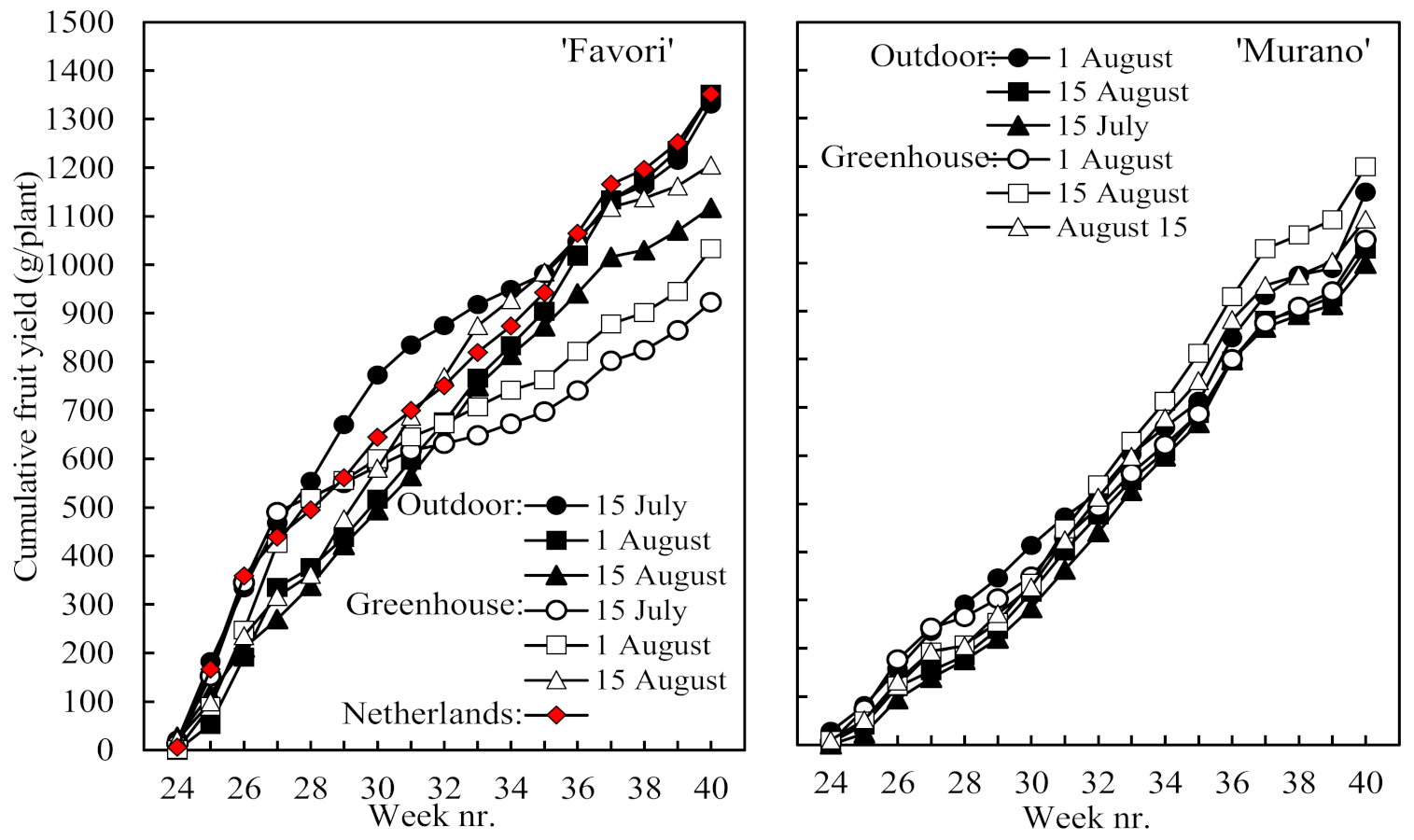
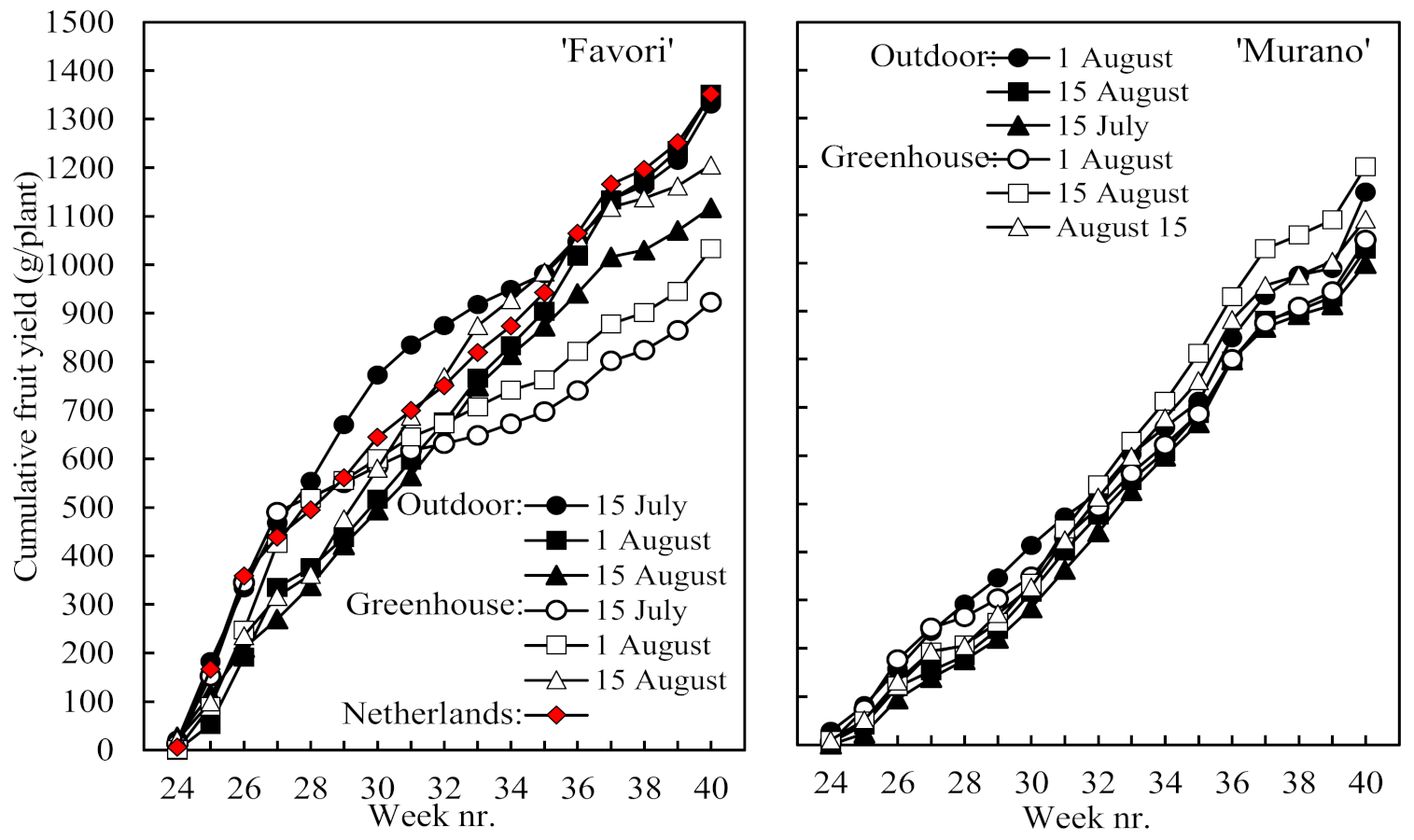
Table 2), the time courses of cumulative yield in
Figure 3 demonstrate enhanced early yield in ‘Favori’ compared with ‘Murano’, especially in plants rooted on 15 July and raised outdoors. It is also seen from
Table 2 that plants rooted on 1 August and raised outdoors in Norway closely matched those produced in The Netherlands on all the yield and earliness criteria.
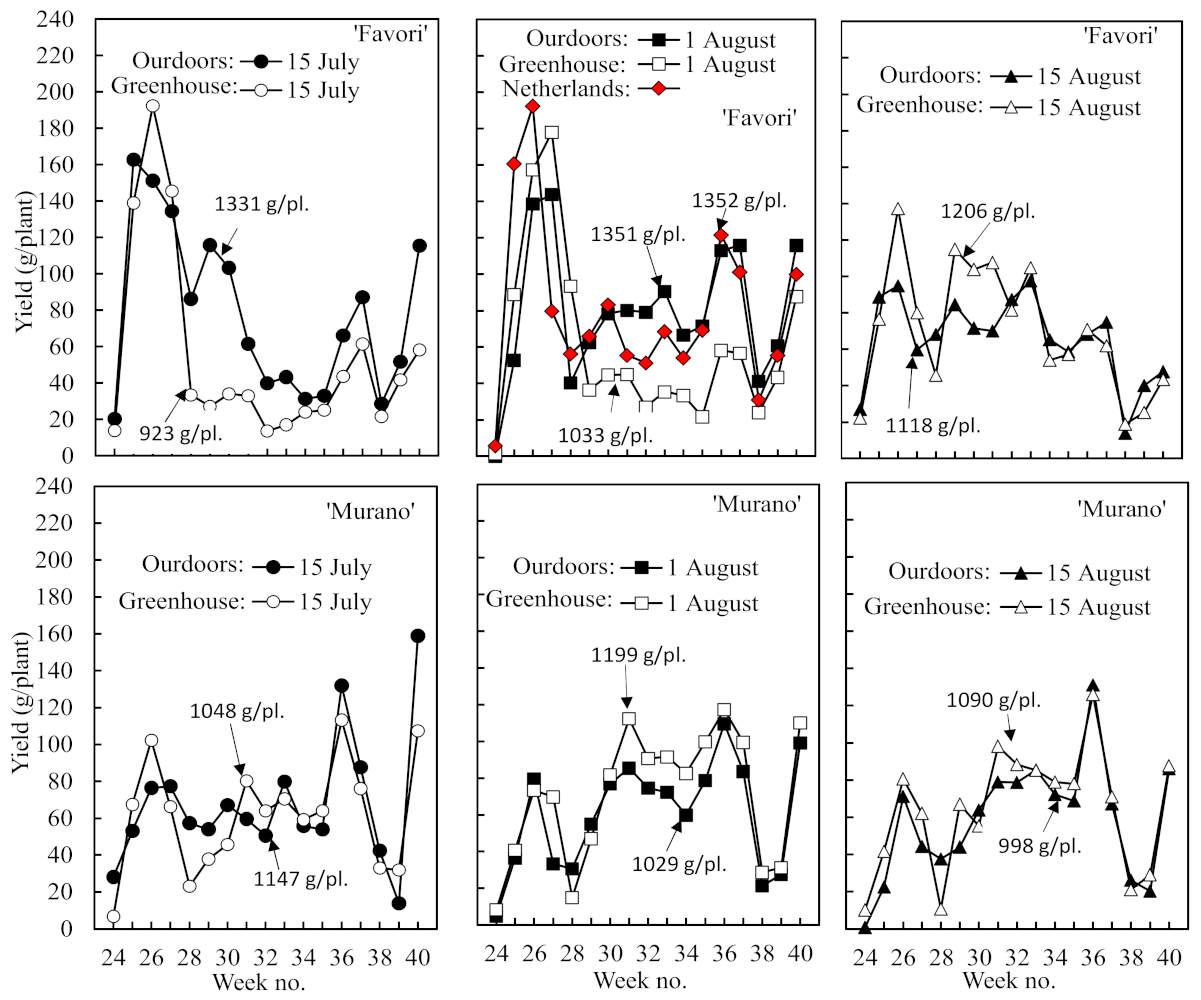
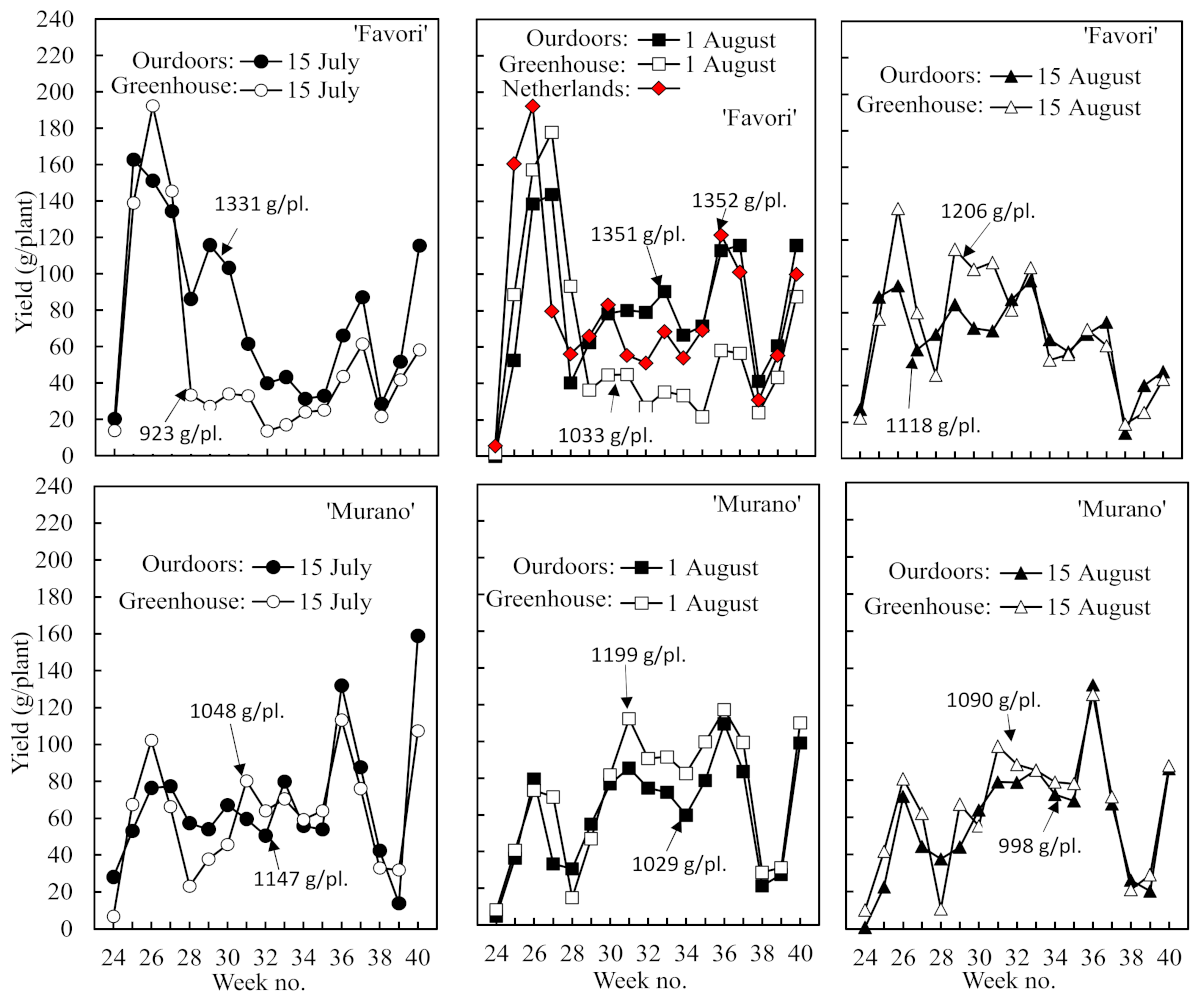
The temporal distribution of the harvest and the length of the off periods between fruit flushes also varied markedly between cultivars and treatments (
Figure 4). While the greater share of the fruit yield was harvested in the early part of the season in ‘Favori’, it was the opposite in ‘Murano’. There was also a clear trend to an extended off period in association with the larger first flush in ‘Favori’ and a more even distribution of the harvest across the season in ‘Murano’ plants with a smaller first flush. Again, the performance of the ‘Favori’ plants of Dutch origin was identical to that of the Norwegian plants rooted on 1 August and raised outdoors.
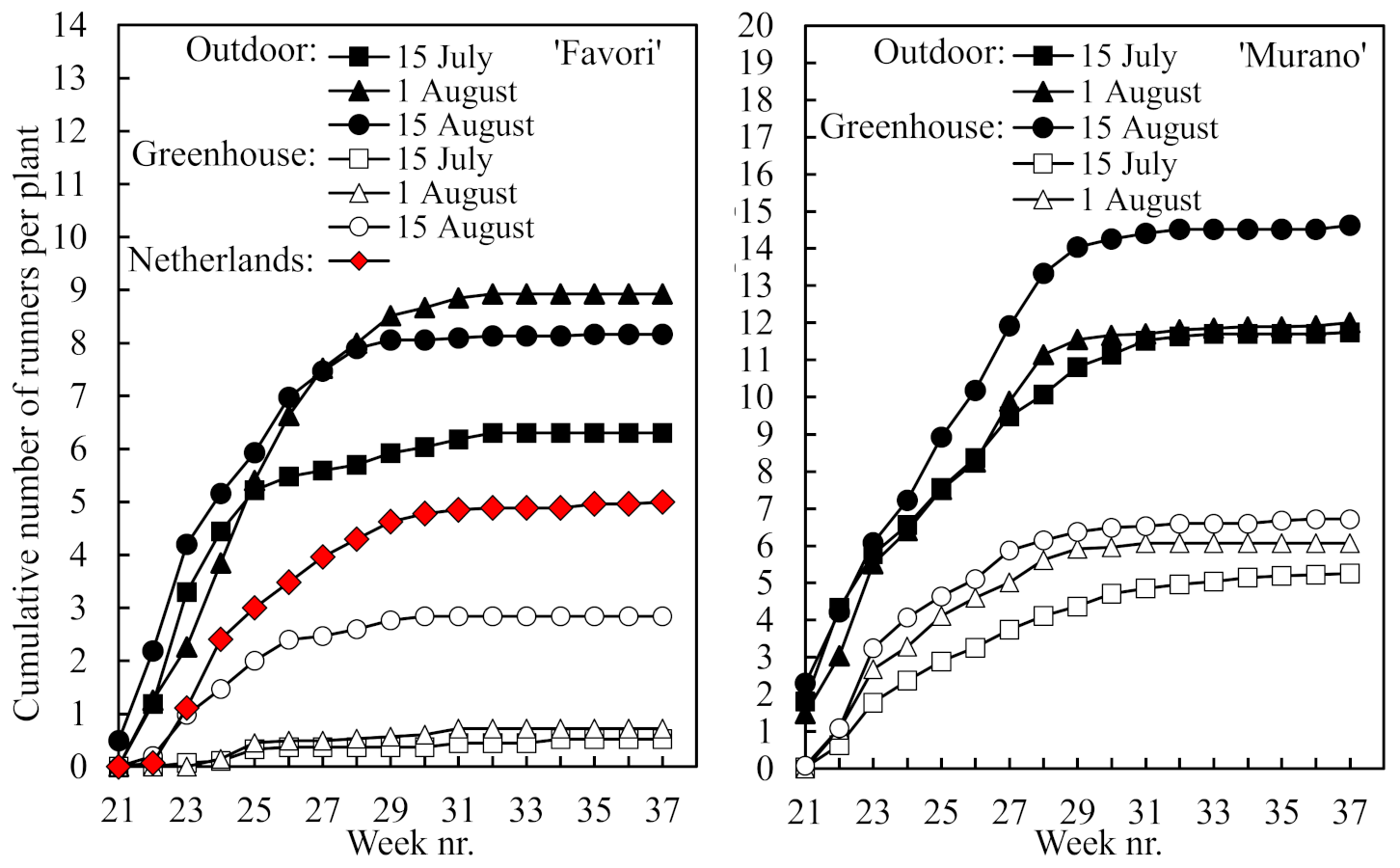
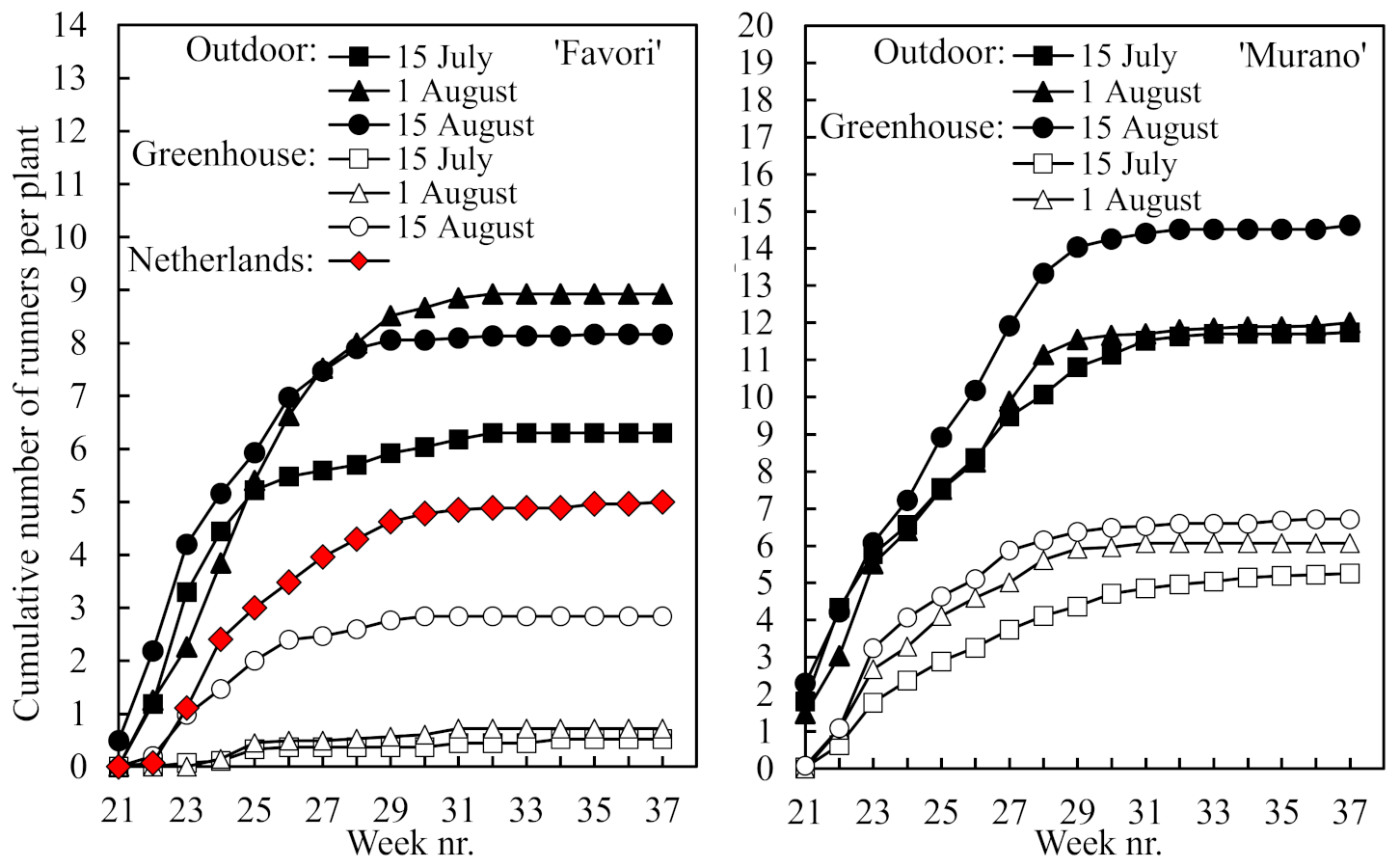
The total number of runners produced per plant during the cropping season is presented in
Figure 5. While the number was almost twice as high in ‘Murano’ than in ‘Favori’, it was significantly higher in the plants raised outdoors than in those raised in the greenhouse, and the number declined the earlier the plants were rooted (for statistics, see
Table 3). However, the number of runners was lower in the Dutch ‘Favori’ plants than in the Norwegian plants rooted on 1 August and raised outdoors. All runners were formed during the first half of the cropping season.
Some aspects of plant structure at termination of the harvest are shown in
Table 3. The number of crowns per plant was rather similar across cultivars and treatments, whereas above-ground fresh weight was significantly higher in plants raised outdoors. There was also significant interaction of environment and rooting date, the weights being higher in early rooted plants raised outdoors but not in those raised in the greenhouse. There was also a pronounced interaction of cultivar x raising environment, the ‘Favori’ plants being markedly larger when raised outdoors, while there was no such effect in ‘Murano’. In both cultivars, the total number of inflorescences produced during the season was higher in plants raised outdoors. While the number was not significantly affected by rooting date, it was generally highest in plants rooted on 1 August. The number of flowers and fruits that did not reach maturity before the harvesting was terminated on 2 October was nearly twice as high in ‘Murano’ than in ‘Favori’, while the effect of rooting date and environment was rather small and variable.
4. Discussion
The results of the experiment revealed significant effects of rooting date and the raising environment on plant yield potential and harvest distribution in both cultivars. The highest yield of 1.350 g/plant was obtained in ‘Favori’ plants rooted on 1 August and raised outdoors. While in both cultivars, the yield of plants raised outdoors declined with delayed rooting, the inverse trend was found in ‘Favori’ plants raised in the greenhouse, while no consistent effect of rooting date was observed in ‘Murano’ plants raised in the greenhouse (
Table 2). The reason for this complex three-factor interaction seems to be severalfold. First, the unexpected low yield of ‘Favori’ plants rooted on 15 July and raised in the greenhouse was in part due to crowdedness caused by excessive leaf and plant growth in LD at 18 °C. Also, under high temperature and LD conditions, strawberry plants allocate less leaf assimilates to the roots and more to the above-ground parts [
13], thus producing a plant with a high top/root ratio under these conditions (cf.
Figure 2). Such plants were probably more prone to waterlogging during the establishment period when the plants were generously watered. Furthermore, the long high temperature/LD period that the early-rooted plants were exposed to also resulted in some mold infestation and leaf yellowing in the dense leaf canopy of these plants. Contrary to expectation, however, exposure to LD conditions did not advance floral initiation in these plants. These results indicate that the ‘Murano’ plants were less influenced by the raising environment than were ‘Favori’ plants. Generally, high yield was closely associated with increased berry size and less to the number of berries, but more so in ‘Favori’ than in ‘Murano’.
Earliness of ripening and berry harvest was generally superior in ‘Favori’, especially in plants raised outdoors. Clearly, a larger share of the total crop was harvested in the early part of the harvest season in ‘Favori’ than in ‘Murano’ (
Figure 3 and
Figure 4). Despite the expected advantage of LD conditions in the greenhouse, ripening lagged behind in the ‘Favori’ plants from the greenhouse, and more the earlier the plants had been rooted (
Figure 3). Daylength neutrality of the plants at low temperature has obviously contributed to these effects.
The seasonal pattern of fruit flushes and off periods also varied significantly between the cultivars and treatments. The large first fruit flush in ‘Favori’ was associated with an extended off period, while the smaller first flush in ‘Murano’ was associated with a more even harvest distribution during the rest of the season (
Figure 4). This concurs with the results of Melis (2020) [
2], who argued that this is due to the repression of renewed floral initiation by the heavy fruit load. This hypothesis is further supported by the fact that the degree of repression was reduced as the size of the first flush was diminished by delayed rooting (
Figure 4). Furthermore, Sønsteby et al. (2021) [
11] found that floral initiation was source-limited in ‘Favori’ plants with a high fruit/leaf ratio. However, as already mentioned, we have observed that the seasonal distribution of the crop can also be modified by temperature in the cropping year.
That flowering indirectly suppresses runner formation is well documented in strawberries [
1,
6]. Therefore, and because runners are known as strong sinks for leaf assimilates in strawberry [
11,
14], fruit yield is usually negatively related to runner numbers [
1,
11,
14]. However, this effect was not clearly seen in the present experiment. Although the number of runners was higher in ‘Murano’ than in the higher-yielding ‘Favori’, the latter cultivar had more runners in the high-yielding plants that were raised outdoors (
Figure 5). The reason for this is not clear but may be related to the suboptimal root growth status of the ‘Favori’ plants raised in the greenhouse (
Figure 2). It should be kept in mind that the runners are initiated concurrently with flower primordia in the year before flowering and fruiting [
11], and it is thus possible that both runner and flower initiation has been hampered by the reduced vigor of the ‘Favori’ plants raised in the greenhouse environment.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}